
Fang-Xin Zhang
Department of Gastroenterology, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Zhi-Yun Deng
Department of Test, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Ju-Zi Dong
Department of Test, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Yun Duan
Department of Gastroenterology, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Shang-Xin Deng
Department of Gastroenterology, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Jia-Yu Chen
Department of Gastroenterology, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Qiang Ma
Department of Gastroenterology, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
Ti-Dong Shan
Department of Gastroenterology, General Hospital of Chinese PLA, Lanzhou Command, Lanzhou, 730050, China
International Journal of Pharmacology
Year: 2013 | Volume: 9 | Issue: 6 | Page No.: 385-389







ABSTRACT
This study is aimed to investigate the effects of prebiotics on the expression of occludin and cytokines in the intestinal mucosal barrier of stressed rats in a high altitude environment. Forty-eight rats were acutely stressed in a high altitude hypoxia environment (3848 m). Rats in the hypoxia control group (n = 24) and prebiotics group (n = 24) were stressed for 2, 4 and 6 days before harvesting the intestinal mucosa. The expression of the occludin protein in the intestinal mucosa was analyzed by immunohistochemistry and tumor necrosis factor-α (TNF-α) and interleukin-10 (IL-10) expression were measured by the enzyme-linked immune sorbent assay in ileum tissue. The high altitude hypoxia environment significantly reduced occludin protein and mRNA expression in the control group, but prebiotics treatment elevated its expression on the 4th day and 6th day. Prebiotics treatment also significantly alleviated changes in TNF-α and IL-l0 induced by high altitude hypoxic stress. Prebiotics may help to maintain the integrity of epithelial cells by regulating expression of the occludin protein and cytokines.
PDF Abstract XML References Citation
Received: July 15, 2013;
Accepted: January 24, 2014;
Published: March 18, 2014
How to cite this article
Fang-Xin Zhang, Zhi-Yun Deng, Ju-Zi Dong, Yun Duan, Shang-Xin Deng, Jia-Yu Chen, Qiang Ma and Ti-Dong Shan, 2013. Effects of Prebiotics on the Expression of the Integral Membrane Protein
Occludin and Cytokines in Rats after Acute High Altitude Exposure. International Journal of Pharmacology, 9: 385-389.
DOI: 10.3923/ijp.2013.385.389
URL: https://scialert.net/abstract/?doi=ijp.2013.385.389
DOI: 10.3923/ijp.2013.385.389
URL: https://scialert.net/abstract/?doi=ijp.2013.385.389
INTRODUCTION
The major environmental features of high altitude areas are low pressure, low temperature, low humidity and high sunshine exposure and these features may have adverse effects on human health (Korzeniewski, 2008). For example, high altitude environments are associated with a series of digestive abnormities, such as loss of appetite, nausea, vomiting, diarrhea and in the worst case, intestinal endotoxemia (IETM) (Abellan et al., 2005). Under high altitude stress, ischemia and hypoxia occur in the gastrointestinal tract before other organs and these changes are restored in the gastrointestinal tract later than in other organs, indicating that it is highly sensitive to this stress. Persistent stress can induce intestinal mucosal damage and compromise intestinal barrier function and immunity, which may lead to invasion of intestinal bacteria and bacterial endotoxins and even result in IETM.
The intestinal epithelium is a single-cell layer that constitutes the largest and most important barrier against external environment. Epithelium maintains its selective barrier function through the formation of complex protein-protein networks that mechanically link adjacent cells and seal the intercellular space. The protein-protein networks that connect epithelial cells form three adhesive complexes: Desmosomes, adherens junctions and tight junctions. These networks allow the intestinal epithelium to act as a selectively permeable barrier, permitting the absorption of nutrients, electrolytes and water while maintaining an effective defense against intraluminal toxins, antigens and enteric flora.
Occludin is a 65 kDa, type II transmembrane protein with a multidomain tetraspan structure and 90% homology among mammals (Ando-Akatsuka et al., 1996). Its extended C terminus has been shown to be essential for occludin to interact with zonula occludens-1 (ZO-1), subsequently mediating its intracellular trafficking to the plasma membrane tight junction site. Occludin is a constitutive molecule of tight junctions and it is involved in regulating the permeability of intestinal mucosal cells (Chen et al., 2002). Investigations have confirmed its roles for the extracellular loops and at least one transmembrane domain in occludin localization and tight junction stability (Cummins, 2012). We previously found that acute high altitude exposure led to the breakage of tight junctions in rat intestinal epithelial cells using electron microscopy (Wu et al., 2010). However, it remains unknown how high altitude exposure affects tight junctions and occludin expression.
Prebiotics are nondigestible food ingredients, which stimulate the growth and/or activity of one bacterium or a group of bacteria in the large bowel and they have a beneficial effect on the health of the host (Schrezenmeir and de Vrese, 2001). Prebiotics represents a very promising treatment for certain infections and inflammatory-associated bowel disorders. Previous studies suggest that prebiotics contribute to (1) Growth of the intestinal mucosa, (2) Function of the intestinal mucosal barrier and balance of body fluids and electrolytes, (3) Supplementation of body with energy and nutritional agents and (4) Regulation of defense against pathogenic microorganisms and antibody-mediated immunity (Ioannidis et al., 2011). Our study aimed to investigate the effects of prebiotics treatment on occludin and cytokine expression in a high altitude rat model.
MATERIALS AND METHODS
Reagents: Polyclonal occludin antibody was obtained from Abcam (UK). The 3, 3'-diaminobenzidine (DAB) kit and enzyme-linked immune sorbent assay (ELISA) kits for tumor necrosis factor-α (TNF-α) and interleukin-10 (IL-10) were obtained from GSGB-Bio (Beijing). Galacto-oligosaccharide (GOS) was obtained from Amresco (USA).
Animals: Forty-eight 6- to 7-week-old healthy adult Wistar rats (24 male and 24 female) with an average body weight of 200±8 g were provided by the Experimental Animal Center of Lanzhou University School of Medicine (altitude 1520 m). Before the experiment, the rats were fed in an environment at a temperature of 23°C and humidity of 50% for one week before being transported to Lanzhou Maxianshan Radar Station (altitude 3848 m) within 12 h. The 48 rats were randomly divided into two groups: Control and treated. Each group was subdivided to three subgroups, which were sacrificed after 2, 4 and 6 days, respectively. The control group was given a normal diet and the treated group was given 1 g GOS (AMRESCO Inc. USA) prebiotics by gavage in addition to the normal diet.
Sampling: After 2, 4 and 6 days in the high altitude environment, the rats were anaesthetized using chloral hydrate (0.3 mL 100 g-1). The ileum tissue was isolated and washed in saline buffer. A 1 cm piece of tissue was fixed in neutral 10% formalin. Another 1 cm piece of ileum tissue was kept in liquid nitrogen for ELISA.
| Table 1: | Primer sequences used in qPCR |
 | |
Immunochemistry: A two-step immunochemistry method was performed. Paraffin sections were dewaxed and incubated with hydrogen peroxide to block endogenous peroxidase activity. Pepsin K was applied following the incubation with anti-occludin antibody (1:100) at 4°C overnight. After incubation with the secondary antibody, the DAB method was performed to develop color before dehydration and sealing. The images were acquired and the expression level of occludin was analyzed by quantitative image processing software (Image-Pro Plus 6 analysis software).
ELISA: The tissue that was stored in liquid nitrogen was ground and put in 0.4 mL of ice cold Phosphate Buffered Saline (PBS) and centrifuged at 4°C for 10 min. The supernatant (0.1 mL) was used to determine the expression of TNF-α and IL-10 by ELISA.
Quantitative real-time polymerase chain reaction (qPCR): The cDNA was isolated using Trizol™ and complementary DNA (cDNA) was obtained using a PrimeScriptTM RT Reagent Kit (Takara, Japan). Reactions were performed on an ABI 7300 Fast Real-time PCR System (Applied Biosystems, USA). The primers are shown in Table 1. The relative expression of messenger RNA (mRNA) was calculated using 2-ΔΔCt methods.
Statistics: All data were processed using SPSS 11.5 and expressed as the Mean±standard deviation. The χ2 test and one-way Analysis of Variance (ANOVA) were used to compare the expression levels of proteins between the control and prebiotics groups. p<0.05 was defined as statistical significance.
RESULTS
Occludin expression: In the high altitude stressed control group, the expression of occludin in the intestinal mucosal epithelial cell membranes was reduced in the 4 and 6 day groups compared to the 2 day group. This result indicates that persistent high altitude stress inhibits the expression of occludin (Fig. 1a-c).
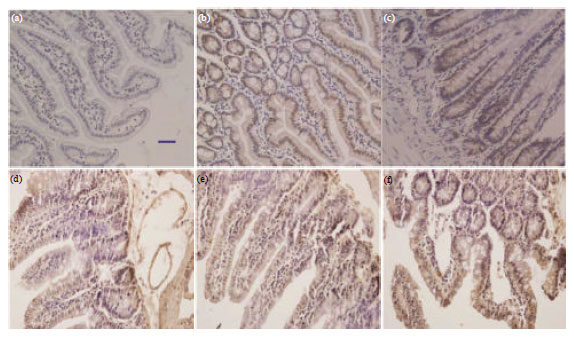 | |
| Fig. 1(a-f): | Immunochemical detection of occludin expression rats were exposed to a high altitude environment for 2, 4, or 6 days. Immunochemistry (Avidin-Biotin Complex method) was performed to examine the occludin expression in the control (a, b and c) and prebiotics (d, e and f) groups |
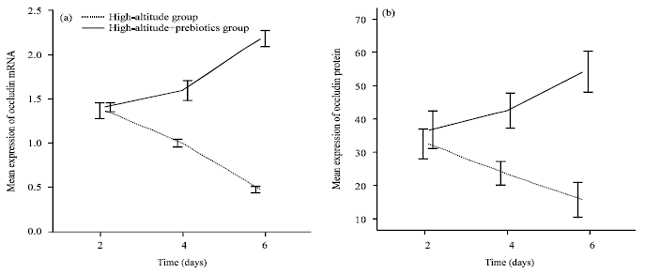 | |
| Fig. 2(a-b): | Expression of occludin mRNA and protein. The mean expression of (a) Occludin mRNA and (b) Protein with 95% confidence intervals in ileum tissue from rats stressed by high altitude and high altitude plus prebiotics treatment. Protein expression was measured by imaging processing software and mRNA was determined by qPCR. *indicates p<0.05, compared to the high altitude group |
Administration of prebiotics significantly enhanced the expression of occludin, particularly strongly at 4 days and 6 days (Fig. 1d-f). Consistent with the changes in occludin protein expression, the mRNA expression of occludin was also significantly reduced in the control group but was increased in the prebiotics group at 4 and 6 days compared to the level at 2 days (Fig. 2).
Expression of TNF-α and IL-10: High altitude stress induced TNF-α expression and reduced IL-10 expression in the ileum tissue.
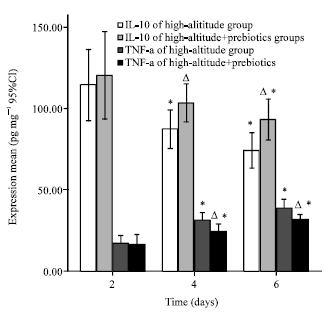 | |
| Fig. 3: | Changes in IL-10 and TNF-α expression caused by high altitude and high altitude plus prebiotics treatment. IL-10 and TNF-α expression levels were determined by ELISA compared to the corresponding high altitude groups, p<0.05. *compared to the 2-day treatment value in the same group, p<0.05 |
However, prebiotics treatment relieved the increase of TNF-α and prevented the marked decrease of IL-10 induced by the high altitude stress (Fig. 3).
DISCUSSION
Our results demonstrate that high altitude exposure reduced occludin expression but prebiotics treatment restored the occludin expression. These results indicate that loss of occludin expression may contribute to the compromised tight junction function induced by high altitude exposure and that prebiotics could restore occludin expression.
Prebiotics treatment reversed the induction of the proinflammatory cytokine TNF-α and the downregulation of the anti-inflammatory IL-10, indicating that prebiotics have an anti-inflammatory effect. This result could be explained by the following (1) Prebiotics do not undergo gastrointestinal hydrolysis, so they can stimulate the digestive and gastrointestinal hormone secretion to expel harmful bacteria and (2) Prebiotics can promote the growth of beneficial flora and they can form a layer of biological protective film to reduce the adhesion of endotoxins and inflammatory cytokines.
Previous studies showed that occludin is essential for intestinal epithelial barrier function. Pathologic bacteria can adhere to intestinal epithelial cells and disrupt the tight junction function in the barrier (Baumgart and Dignass, 2002; Blikslager et al., 2007). Another report showed that incubating colon cancer cells with pathogenic E. coli (EIEC029:NM) reduced electrical resistance, inhibited cell-cell adhesion, disrupted the cytoskeleton and resulted in loss of expression of ZO-1 and occludin; however, probiotics reversed these changes (Johnson-Henry et al., 2008). In another report, probiotics markedly increased occludin expression by normalizing intestinal flora in an abdominal infection rat model (Qin et al., 2005). Thus, prebiotics might selectively promote the growth of beneficial intestinal flora and induce occludin expression to correct severe dysbacteriosis caused by high altitude exposure. The possible mechanisms of prebiotics are that they (1) Improve the microcirculation of the intestinal mucosa and prevent disruption of tight junction function due to energy depletion and (2) Regulate the receptors that are associated with the beneficial bacteria and activate occludin phosphorylation to maintain occludin expression and tight junction structure.
CONCLUSION
Overall, our study indicates that prebiotics can regulate the expression of occludin and the cytokines TNF-α and IL-10, which may help to maintain tight junction and intestinal barrier functions under the stress of high altitude exposure.
REFERENCES
- Abellan, R., A.F. Remacha, R. Ventura, M.P. Sarda, J. Segura and F.A. Rodriguez, 2005. Hematologic response to four weeks of intermittent hypobaric hypoxia in highly trained athletes. Haematologica, 90: 126-127.
PubMed - Ando-Akatsuka, Y., M. Saitou, T. Hirase, M. Kishi and A. Sakakibara et al., 1996. Interspecies diversity of the occludin sequence: CDNA cloning of human, mouse, dog and rat-kangaroo homologues. J. Cell Biol., 133: 43-47.
PubMedDirect Link - Baumgart, D.C. and A.U. Dignass, 2002. Intestinal barrier function. Curr. Opin. Clin. Nutr. Metab. Care, 5: 685-694.
PubMed - Blikslager, A.T., A.J. Moeser, J.L. Gookin, S.L. Jones and J. Odle, 2007. Restoration of barrier function in injured intestinal mucosa. Physiol. Rev., 87: 545-564.
CrossRefPubMedDirect Link - Chen, Y.H., Q. Lu, D.A. Goodenough and B. Jeansonne, 2002. Nonreceptor tyrosine kinase c-yes interacts with occludin during tight junction formation in canine kidney epithelial cells. Mol. Biol. Cell, 13: 1227-1237.
CrossRefDirect Link - Cummins, P.M., 2012. Occludin: One protein, many forms. Mol. Cell. Biol., 32: 242-250.
CrossRefPubMedDirect Link - Ioannidis, O., I. Varnalidis, G. Paraskevas and D. Botsios, 2011. Nutritional modulation of the inflammatory bowel response. Digestion, 84: 89-101.
CrossRefPubMedDirect Link - Johnson-Henry, K.C., K.A. Donato, G. Shen-Tu, M. Gordanpour and P.M. Sherman, 2008. Lactobacillus rhamnosus strain GG prevents enterohemorrhagic Escherichia coli O157: H7-induced changes in epithelial barrier function. Infect. Immunity, 76: 1340-1348.
CrossRef - Korzeniewski, K., 2008. Health problems in high mountain conditions. Polski Merkuriusz Lekarski, 25: 161-165.
PubMed - Qin, H.L., T.Y. Shen, Z.G. Gao, X.B. Fan, X.M. Hang, Y.Q. Jiang and H.Z. Zhang, 2005. Effect of lactobacillus on the gut microflora and barrier function of the rats with abdominal infection. World J. Gastroenterol., 11: 2591-2596.
PubMed - Schrezenmeir, J. and M. de Vrese, 2001. Probiotics, prebiotics and synbiotics-approaching a definition. Am. J. Clin. Nutr., 73: 361S-364S.
PubMedDirect Link