
Naglaa F. El-Orabi
Department of Pharmacology, College of Pharmacy, King Saud University, Riyadh, Kingdom of Saudi Arabia
Omar H. Abd-Elkader
Department of Zoology, College of Science, King Saud University, Riyadh, Kingdom of Saudi Arabia
Dean D. Schwartz
Department of Anatomy, Physiology and Pharmacology, College of Veterinary Medicine, Auburn University, Auburn, AL, USA
International Journal of Pharmacology
Year: 2013 | Volume: 9 | Issue: 7 | Page No.: 430-441







ABSTRACT
Exertional heat injury represents a major risk for people working or exercising in hot environments. Currently, no pharmacological therapies are available and little is known about the molecular response to heat stress in the brain. This study examined changes in gene expression associated with hyperthermia in the hippocampus of pigs. Twelve pigs were kept at either 70°F (control) or 90°F (heat stressed) for 4, 8, 12 and 24 h. The heart rate, respiratory rate and body temperature of the pigs were monitored hourly. At 4, 8, 12 and 24 h, the hippocampus was excised from three pigs in each group and total RNA was obtained for use in differential display PCR. A total of 31 differentially expressed cDNAs were isolated. Differential expressions of eight genes were further confirmed by real-time PCR analysis. Only slight increases in the expression HSP70, HSP90 and HSP27 genes were reported after heat stress at all time points. After 12 h of heat stress, DNA polymerase ε p12 gene expression was stimulated to 2.55-fold of control level while it was down-regulated after both 4 and 8 h of heat stress exposure. Two genes encoding proteasome 26S subunits, PSMD10 and PSMB9, were also induced after exposure to heat stress for 4, 8 and 12 h to 2-fold or more of control levels. After 24 h, only PSMD10 was still elevated significantly. The superoxid dismutase-1 gene was significantly inhibited after both 8 and 12 h of heat stress, 0.39 and 0.45 fold of control levels respectively. The sodium-channel voltage gated β1 subunit gene was inhibited after heat stress. This inhibition was only significant after 24 h. In conclusion, most of the hippocampal genes of which the expression is altered by heat stress are play a role in cellular protection versus hyperthermic insult and some of the altered genes may play a role in heat-related brain pathogenesis. A greater understanding of heat-induced molecular changes in the brain could facilitate development of effective therapeutical strategies for treatment of heat-related illness.
PDF Abstract XML References Citation
Received: December 11, 2013;
Accepted: February 19, 2014;
Published: April 15, 2014
How to cite this article
Naglaa F. El-Orabi, Omar H. Abd-Elkader and Dean D. Schwartz, 2013. Differential Expression of Hippocampal Genes under Heat Stress. International Journal of Pharmacology, 9: 430-441.
DOI: 10.3923/ijp.2013.430.441
URL: https://scialert.net/abstract/?doi=ijp.2013.430.441
DOI: 10.3923/ijp.2013.430.441
URL: https://scialert.net/abstract/?doi=ijp.2013.430.441
INTRODUCTION
Heat stress usually denotes perceived discomfort and physiological strain associated with exposure to a hot environment, due to physical exercise or daily work, for a long period (Bergeron et al., 2011; Bouchama and Konchel, 2002). Exertional heat illness is a critical risk to individuals working or exercising in the heat and significant morbidity and mortality have resulted from heat stroke associated with military training, as well as to athletes exercising during warm weather, or individuals participating in mining activities (Mountjoy et al., 2012; Khosla and Guntupalli, 1999). Heat stress-related mortality is one of the main influences of climatic extremes on human society and it accounts for at least 7% of wilderness-related deaths (Kysely, 2004; Lugo-amador et al., 2004). Some individuals are more vulnerable to heat stress complications even if they live in temperate environments, such as patients suffering from chronic medical conditions, the elderly, pediatric and disabled populations (Hermesh et al., 2000; Coris et al., 2004) and people engaged in activities involving exposure to excessive heat such as fire fighting and intense exercising (Basu, 2009; Chou et al., 2003). Heat stress and associated hyperthermia can be presented as a spectrum of clinical symptoms ranging from weakness, headache, profuse sweating and malaise to severe life threatening complications, such as hyperpyrexia, delirium, multiple organ failure and death (Atha, 2013; Chia and Teo, 2003). No pharmacological preventative or intervention is available for heat-provoked sickness. This is due mainly to the lack of understanding of the molecular and cellular responses to heat stress even though the medical symptoms are well known (Sharma and Hoopes, 2003).
Heat stress has a potent effect in all biological systems. Studies at the cellular level in several animal models and in humans suggest that heat can directly induce nervous tissue injury and brain dysfunction which have long-term behavioral, physiological and neuropathological consequences (Sharma and Hoopes, 2003). Brain tissues appear to be more susceptible to heat than other body tissues (Maroni et al., 2003).
The mechanisms of brain injury after hyperthermic insults are not completely understood (Le Greves et al., 1997). No single factor is responsible for all the neuropathological changes seen in the central nervous system (CNS). In fact, the final outcome is due to a combination of many factors and neurochemicals exerting a synergistic influence on the CNS (Sharma and Hoopes, 2003).
At the molecular level, exposure to stressors elicits activation of specific signaling pathways, leading to a stimulus-specific program of gene expression that is related to cell differentiation, proliferation, survival, or programmed death (apoptosis). Mammalian cells respond to stresses, such as increased environmental temperature and oxidative and chemical stressors with activation of heat-shock proteins (HSPs) (Fulda et al., 2010; Sherman and Goldberg, 2001). HSPs are molecular chaperones that assist many cellular processes involving proteins, including folding, transport through membranes, degradation, escape from aggregation and reactivation of denatured proteins (Borges and Ramos, 2005). Additionally, HSPs facilitate cellular protection through anti-apoptotic actions (Franklin et al., 2005). HSPs play a central role in allowing cells to adapt to gradual changes in their environment and to survive in otherwise lethal conditions (Garrido et al., 2001). Thermal stress also leads to changes in expression of a substantial number of genes (ca.50) other than HSPs which affect a number of diverse mechanisms, such as membrane transport, metabolism and growth (Gupta et al., 2013; Sonna et al., 2002).
In the brain, little is known about changes in gene expression in systems other than HSPs in response to elevated environmental temperature (Sharma and Hoopes, 2003). In the present study, the hippocampus was chosen as a representative brain region. Hippocampal formation is involved in integration of information arriving from different sensory organs and their associated brain regions and is essential for memory storage and retrieval, playing an important role in learning, motivation and emotion (Fountoulakis et al., 2005). Moreover, it is highly sensitive and vulnerable to the effect of trauma and different stressors, including heat stress (Maroni et al., 2003; McEwen and Gianaros, 2010). Previous studies have shown that whole body hyperthermia in rats cause profound edema and cell injury in the hippocampus and lead to upregulation of both constitutive and inducible isoforms of nitric oxide synthase (cNOS and iNOS), heme oxygenase-2 (Alm et al., 2000; Sharma et al., 2000), HSP70 (Maroni et al., 2003), HSP27 and HSP32 (Bechtold and Brown, 2003). It also enhances phosphorylation and activation of different kinases, such as members of the MAPK family and PKB and stimulates the activation of transcription factors, such as HSF and MEF2 (Maroni et al., 2003).
In the current study, we investigated the differential gene expression profile in the pig hippocampus in response to elevated temperature in order to identify novel genes associated with the molecular response of the brain to heat stress. These genes could be considered as pharmacological targets for the development of effective therapeutic interventions to treat heat-related brain tissue injuries and associated illnesses.
MATERIALS AND METHODS
Animals and heat stress: Twenty four pigs (100 lbs each) were acclimated for 48 h and then moved to pens and maintained at either 70°F (control groups) or 90°F (heat-stressed groups) for 4, 8, 12, or 24 h. The heart rate, respiration, behavior and, body temperature of the pigs were monitored hourly. At the end of each time point, 3 pigs in each group were euthanized using pentobarbital sodium (50 mg kg-1 body weight) and the hippocampi were excised and rapidly stored in RNALater (Ambion Inc., Austin, TX, USA) and stored at -20°C until required for use. The experiment was repeated twice.
RNA isolation: Total RNA was extracted from homogenized hippocampi using the RNeasy midi kit (Qiagen, Valencia, CA, USA). RNA quality and quantity were assessed by using denaturing-formaldehyde /agarose gel electrophoresis and UV spectrophotometry (ratio of 260-to-280 nm). All RNA samples were stored at -80°C until required for gene expression analysis.
Analysis of gene expression: The mRNA differential display PCR (DD-PCR) was performed using RNAimage kits (Genhunter, Nashville, TN, USA), as described previously (Liang et al., 1994). In brief, total RNA from 12 h samples was reverse transcribed using three different one-base anchored primer (H-T11M, where M = G, C, or A). Then, the resulting cDNAs were amplified by polymerase chain reaction (PCR), using a combination of one of three 3' oligo dT-anchored primers (H-T11M) and one of 80 arbitrary 5' primers (HAPx, x = 1-80); a total of 240 primer pairs were used. Finally, the amplified cDNAs were separated on a 6% denaturizing polyacrylamide gel, blotted onto a sheet of 3M paper and dried at 80°C. The blot was exposed to BioMax Kodak film for 24-48 h at -80°C. The film was developed and the bands differentially expressed between the control and heat-stressed groups were excised from the gel according to the pattern on the film. The PCR products were extracted from the dried gel and the DNA was precipitated. The DNA pellet was dissolved in 10 μL nuclease-free water. Isolated DNA was reamplified by PCR using the original primer pair.
Selected reamplified DNA fragments were ligated directly into the pCRII plasmid vector (TA Cloning® kit, Invitrogen, Carlsbad, CA, USA) and the ligation products transformed into One Shot INVαF` E. coli cells (Invitrogen). Plasmids were isolated using the Wizard® Plus Minipreps DNA Purification System (Promega Corp., Madison, WI, USA) and EcoRI restriction digestion was performed to verify the presence of the insert.
Plasmid solutions (100-200 ng μL-1) were prepared in nuclease-free water and sequenced at Auburn University Genomic and Sequencing Laboratory.
The resulting DNA sequences were analyzed using the Vector NTI suite (Invitrogen) and blast searchs were performed against known sequences in NCBI GenBank (www.ncbi.nih.gov/Genbank). Fragments corresponding to sequences in the 3' open reading frame of genes were then further investigated for differential expression.
Real-time RT-PCR analysis: The levels of expression of the selected genes were quantified using real-time RT-PCR analysis. Briefly, total RNA was extracted from pig hippocampi as described above and reverse transcribed into cDNA using an iScript cDNA synthesis kit (BioRad Laboratories, Hercules, CA, USA). Real-time PCR was carried out in an iCycler My iQ Real-Time PCR detection System (BioRad). Oligonucleotide primers specific for the genes of interest were designed using the Vector NTI® suite software (Informax) and synthesized by Sigma-Genosys (The Woodlands, TX, USA). A list of primer sets that were designed for the selected genes is included in Table 1. The thermal cycling conditions were set at 95°C for 3 min, fllowed by 45 repeats of a two-step cycle; 10 s at 95 and 45 s at 65°C. Melting curve analysis was carried out to determine the specificity of PCR. The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and acidic ribiosomal phosphoprotein (364B) were used as internal references. A negative control without cDNA template was run simultaneously with every assay. Each sample was analyzed in duplicate.
Gene expression quantification was calculated with the method described by Vandesompele et al. (2002) and using the gene expression macro software (BioRad).
Statistical analysis of data: All values were expressed as Mean±Sem. Data were processed with one-way analysis of variance (ANOVA), correlation and regression analysis and Student’s t test for comparison of groups. The level of significance was set at p≤0.05.
| Table 1: | Primers for all genes verified by real-time PCR |
 | |
| Table 2: | Blast search results showing matches of the sequences of the isolated PCR fragments that appear to be differentially expressed after heat stress in pig hippocampi. The genes marked with the (*) are those of which the genes sequences matched the 3'open reading frames of the specified genes |
 | |
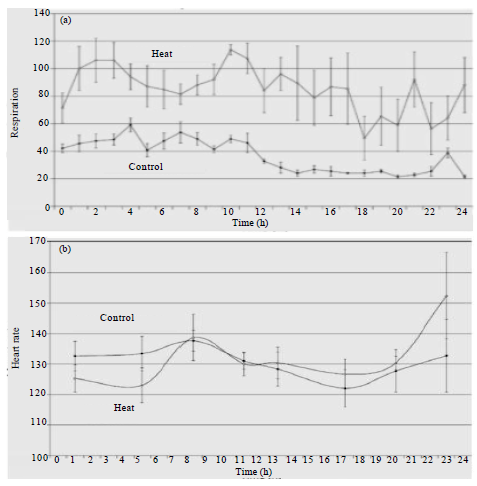 | |
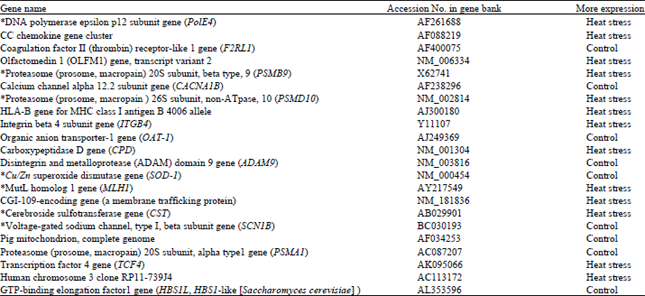
| Fig. 1(a-b): | (a) Changes in respiratory rate (breaths min-1) and (b) Changes in heart rate with time, in control and heat-stressed pigs. (a), all points from 0 to 13 h are significantly different between treatments (p = 0.05). Values at time-points from 14 to 23 h are not significantly different. (b), values at none of the time-points were significantly different between treatments |
RESULTS
Effect of heat stress on core body temperature, respiratory rate and heart rate: The average core body temperature in control animals was 102.5±0.25°F while, in heat-stressed pigs core body temperature was significantly elevated to 104.5±0.5°F.
Respiratory rate was also elevated in heat-stressed pigs, but there was no change in their heart rate. Changes in respiratory rate and heart rate over time are shown in Fig. 1(a, b).
Identity of differentially expressed bands: A total of 240 different primer combinations were used in this study and the separated cDNA fragments in 39 gels were analyzed after auto-radiography. Thirty-one differentially expressed cDNA clones either induced, or inhibited, in heat-stressed hippocampi as compared to control hippocampi. A representative image of a DD-PCR gel is shown in Fig. 2.
Blast search results: The 31 isolated differentially expressed DNA fragments were cloned, sequenced and analyzed. Blast searches against sequences in the NCBI database determined that 21 sequences matched to known human or porcine genes, one sequence matched to human genes with unknown function, four sequences matched to known genes of species other than human and pigs and five sequences had no significance similarity with any database entry. A list of genes that were included in the blast searches and that matched to human or porcine genes are shown in Table 2. Of these sequences, seven matched to the 3' open reading frame of known human genes. The 3' terminuses of the open reading frames were targeted for amplification by DD-PCR. Five of these genes were chosen for further investigation by real-time PCR.
Quantitative real-time PCR results
Effect of heat stress on gene expression after 12 h: Five genes, viz. POLE4, PSMB9, PSMD10, SOD-1 and SCN1B, were selected for verification of differential gene expression between control and heat-stressed hippocampi by real-time PCR. The samples used for this analysis were those originally used for the DD-PCR.
The differential expression of these five genes after 12 h of heat stress with DD-PCR was confirmed by real-time PCR analysis. The expression of POLE4, PSMB9 and PSMD10 was up-regulated significantly to levels 2.55±0.38, 2.7±0.6 and 2.6±0.28 times that of the control, respectively. In contrast, the expression of both SOD-1 and SCN1B was down-regulated to levels 0.45±0.1 and 0.7±0.15 times that of the control, respectively, as shown in Fig. 3.
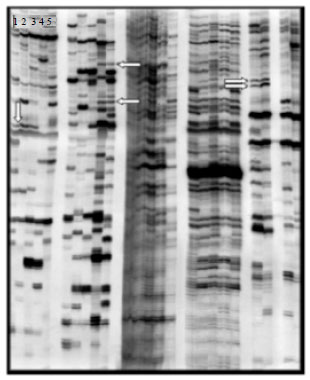 | |
| Fig. 2: | Representative autoradiograph of polyacrylamide sequencing gel, displaying PCR fragments. The lanes labeled 1, 2, and 3 contain PCR fragments representing mRNA isolated from hippocampi of control pigs. The lanes labeled 4 and 5 show PCR fragments representing mRNA from hippocampi of heat-stressed (12 h) pigs. Arrows show some differentially expressed PCR fragments between control and heat-stressed groups |
Effect of heat stress on the time course of gene expression: To determine a time course of gene expression patterns, pigs were heat-stressed for 4, 8, 12 and 24 h and RNA samples isolated from both control and heat-stressed pig hippocampi at each time point were utilized to test the expression pattern of PolE4, PSMB9, PSMD10, SOD-1 and SCN1B, using real-time PCR. We noticed that none of the 31 isolated DNA fragments differentially expressed between control and heat-stressed groups matched to any of the HSP gene family which seemed incompatible with effect of heat stress. However, the biological importance of this group of genes was previously illustrated by reports that expression of various HSP-encoding genes were altered in hippocampus following multiple stressors, including heat stress, such as HSP70 (Pavlik et al., 2003), HSP90 (Lim et al., 2003) and HSP27 (Reynolds and Allen, 2003). We therefore, designed primer sets against one HSP gene from each HSP family in order to detect their expression along with that of our genes of interest.
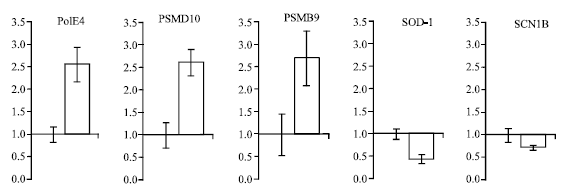 | |
| Fig. 3: | Quantitative real-time PCR analysis of gene expression in pig hippocampus after heat stress for 12 h, compared with control. Data represent Means±SEM |
We measured the expression of HSP27 (low molecular weight), HSP70 (the most important form of HSPs and of intermediate molecular weight) and HSP90 (high molecular weight) in the pig hippocampi. Relative quantification of selected genes is shown in Fig. 4(a-h).
Expression of PolyE4 was significantly inhibited (0.23±0.05) after 8 h of heat stress and was then enhanced (2.55±0.38) in response to 12 h of heat stress. The expression decreased to around basal level by 24 h.
The proteasomal subunit gene (PSMB9) showed significant enhancement of expression by heat stress at all time points, to levels 3.4±0.24, 2.69±0.27, 2.7±0.6 and 3.9±0.8 times that of the control after 4, 8, 12 and 24 h respectively. Expression of another proteasomal subunit gene (PSMD10) was also increased by heat stress; this increase was only significant at the 12 h time-point (2.6±0.28) and the expression returned to basal levels at 24 h.
Expression of the anti-oxidant gene, SOD-1, displayed significant down-regulation, to levels 0.39±0.06 and 0.45±0.1 times that of control after 8 and 12 h of heat stress. By 24 h, the expression of SOD-1 returned to its normal basal level.
The expression of the β1 subunit of voltage-gated sodium channels gene (SCN1B) was decreased by heat stress and this inhibition was significant at the 8 h time-point (0.59±0.09).
All tested HSP genes (HSP70, HSP27 and HSP90) showed minimal stimulation in response to heat stress which was non-significant, except for HSP70 after 4 h (1.67±0.3).
DISCUSSION
The present study investigated the change in porcine hippocampal gene expression profile in response to heat stress. The pigs were maintained at either 70°F (control) or 90°F (heat stressed) for 4, 8, 12, or 24 h. They were monitored hourly for changes in body temperature and respiratory rate. Both elevated body temperature and an increase in respiratory frequency are good indicators of heat-stressed animals (Gaughan et al., 2008; Huynh et al., 2005; Berman, 2005). At the end of each experiment, the hippocampi, as representative brain tissues, were excised from animals to investigate changes in gene expression patterns, using mRNA differential display RT-PCR to identify novel genes associated with heat stress in the brain of pigs. Hyperthermia, like many other stressors, triggers a reactive cellular response through activation of specific transduction pathways which leads to alterations in the expression of several gene categories that are related to either protection of cells against heat damage or to cell death (Fulda et al., 2010 Maroni et al., 2003). Changes in the expression of eight genes was confirmed and quantified by real-time PCR. These eight genes fall into four functional gene categories; viz. metabolism, ion channels, DNA replication and repair and HSPs.
DNA polymerase ∈ p12 (PolyE4) gene expression was decreased after 8 h, but showed up-regulation to more than 2.5- fold of control levels after 12 h of heat stress. PolyE4 is a structural subunit of DNA polymerase ε that contains four subunits; i.e., a large catalytic subunit A (p261) and three smaller regulatory subunits B (p59), C (p12) and D (p17) (Pospiech and Syvaoja, 2003). Polymerase ε is involved in replication of cellular DNA with both polymerase α and polymerase δ. It fulfils roles in the DNA repair synthesis step of nucleotide excision repair and base excision repair; moreover, it has been implicated in the recombination process in the cell (Takahashi et al., 2005; Rusyn et al., 2004). It has been reported that polymerase ε in adult rat brain tissues is mainly involved in different mechanisms of DNA repair, but not DNA synthesis (Raji et al., 2002; Prapurna and Rao, 1997).
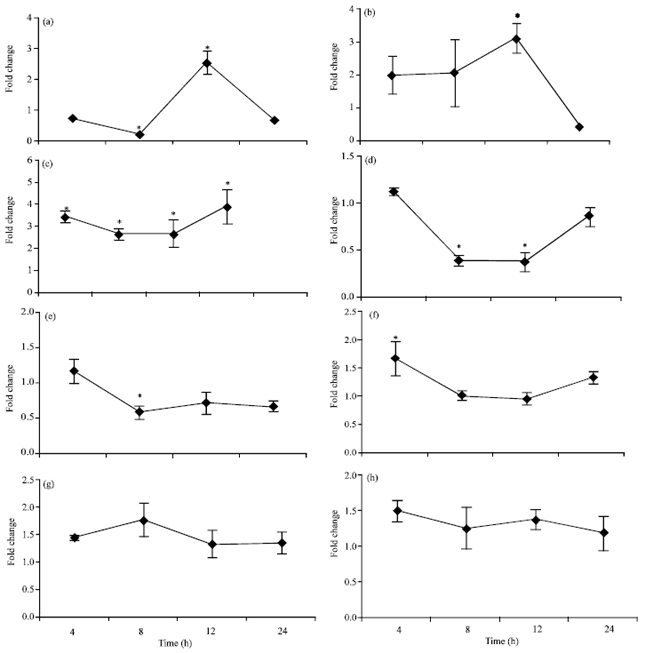 | |
| Fig. 4(a-h): | Quantitative real-time PCR analysis of expression patterns of (a) PolyE4, (b) PSMD10, (c) SMB9, (d) SOD-1, (e) SCN1B, (f) HSP70, (g) HSP27 and (h) HSP90 in pig hippocampi after heat stress for 4, 8, 12, and 24 h compared with that in control pigs. Data represent Mean±SEM *Significantly different from control at p≤0.05 |
The increase in the DNA pol ∈ p12 subunit expression observed in this study suggests up-regulated expression of DNA polymerase ε enzyme as a whole (Lee et al., 2001) which may be linked to increased DNA repair activity. Up-regulated expression of DNA repair genes at the mRNA and/or protein level has previously been reported in many conditions associated with oxidative stress like chemical-induced oxidative stress in mouse liver (Cabelof et al., 2002), transient focal ischemia and reperfusion in rat brain (Lan et al., 2003) and in blood cells of Chronic Fatigue Syndrome (CFS) patients upon exercise (Steinau et al., 2004; Chaudhuri and Behan, 2004).
Two proteasomal subunit genes were up-regulated by heat stress, viz. PSMB9 and PSMD10. These two genes represent the 20 S and 19 S subunits of the 26 S proteasome, respectively. The 26 S proteasome is a multisubunit ATPase-dependent protease which is distributed in the cytoplasmic and nuclear compartments of the cell and is made up of at least 32 different protein subunits. It is formed by the association of the 20 S proteasome, which forms the proteolytic core and the 19 S complex which is the regulatory element that caps the 20S at both ends (Bose et al., 2004; Piccinini et al., 2003). Proteasome 26 S plays a central role in the degradation of intracellular proteins. It removes abnormal proteins and mediates post-translational modification and degradation of proteins which is essential for most cellular processes, such as cell cycling, DNA repair, cell signaling, gene transcription and apoptosis (Ardley et al., 2005). Proteasomal subunits genes (PSM genes) are among the heat stress-inducible genes and are thought to be involved in tissue protection. Induction of PSM gene expression may be an adaptation to a decrease in the levels of functional proteasomes. Since direct modification of the catalytic core subunits of the proteasome inhibits its activities, the heat stress-induced accumulation of abnormal denatured proteins affects proteasomal activity and triggers an increase in expression of PSM genes which suggests an autoregulatory feedback mechanism that helps to compensate for the reduced proteasomal activity (Meiners et al., 2003; Piccinini et al., 2003).
The minimal induction of the three investigated HSP genes (HSP70, HSP90 and HSP27) that we demonstrated in this study (≤1.7 fold) at all time points may also support the above PMS gene findings. HSPs play important roles in cell protection and cell recovery from heat-induced damage through their role in preventing protein aggregation, catalyzing refolding of misfolded proteins, facilitating degradation of severely damaged proteins and playing a role in DNA damage repair (Pandita et al., 2004). Many studies have shown the induction of different HSPs at either mRNA or protein level in the hippocampus and hippocampal neorons in response to heat stress (Yao et al., 2007; Maroni et al., 2003; Bechtold and Brown, 2003; Lim et al., 2003; Reynolds and Allen, 2003). However, another study has been reported that the brain shows a milder HSP response after heat stress than do other body tissues and that, among the different brain areas, the hippocampus shows the weakest response (Maroni et al., 2003) which may explain the vulnerability of the hippocampus to heat insults (Sharma et al., 1994). Hippocampal neurons were unable to induce a heat shock response after heat stress, due to a lack of heat shock factor-1 (HSF-1) which acts as a transcription factor for HSP genes by interacting with a Heat Shock Element (HSE) in the promoter region of these genes (Marcuccilli et al., 1996). Hippocampal neurons can express heat shock factor-2 (HSF-2) instead which thought to explain the limited expression of HSPs in response to heat stress (Kaarniranta et al., 2002). Additionally, some studies attributed the disruption in heat shock response to the enhancement of proteasomal activity as a result of heat stress which in turn accelerates the turnover of denatured proteins and failure to activate HSFs (Awasthi and Wagner, 2005; Rattan, 2004; Friant et al., 2003); this supports our findings on the stimulation of some proteasomal subunits by heat stress.
The superoxide dismutase-1 gene (Cu/Zn SOD or SOD-1) was significantly inhibited after heat stress. SOD-1 is one of the key antioxidant enzymes that catalyzes the dismutation of superoxide radical (O2¯) to H2O2 (Peluffo et al., 2005). It is the cytosolic isozyme of a group of antioxidant metalloenzymes, superoxide dismutases which are involved in O2¯ detoxification in normal cellular metabolism and after cell injury. Many stressors and pathological conditions that are associated with accumulation of Reactive Oxygen Species (ROS) and enhanced oxidative stress have been reported to reduce the level of SOD-1 mRNA, with or without lowering the protein level and/or enzymatic activity in different tissues and different species. These include conditions like chemical-induced excitotoxic injury (Peluffo et al., 2005), hypoxia/ischemia (Martin et al., 2002; Rawal et al., 2004), cold stress (Niakao et al., 1999), immobilization stress (Oishi and Machida, 2002), chronic fatigue syndrome (Steinau et al., 2004), septic shock (Ghosh et al., 1996), type-II diabetes (Sakuraba et al., 2002), viral infection (Hosakote et al., 2011) and inflammatory bowl disease (Segui et al., 2004). In the CNS, SOD-1 plays an important role in neuroprotection and it has been reported that SOD-1 is rapidly downregulated after several types of CNS injuries (Peluffo et al., 2005). Hyperthermia induces CNS ischemia which is associated with increased production of ROS (Zhao et al., 2006; Perez-Crespo et al., 2005; Niu et al., 2003). Down-regulation in SOD-1 expression could be a consequence of increased oxidative stress (Ghosh et al., 2007; Li et al., 2004). One other possible mechanism for heat-induced downregulation of SOD-1 is that heat stress inhibits the activation and translocation of transcription factor NF-êB and generally blocks the expression of NF-êB-dependent targets (Janus et al., 2011; Kitamura et al., 2006; Chen et al., 2004) like the SOD-1(Milani et al., 2011; Rojo et al., 2004).
Heat stress did not reduce only the expression of the SOD-1 but also down-regulated the β1s ubunit of the voltage-gated sodium channel gene (SCN1B) in the pig hippocampus. The voltage-gated sodium channels (VGSCs) are membrane proteins that play an important role in electrical signaling by generation and propagation of action potentials in excitable tissues, like the central and peripheral nervous systems and muscles, by mediating the rapid entry of Na+ ions into excitable cells upon voltage changes across the plasma membrane. The VGSCs are heteromultimeric proteins that consist of a pore-forming α-subunit which is responsible for the voltage-sensitive characteristics of the channel and auxiliary β-subunits which act as regulators of the α-subunit. The β1-subunit alters gating, voltage sensitivity and recovery from fast inactivation of VGSCs (Qin et al., 2003; Takahashi et al., 2003). Down-regulation of the expressionof genes encoding VGSCs subunits has been reported in the brain following oxidative insults and febrile seizures. This down-regulation of ion channel subunits may play a key role in neurophysiological recovery from brain injury (Yin et al., 2013; Lu et al., 2004). Down-regulation of the β1 accessory subunit is suggested to be a compensatory mechanism by which brain cells down-modulate VGSCs to reduce sodium influx and preserve the inward sodium gradient and diminish the cellular energy demand which decreases neuronal excitability and provides neuroprotection against stressful conditions (Fung, 2000; Obrenovitch, 1998).
In summary, the expression of five genes was confirmed to be altered in pig hippocampus in response to heat stress of 4, 8 12 and 24 h. Most of these genes are involved in cellular protection mechanisms against stress-induced protein and DNA damage and in reduction of cellular energy demand. The down-regulation of the SOD-1 may represent a cause of heat-induced reduction in the antioxidant capacity of the brain which could be a reason for brain injury after heat stress. A genomic approach, such as that used here, could not only yield novel insight into the molecular changes that area induced in the brain in response to heat stress, but may also indicate therapeutic targets and new strategies for the treatment of heat-related illnesses.
ACKNOWLEDGEMENT
We would like to thank Dr. H. Cheng, Purdue University, IN, USA, for helping with all pig experiments, Dr. N. Deraz, National Research Center, Cairo, Egypt, for writing assistance and critical revision of the study, Dr. C. Rogers, Dr. H. Gray-Edwards, Mrs. T. Albrecht-Schmitt, Auburn University, AL, USA and Dr. M. Hassan, Animal Care and Research Center, King Saud University, KSA, for their technical assistance. We also greatly appreciate the financial support of the Office of Naval Research, VA, USA (grant No. NAVY-N00014-03-10293). Author would like to extend their sincere appreciation to the Deanship of Scientific Research at King Saud University, Riyadh, KSA for funding the work through the research group project No. RGP-VPP-306 to Dr. O. Abd-Elkader.
REFERENCES
- Alm, P., H.S. Sharma, P.O. Sjoquist and J. Westman, 2000. A new antioxidant compound H-290/51 attenuate nitric oxide synthase and heme oxygenase expression following hyperthrmic brain injury. Amino Acids, 19: 383-394.
PubMedDirect Link - Ardley, H.C., C.C. Hung and P.A. Robinson, 2005. The aggravating role of the ubiquitin-proteasome system in neurodegeneration. FEBS Lett., 579: 571-576.
CrossRef - Awasthi, N. and B.J. Wagner, 2005. Upregulation of heat shock protein expression by proteasome inhibition: An antiapoptotic mechanism in the lens. Invest. Ophthalmol. Visual Sci., 46: 2082-2091.
CrossRef - Basu, R., 2009. High ambient temperature and mortality: A review of epidemiologic studies from 2001 to 2008. Environ. Health, 8: 40-52.
Direct Link - Bechtold, D.A. and I.R. Brown, 2003. Induction of Hsp27 and Hsp32 stress proteins and vimentin in glial cells of the rat hippocampus following hyperthermia. Neurochem. Res., 28: 1163-1173.
CrossRef - Bergeron, M., C. Devore and S. Rice, 2011. Policy statement-Climatic heat stress and exercising children and adolescents. Pediatrics, 28: e741-e747.
Direct Link - Berman, A., 2005. Estimates of heat stress relief needs for Holstein dairy cows. J. Anim. Sci., 83: 1377-1384.
CrossRefDirect Link - Borges, J.C. and C.H. Ramos, 2005. Protein folding assisted by chaperones. Protein Peptide Lett., 12: 257-261.
CrossRef - Bose, S., F.L.L. Stratford, K.I. Broadfoot, G.G.F. Mason and A.J. Rivett, 2004. Phosphorylation of 20 S proteasome alpha subunit C8 (α7) stabilizes the 26S proteasome and plays a role in the regulation of proteasome complex by γ-interferon. Biochem. J., 378: 177-184.
Direct Link - Cabelof, D.C., J.J. Raffoul, S. Yanamadala, Z. Guo and A.R. Heydari, 2002. Induction of DNA polymerase β-dependent base excision repair in response to oxidative stress in vivo. Carcinogenesis, 23: 1419-1425.
CrossRef - Chaudhuri, A. and P.O. Behan, 2004. In vivo magnetic resonance spectroscopy in chronic fatigue syndrome. Prostaglandins Leukotrienes Essential Fatty Acids, 71: 181-183.
CrossRef - Chen, Y., A.P. Arrigo and R.W. Currie, 2004. Heat shock treatment suppresses angiotensin II-induced activation of NF-κB pathway and heart inflammation: A role for IKK depletion by heat shock? Am. J. Physiol. Heart Circulatory Physiol., 287: H1104-H1114.
PubMedDirect Link - Chia, S.E. and K.J. Teo, 2003. Prognosis of adult men with heat exhaustion with regard to postural stability and neurobehavioral effects: A 6-month follow-up study. Neurotoxicol. Teratol., 25: 503-508.
CrossRef - Chou, Y.T., S.T. Lai, C.C. Lee and M.T. Lin, 2003. Hypothermia attenuates circulatory shock and cerebral ischaemia in experimental heatstroke. Shock, 19: 388-393.
PubMed - Coris, E.E., A.M. Ramirez and D.J. van Durme, 2004. Heat illness in athletes. Sports Med., 34: 9-16.
PubMed - Fountoulakis, M., G.T. Tsangaris, A. Maris and G. Lubec, 2005. The rat brain hippocampus proteome. J. Chromatogr. B, 819: 115-129.
CrossRef - Franklin, T.B., A.M. Krueger-Naug, D.B. Clarke, A.P. Arrigo and R.W. Currie, 2005. The role of heat shock proteins Hsp70 and Hsp27 in cellular protection of the central nervous system. Int. J. Hyperthermia, 21: 379-392.
CrossRef - Friant, S., K.D. Meier and H. Riezman, 2003. Increased ubiquetin-dependent degradation can replace the essential requirement for heat shock protein induction. EMBO J., 22: 3783-3791.
PubMed - Fulda, S., A.M. Gorman, O. Hori and A. Samali, 2010. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol.
CrossRefDirect Link - Fung, M.L., 2000. Role of voltage-gated Na+ channels in hypoxia-induced neuronal injuries. Clin. Exp. Pharmacol. Physiol., 27: 569-574.
CrossRefPubMedDirect Link - Garrido, C., S. Gurbuxani, L. Ravagnan and G. Kroemer, 2001. Heat shock proteins: Endogenous modulators of apoptotic cell death. Biochem. Biophys. Res. Commun., 286: 433-442.
CrossRefDirect Link - Gaughan, J.B., T.L. Mader, S.M. Holt and A. Lisle, 2008. A new heat load index for feedlot cattle. J. Anim. Sci., 86: 226-234.
PubMed - Ghosh, B., C.D. Hanevold, K. Dobashi, J.K. Orak and I. Singh, 1996. Tissue differences in antioxidant enzyme gene expression in response to endotoxin. Free Radical Biol. Med., 21: 533-540.
CrossRef - Ghosh, G., K. De, S. Maity, D. Bandyopadhyay, S. Bhattacharya, R.J. Reiter and A. Bandyopadhyay, 2007. Melatonin protects against oxidative damage and restores expression of GLUT4 gene in the hyperthyroid rat heart. J. Pineal Res., 42: 71-82.
CrossRef - Gupta, M., S. Kumar, S.S. Dangi and B.L. Jangir, 2013. Physiological, biochemical and molecular responses to thermal stress in goats. Int. J. Livestock Res., 3: 27-38.
Direct Link - Hermesh, H., R. Shiloh, Y. Epstein, H. Manaim, A. Weizman and H. Munitz, 2000. Heat intolerance in patients with chronic schizophrenia maintained with antipsychotic drugs. Am. J. Psychiatry, 157: 1327-1329.
CrossRef - Hosakote, Y.M., P.D. Jantzi, D.L. Esham, H. Spratt, A. Kurosky, A. Casola and R.P. Garofalo, 2011. Viral-mediated inhibition of antioxidant enzymes contributes to the pathogenesis of severe respiratory syncytial virus bronchiolitis. Am. J. Respir. Crit. Care Med., 183: 1550-1560.
CrossRef - Huynh, T.T.T., A.J.A. Aarnink, M.W.A. Verstegen, W.J.J. Gerrits, M.J.W. Heetkamp, B. Kemp and T.T. Canh, 2005. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci., 83: 1385-1396.
Direct Link - Kaarniranta, K., N. Oksala, H.M. Karjalainen, T. Suuronen and L. Sistonen et al., 2002. Neuronal cells show regulatory differences in the hsp70 gene response. Mol. Brain Res., 101: 136-140.
CrossRef - Kitamura, C., T. Nishihara, Y. Ueno, K.K. Chen, T. Morotomi and J. Yano et al., 2006. Effects of sequential exposure to lipopolysaccharide and heat stress on dental pulp cells. J. Cell. Biochem., 99: 797-806.
CrossRefDirect Link - Kysely, J., 2004. Mortality and displaced mortality during heat waves in the Czech republic. Int. J. Biometeorol., 49: 91-97.
CrossRefDirect Link - Lan, J., W. Li, F. Zhang, F.Y. Sun, T. Nagayama, C. O'Horo and J. Chen, 2003. Inducible repair of oxidative DNA lesions in the rat brain after transient focal ischemia and reperfusion. J. Cerebral Blood Flow Metab., 23: 1324-1339.
CrossRef - Le Grevès, P., H.S. Sharma, J. Westman, P. Alm and F. Nyberg, 1997. Acute heat stress induces edema and nitric oxide synthase upregulation and down-regulates mRNA levels of the NMDAR1, NMDAR2A and NMDAR2B subunits in the rat hippocampus. Acta Neurochirurgica Supplements, 70: 275-278.
CrossRef - Lee, B.P.L., W.J. Rushlow, C. Chakraborty and P.K. Lala, 2001. Differential gene expression in premalignant human trophoblast: Role of IGFBP‐5. Int. J. Cancer, 94: 674-684.
CrossRefDirect Link - Li, H., Y. Gu, Y. Zhang, M.J. Lucas and Y. Wang, 2004. High glucose levels down-regulate glucose transporter expression that correlates with increased oxidative stress in placental trophoblast cells in vitro. J. Soc. Gynecol. Invest., 11: 75-81.
CrossRefDirect Link - Liang, P., W. Zhu, X. Zhang, Z. Guo and R.P. O'Connell et al., 1994. Differential display using one-base anchored oligo-dT primers. Nucleic Acids Res., 22: 5763-5764.
Direct Link - Lim, M.C., S.M. Brooke and R.M. Sapolsky, 2003. gp120 neurotoxicity fails to induce heat shock defenses, while the over expression of hsp70 protects against gp120. Brain Res. Bull., 61: 183-188.
CrossRef - Lu, X.C., A.J. Williams, C. Yao, R. Berti and J.A. Hartings et al., 2004. Microarray analysis of acute and delayed gene expression profile in rats after focal ischemic brain injury and reperfusion. J. Neurosci. Res., 77: 843-857.
CrossRefDirect Link - Lugo-Amador, N.M., T. Rothenhaus and P. Moyer, 2004. Heat-related illness. Emerg. Med. Clin. N. Am., 22: 315-327.
PubMed - Marcuccilli, C.J., S.K. Mathur, R.I. Morimoto and R.J. Miller, 1996. Regulatory differences in the stress response of hippocampal neurons and glial cells after heat shock. J. Neurosci., 16: 478-485.
Direct Link - Maroni, P., P. Bendinelli, L. Tiberio, F. Rovetta, R. Piccoletti and L. Schiaffonati, 2003. In vivo heat-shock response in the brain: Signalling pathway and transcription factor activation. Mol. Brain Res., 119: 90-99.
CrossRef - Martin, R., G. Fitzl, C. Mozet, H. Martin, K. Welt and E. Wieland, 2002. Effect of age and hypoxia/reoxygenation on mRNA expression of antioxidative enzymes in rat liver and kidneys. Exp. Gerontol., 37: 1481-1487.
CrossRefDirect Link - McEwen, B.S. and P.J. Gianaros, 2010. Central role of the brain in stress and adaptation: links to socioeconomic status, health and disease. Ann. N. Y. Acad. Sci., 1186: 190-222.
CrossRefDirect Link - Meiners, S., D. Heyken, A. Weller, A. Ludwig, K. Stangl, P.M. Kloetzel and E. Kruger, 2003. Inhibition of proteasome activity induces concerted expression of proteasome genes and de novo formation of mammalian proteasomes. J. Biol. Chem., 278: 21517-21525.
CrossRefDirect Link - Milani, P., S. Gagliardi, E. Cova and C. Cereda, 2011. SOD1 transcriptional and posttranscriptional regulation and its potential implications in ALS. Neurol. Res. Int., (In Press).
CrossRefDirect Link - Mountjoy, M., J.M. Alonso, M.F. Bergeron, J. Dvorak, S. Miller and S. Migliorini, 2012. Hyperthermic-related challenges in aquatics, athletics, football, tennis and triathlon. Br. J. Sports Med., 46: 800-804.
CrossRef - Niakao, C., T. Ookawara, T. Kizaki, K. Suzuki, S. Haga, Y. Sato and H. Ohno, 1999. Effects of acute cold stress on mrna expression and immunoreactivity of three superoxide dismutase isoenzymes in genetically obese mice. Res. Commun. Mol. Pathol. Pharmacol., 106: 47-61.
PubMed - Niu, K.C., K.C. Lin, C.Y. Yang and M.T. Lin, 2003. Protective effects of α‐tocopherol and mannitol in both circulatory shock and cerebral ischaemia injury in rat heatstroke. Clin. Exp. Pharmacol. Physiol., 30: 745-751.
CrossRef - Obrenovitch, T.P., 1998. Neuroprotective strategies: Voltage-gated Na+-channel down-modulation versus presynaptic glutamate release inhibition. Rev. Neurosci., 9: 203-212.
CrossRef - Oishi, K. and K. Machida, 2002. Different effects of immobilization stress on the mRNA expression of antioxidant enzymes in rat peripheral organs. Scand. J. Clin. Lab. Invest., 62: 115-121.
CrossRef - Pandita, T.K., R. Higashikubo and C.R. Hunt, 2004. HSP70 and genomic stability. Cell Cycle, 3: 591-592.
Direct Link - Pavlik, A., I.S. Aneja, J. Lexa and B.A. Al-Zoabi, 2003. Identification of cerebral neurons and glial cell types inducing heat shock protein Hsp70 following heat stress in the rat. Brain Res., 973: 179-189.
CrossRefPubMedDirect Link - Peluffo, H., L. Acarin, M. Faiz, B. Castellano and B. Gonzalez, 2005. Cu/Zn superoxide dismutase expression in the postnatal rat brain following an excitotoxic injury. J. Neuroinflammation, Vol. 2.
CrossRefDirect Link - Perez-Crespo, M., M.A. Ramirez, R. Fernandez-Gonzalez, D. Rizos, P. Lonergan, B. Pintado and A. Gutierrez-Adan, 2005. Differential sensitivity of male and female mouse embryos to oxidative induced heat-stress is mediated by glucose-6-phosphate dehydrogenase gene expression. Mol. Reprod. Dev., 72: 502-510.
CrossRefDirect Link - Piccinini, M., M. Mostert, S. Croce, S. Baldovino, M. Papotti and M.T. Rinaudo, 2003. Interferon-γ-inducible subunits are incorporated in human brain 20 S proteasome. J. Neuroimmun., 135: 135-140.
CrossRef - Pospiech, H. and J.E. Syvaoja, 2003. DNA polymerase ε-more than a polymerase. Sci. World J., 3: 87-104.
CrossRefPubMedDirect Link - Prapurna, D.R. and K.S. Rao, 1997. DNA polymerases δ and ε in developing and aging rat brain. Int. J. Dev. Neurosci., 15: 67-73.
CrossRefPubMedDirect Link - Qin, N., M.R. D'Andrea, M.L. Lubin, N. Shafaee, E.E. Codd and A.M. Correa, 2003. Molecular cloning and functional expression of the human sodium channel β1B subunit, a novel splicing variant of the β1 subunit. Eur. J. Biochem., 270: 4762-4770.
CrossRef - Raji, N.S., T. Hari Krishna and K.S. Rao, 2002. DNA-polymerase α, β, δ and ε activities in isolated neuronal and astroglial cell fractions from developing and aging rat cerebral cortex. Int. J. Dev. Neurosci., 20: 491-496.
CrossRef - Rattan, S.I., 2004. Mechanisms of hormesis through mild heat stress on human cells. Ann. New York Acad. Sci., 1019: 554-558.
CrossRef - Rawal, A.K., M.G. Muddeshwar and S.K. Biswas, 2004. Rubia cordifolia, Fagonia cretica linn and Tinospora cordifolia exert neuroprotection by modulating the antioxidant system in rat hippocampal slices subjected to oxygen glucose deprivation. BMC Complement. Altern. Med., Vol. 4.
CrossRefDirect Link - Reynolds, L.P. and G.V. Allen, 2003. A review of heat shock protein induction following cerebellar injury. Cerebellum, 2: 171-177.
CrossRefDirect Link - Rojo, A., M. Salinas, D. Martin, R. Perona and A. Cuadrado, 2004. Regulation of Cu/Zn-superoxide dismutase expression via the phosphatidylinositol 3 kinase/Akt pathway and nuclear factor-kappaB. J. Neurosci., 24: 7324-7334.
Direct Link - Rusyn, I., S. Asakura, B. Pachkowski, B.U. Bradford and M.F. Denissenko et al., 2004. Expression of base excision DNA repair genes is a sensitive biomarker for in vivo detection of chemical-induced chronic oxidative stress identification of the molecular source of radicals responsible for DNA damage by peroxisome proliferators. Cancer Res., 64: 1050-1057.
CrossRef - Sakuraba, H., H. Mizukami, N. Yagihashi, R. Wada, C. Hanyu and S. Yagihashi, 2002. Reduced beta-cell mass and expression of oxidative stress-related DNA damage in the islet of Japanese type II diabetic patients. Diabetologia, 45: 85-96.
CrossRefPubMedDirect Link - Segui, J., M. Gironella, M. Sans, S. Granell and F. Gil et al., 2004. Superoxide dismutase ameliorates TNBS-induced colitis by reducing oxidative stress, adhesion molecule expression and leukocyte recruitment into the inflamed intestine. J. Leukocyte Biol., 76: 537-544.
CrossRefDirect Link - Sharma, H.S., J. Westman, F. Nyberg, C. Zimmer, J. Cervos-Navarro and P.K. Dey, 1994. Selective Vulnerability of Rat Hippocampus in Heat Stress. In: Temperature Regulation: Recent Physiological and Pharmacological Advances, Milton, A.S. (Ed.). Birkhauser Basel, USA., ISBN:978-3-0348-9646-7, pp: 267-272.
- Sharma, H.S. and P.J. Hoopes, 2003. Hyperthermia induced pathophysiology of the central nervous system. Int. J. Hyperthermia, 19: 325-354.
Direct Link - Sharma, H.S., K. Drieu, P. Alm and J. Westman, 2000. Role of nitric oxide in Blood-brain Barrier permeability, brain edema and cell damage following hyperthermic brain injury. An experimental study using EGB-761 and Gingkolide B pretreatment in the rat. Acta Neurochir., 76: 81-86.
CrossRefDirect Link - Sherman, M.Y. and A.L. Goldberg, 2001. Cellular defense against unfolded proteins: A cell biologist thinks about neurodegenerative diseases. Neuron, 29: 15-32.
PubMed - Sonna, L.A., J. Fujita, S.L. Gaffin and C.M. Lilly, 2002. Invited review: Effects of heat and cold stress on mammalian gene expression. J. Applied Physiol., 92: 1725-1742.
PubMedDirect Link - Steinau, M., E.R. Unger, S.D. Vernon, J.F. Jones and M.S. Rajeevan, 2004. Differential-display PCR of peripheral blood for biomarker discovery in chronic fatigue syndrome. J. Mol. Med., 82: 750-755.
CrossRef - Takahashi, N., S. Kikuchi, Y. Dai, K. Kobayashi, T. Fukuoka and K. Noguchi, 2003. Expression of auxiliary β subunits of sodium channels in primary afferent neurons and the effect of nerve injury. Neuroscience, 121: 441-450.
CrossRef - Takahashi, Y., S.I. Moriwaki, Y. Sugiyama, Y. Endo and K. Yamazaki et al., 2005. Decreased gene expression responsible for post-ultraviolet DNA repair synthesis in aging: A possible mechanism of age-related reduction in DNA repair capacity. J. Invest. Dermatol., 124: 435-442.
CrossRef - Vandesompele, J., K. De Preter, F. Pattyn, B. Poppe, N. van Roy, A. De Paepe and F. Speleman, 2002. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol., Vol. 3.
CrossRef - Yao, S., M. Peng, X. Zhu, M. Cheng and X. Qi, 2007. Heat shock protein72 protects hippocampal neurons from apoptosis induced by chronic psychological stress. Int. J. Neurosci., 117: 1551-1564.
CrossRef - Yin, Y.H., N. Ahmad and M. Makmor-Bakry, 2013. Pathogenesis of epilepsy: Challenges in animal models. Iran J. Basic Med. Sci., 16: 1119-1132.
Direct Link - Zhao, Q.L., Y. Fujiwara and T. Kondo, 2006. Mechanism of cell death induction by nitroxide and hyperthermia. Free Radical Biol. Med., 40: 1131-1143.
CrossRef