
Ha Na Ko
Department of Chemistry and Center for Cosmetic Sciences, Jeju National University, Ara-1, Jeju 690-756, Korea
Tae-Heon Oh
Department of Chemistry and Center for Cosmetic Sciences, Jeju National University, Ara-1, Jeju 690-756, Korea
Jong Seok Baik
Department of Chemistry and Center for Cosmetic Sciences, Jeju National University, Ara-1, Jeju 690-756, Korea
Chang-Gu Hyun
LINC Agency, Jeju National University, Ara-1, Jeju 690-756, Korea
Sang Suk Kim
Citrus Research Station, National Institute of Horticulture and Herbal Science, Seogwipo 697-943, Korea
Nam Ho Lee
Department of Chemistry and Center for Cosmetic Sciences, Jeju National University, Ara-1, Jeju 690-756, Korea
International Journal of Pharmacology
Year: 2013 | Volume: 9 | Issue: 2 | Page No.: 157-163







ABSTRACT
Development of natural products possessing anti-inflammatory activities has been the focus of research in our laboratory. In this study, the extract from Carpinus turczaninowii branches was studied and their anti-inflammatory constituents were identified. The aqueous ethanol extract was fractionated successively to afford n-hexane, ethyl acetate, n-butanol and aqueous layers. Upon anti-inflammatory tests for Nitric Oxide (NO) production, using RAW 264.7 cells, the ethyl acetate fraction exhibited potent inhibitory activities without causing cell toxicities. The ethyl acetate fraction was subjected to further purification to isolate carpinontriol A and carpinontriol B as active constituents. These diarylheptanoid compounds were isolated for the first time from this plant. The isolates reduced the lipopolysaccharide (LPS)-induced secretion of NO and IL-6 productions in a dose-dependent manner in RAW 264.7 cells which indicates their anti-inflammatory effects. Based on these results, it was suggested that C. turczaninowii extracts containing carpinontriols A and B could be considered potential anti-inflammatory agents for pharmaceuticals or cosmetics.
PDF Abstract XML References Citation
Received: March 17, 2013;
Accepted: May 04, 2013;
Published: June 05, 2013
How to cite this article
Ha Na Ko, Tae-Heon Oh, Jong Seok Baik, Chang-Gu Hyun, Sang Suk Kim and Nam Ho Lee, 2013. Anti-inflammatory Activities for the Extracts and Carpinontriols from Branches of Carpinus turczaninowii. International Journal of Pharmacology, 9: 157-163.
DOI: 10.3923/ijp.2013.157.163
URL: https://scialert.net/abstract/?doi=ijp.2013.157.163
DOI: 10.3923/ijp.2013.157.163
URL: https://scialert.net/abstract/?doi=ijp.2013.157.163
INTRODUCTION
Inflammation involves complex reactions of vascular tissues facing to various stimuli including pathogens, damaged cells or irritants. In the process of inflammation, activation of macrophages is the key step resulting in the production of inflammatory cytokines and mediators, such as Nitric Oxide (NO) and interleukin IL-6. Indeed, pro-inflammatory cytokines activate immune cells to up-regulate these inflammatory parameters. Therefore, regulation of these factors are useful targets in the development of new anti-inflammatory drugs, as well as in studies of their defense mechanisms with molecular levels (Zeilhofer and Brune, 2006; Jachak, 2007; Yoon et al., 2009).
Natural products from herbal plants have attracted attention as alternative therapies especially in Asian countries. A lot of traditional Chinese medicines have become the focus of studies aiming to elucidate their underlying efficacies as anti-cancer, anti-inflammation and anti-viral agents. Our research group also has investigated the potential role of natural products as a complementary medication in suppress of inflammation (Yoon et al., 2010a; Moon et al. 2011).
Carpinus belonging to the Butalaceae family is a genus of over 40 species. Most of this woody flora is distributed in the northern hemisphere including parts of China and Korea (Chang and Chang, 2010). The species C. turczaninowii (Betulaceae family), a deciduous tree endemic to Korea, has not been considered as a medicinal plant. However, our group has recently reported preliminary results of the high anti-inflammatory activity of C. turczaninowii extract (Yang et al., 2009). The leaves of C. tschonoskii have also been recently described to have anti-inflammatory properties in its extract (Kang et al. 2012). The extract of C. tschonoskii has exhibited cell-protection efficacies against Reactive Oxygen Species (ROS) damage by up-regulating catalase enzyme activity in V79-4 cells (Zhang et al., 2007). Previous chemical investigation of C. turczaninowii was only limited to the identification of flavonoids such as naringenin and quercetin glycosides (Jeon and Jang, 2000).
In this study, investigations of anti-inflammatory metabolites from this plant led to the isolation of two known diarylheptanoids, carpinontriol A and carpinontriol B, as the major constituents. These isolates were found to reduce the Lipopolysaccharide (LPs)-induced secretion of Nitric Oxide (NO) and to reduce IL-6 production in a dose-dependent manner in RAW 264.7 cells.
MATERIALS AND METHODS
Materials: Branches of Carpinus turczaninowii were collected in December 2009 from Halla Botanical Garden in Jeju Island, Korea. Voucher specimen (number 308) was deposited at the herbarium of Department of Chemistry, Jeju National University.
General: MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] was purchased from Sigma. All solvents used in this experiment were of analytical grade. 1H (400 MHZ) and 13C (100MHz) NMR spectra were recorded on a JEOL (JNM-ECX 400) instrument with chemical shift data reported in ppm relative to the solvent used. UV spectra were obtained in methanol using Biochrom Libra S22 (Cambridge Inc.) or SunriseTM (Tecan Co.) spectrophotometers. Concentrations of murine IL-6 in the culture supernatants were determined by ELISA (BD biosciences, USA). Silica gel (0.063-0.2 mm) from Merck was used for normal phased column chromatography. Merck silica gel 60 F254 coated on aluminum plates was used for Thin Layer Chromatography (TLC).
Isolation of carpinontriol A and carpinontriol B: The shade dried C. turczaninowii (707.0 g) was cut into small pieces and extracted with 70% aq. ethanol (14 L) at room temperature for 24 h. The gummy extract (89.0 g) was obtained after concentration of the filtered solution. Part of the ethanol extract (31.1 g) was suspended in water (2 L) and fractionated into n-hexane (0.37 g), ethyl acetate (4.1 g), n-butanol (7.9 g) and water (18.3 g) portions. The ethyl acetate (EtOAc) layer was subjected to Vacuum Liquid Chromatography (VLC) on silica gel using step-gradients (n-hexane/EtOAc to EtOAc/MeOH, 300 mL each) to provide 30 fractions (V1-V30). The combined fraction (295.9 mg) of V17 and V18, showing similar polarity patterns by TLC analysis, was purified by silica gel column chromatography (CC) with chloroform/EtOAc/MeOH (3:3:1) to afford compound 1 (carpinontriol A, 36.0 mg). The fraction V13 (91.9 mg) was also purified by silica gel CC with chloroform/EtOAc/MeOH (2:3:1) to give the compound 2 (carpinontriol B, 26.9 mg).
Cell culture: The murine macrophage cell line RAW 264.7 was purchased from the Korean Cell Line Bank (KCLB; Seoul, Korea). RAW 264.7 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; GIBCO Inc., NY) supplemented with 100 U mL-1 of penicillin, 100 μg mL-1 of streptomycin and 10% fetal bovine serum (FBS; GIBCO Inc., NY). The cells were incubated in an atmosphere of 5% CO2 at 37°C and were subcultured every 3 days.
MTT assay for cell viability: Cell viability was determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. RAW 264.7 cells were cultured in 96-well plates for 18 h, followed by treatment with LPS (1 μg mL-1) in the presence of various concentrations of samples (ethanol extracts, solvent fractions, carpinontriol A and carpinontriol B). After 24 h cell incubation, MTT was added to the medium and it was allowed to stand for 4 h. After removing the supernatant, the formazan crystals were dissolved in DMSO. Absorbance was measured at 540 nm. The percentage of cells showing cytotoxicity was determined relative to the control group.
Measurement of nitric oxide (NO) concentration: Nitric oxide production was assayed by measuring the nitrite in the supernatants of cultured RAW 264.7 cells. The cells were seeded at densities of 2.5x105 cells/mL in 24 well culture plates and were cultured for 18 h. The cells were stimulated with LPS (1 μg mL-1) and various concentrations of samples (ethanol extracts, solvent fractions, carpinontriol A and carpinontriol B) for 24 h and were then centrifuged. The supernatant was mixed with an equal volume of Griess reagent (1% sulfanilamide, 0.1% naphthylethylenediamine dihydrochloride and 2.5% phosphoric acid) and was incubated at room temperature for 10 min. The concentrations of nitrite were then determined at an optical density of 540 nm. Nitrite concentration was determined by comparing the optical density with the standard curve obtained with sodium nitrite. All experiments were performed in triplicate.
Detection of IL-6 in supernatant: Sandwich ELISA was used to determine the inhibitory effects of 50-500 μM carpinontriol A and carpinontriol B on the production of cytokine interleukin-6. Macrophage RAW 264.7 cells were stimulated for 24 h before the supernatant was harvested and were then assayed according to the manufacturer’s protocol for the relevant ELISA kit. Results from three independent experiments were used for statistical analysis.
Statistical analysis: The Student’s t-test and one-way ANOVA were used to determine statistically significant differences between the values for the various experimental and control groups. Data are expressed as Means±Standard errors (SEM) and the results represent at least three independent experiments. Values are the Mean±SEM of triplicate experiments. *p<0.05; **p<0.01.
RESULTS AND DISCUSSION
It is well-established that RAW 264.7 cells, a mouse macrophage cell line, induce the expression of the enzyme iNOS which produces large amounts of nitric oxide when activated with LPS. Therefore, the modulation of macrophage-mediated inflammatory responses is recognized as a promising new therapeutic approach against inflammatory diseases.
To investigate the effect of C. turczaninowii extracts and fractions on NO production, the accumulation of nitrite, a stable oxidized product of NO, was measured in culture media. The production of nitric oxide was compared for RAW 264.7 macrophages in the presence or absence of C. turczaninowii extracts for 24 h. As shown in Fig. 1, the ethanol extract of C. turczaninowii (100 μg mL-1) inhibited LPS-induced NO production in RAW 264.7 cells by 36.9%. In order to determine the active constituents responsible for the anti-inflammatory activities, the solvent fractions (n-hexane, EtOAc, BuOH and water layers) prepared from the extract were screened. As shown in Fig. 1, n-hexane and EtOAc fractions exhibited potent NO inhibition activities. However, the n-hexane layer exhibited considerable cell toxicity as determined by MTT assay (Fig. 1). On the other hand, the EtOAc fraction showed little loss of viable cells at a concentration of 100 μg mL-1, indicating that the observed NO inhibition is not merely as a result of cell-destruction effects. In Fig. 2, the NO inhibition and cytotoxicity data with varying concentrations 25-200 μg mL-1 were presented for the ethanol extract (A) as well as EtOAc (B) and BuOH (C) fractions. It was shown that they all decreased NO generation in a dose-dependent manner (Fig. 2, a-c). The most strong inhibition activity was observed in EtOAc fraction with IC50 of 57.5 μg mL-1. They did not display any cytotoxicities at concentrations up to 100 μg mL-1. Slight cell toxicity was appeared at the high concentration (200 μg mL-1) for EtOAC fraction.
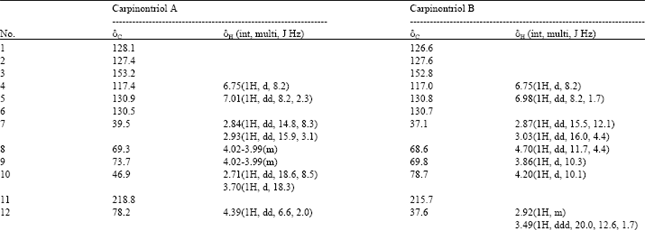
As the EtOAc fraction showed strong inhibition of NO synthesis without causing cell toxicity under 200 μg mL-1, it was chosen for further fractionation to determine the active constituents. Through chromatographic purification procedures, the compounds 1 and 2 were isolated. Their chemical structures were verified based on interpretation of spectroscopic information, including 1D and 2D NMR data. The compounds 1 and 2 showed 19 carbon peaks in 13C NMR spectra which are characteristic indications for the presence of a diarylheptanoid skeleton (Table 1). Twelve sp2 carbons peaks corresponding to two aromatics also support the suggested structures. The signals at δ 218.8 and 215.7 in 13C NMR spectrum indicate the presence of carbonyl group for the compounds 1 and 2, respectively. The compound 1 showed three oxygen-bearing aliphatic carbons (δ 46.9, 69.3, 78.2) as indicated in HMQC data. Likewise, the similar carbons (δ 68.6, 69.8, 78.7) attached to oxygen were also observed in the compound 2. Detailed investigation of spectroscopic data led to the identification of the compounds 1 and 2 as carpinontriol A and B, respectively. The suggested structures were finally confirmed by comparison of observed data to literature values (Lee et al., 2002). The carpinontriol A and B are known compounds and have diarylheptanoid skeleton bearing one carbonyl and five hydroxyl functional groups (Fig. 3). As shown in Fig. 3, they have almost the same chemical structures, except for the position of one hydroxyl group either in C-10 (compound 1) or C-12 (compound 2). Interestingly, compounds 1 and 2 were isolated for the first time from C. turczaninowii.
To assess the effect of the isolates on NO production in RAW 264.7 cells, various concentrations (50, 100, 300 and 500 μM) of the compounds 1 and 2 were applied to the LPS-treated macrophage. The levels of nitric oxide were measured by Griess reaction.
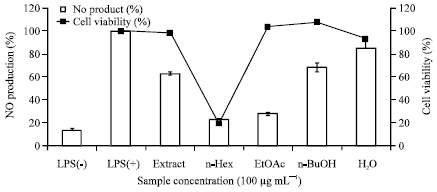 | |
| Fig. 1: | Effect of solvent fractions from C. turczaninowii on NO production in LPS-stimulated RAW 264.7 cells. The cells were stimulated with 1 μg mL-1 of LPS only, or with LPS plus C. turczaninowii extract (100 μg mL-1) for 24 h. Nitric oxide production was determined by the Griess reagent method. Cell viability was determined after 24 h culture of cells stimulated with LPS (1 μg mL-1) in the presence of C. turczaninowii. The data represent the Mean±SD of triplicate experiments |
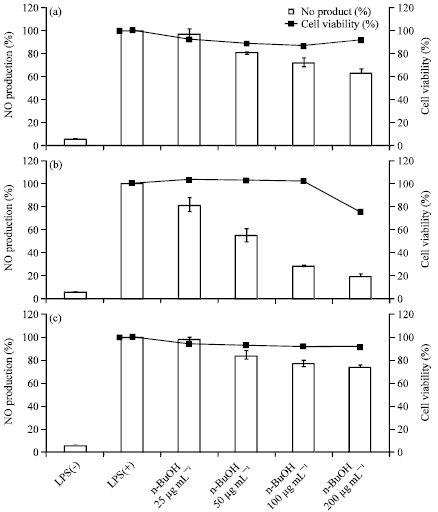 | |
| Fig. 2(a-c): | Effect of EtOH extract (a), EtOAc layer (b) and n-BuOH layer (c) on NO production in LPS-stimulated RAW 264.7 cells. The cells were stimulated with 1 μg mL-1 of LPS only, or with LPS plus EtOH extract, EtOAc layer or n-BuOH layer (100 μg mL-1) for 24 h. Nitric oxide production was determined by the Griess reagent method. The data represent the Mean±SD of triplicate experiments |
| Table 1: | 1H and 13C NMR data of carpinontriol A and carpinontriol B (100 and 400 MHZ, CD3OD) |
 | |
 | |
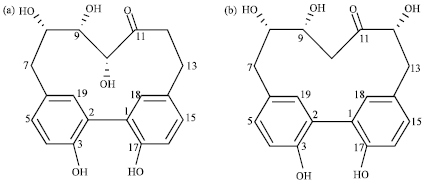 | |
| Fig. 3: | Chemical structures of isolated compounds 1 (carpinontriol A) and 2 (carpinontriol B) |
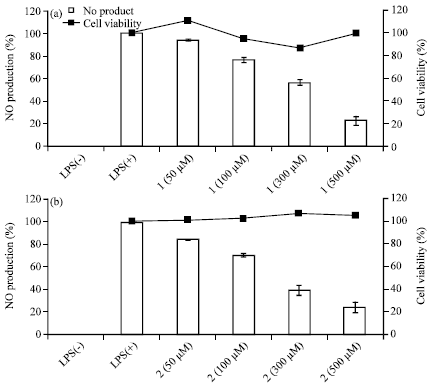 | |
| Fig. 4(a-b): | Effect of carpinontriol A (A) and carpinontriol B (B) on nitric oxide production in LPS-stimulated RAW 264.7 cells. The cells were stimulated with 1 μg mL-1 of LPS only, or with LPS plus various concentrations (50-500 μM) of carpinontriol A and carpinontriol B for 24 h. Nitric oxide production was determined using the Griess reagent method. Cell viability was determined from the 24 h culture of cells stimulated with LPS (1 μg mL-1) in the presence of carpinontriol A and carpinontriol B. Values are presented as the Mean±SEM of triplicate experiments |
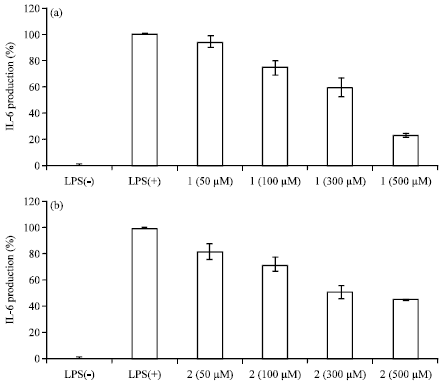 | |
| Fig. 5(a-b): | Effects of carpinontriol A and carpinontriol B on IL-6 production in LPS-stimulated RAW 264.7 cells. The cells were stimulated with 1 μg mL-1 LPS only or with LPS plus various concentrations (50-500 μM) of carpinontriol A and carpinontriol B for 24 h. IL-6 produced and released into the culture medium was assayed using the ELISA method |
As appeared in Fig. 4, the carpinontriol A (1) and B (2) highly reduced the NO synthesis with IC50 396.0 and 199.4 μM, respectively. Nitric oxide inhibition was dose-dependent for the both compounds. Upon the examination of the cytotoxicities of carpinontriols A and B by MTT assay, they did not damage the cell viability at the employed concentrations (50, 100, 300 and 500 μM). Thus, it is clear that the observed nitric oxide inhibition activities are not caused by the cell toxicities.
Interleukin 6 (IL-6) is one of the earliest and most important pro-inflammatory cytokines produced in response to inflammatory stimuli. As our continuing research searching for anti-inflammatory agents, a number of plant extracts and natural products were discovered to suppress the secretion of IL-6 in LPS-treated macrophages in vitro (Yang et al., 2010; Yoon et al., 2010b, c). As an identical program, experiments were conducted to evaluate the compounds 1 and 2 for the inhibitory effects on IL-6 secretion in LPS-stimulated RAW 264.7 cells. The macrophage cells were pretreated for 1 h with isolates 1 and 2 and IL-6 levels were measured 4 h after LPS stimulation. As shown in Fig. 5, it was clear that carpinontriol A and B reduced the LPS-induced secretion of IL-6 in a dose-dependent manner. Though the degree of inhibition is very similar, but slightly higher potency was observed for the compound 1 at higher concentration (500 μM).
CONCLUSION
The present study demonstrates that the ethanol extracts of C. turczaninowii possess strong inhibition properties towards nitric oxide production in LPs-stimulated macrophages without causing cell toxicity. The isolated compounds, carpinontriol A and carpinontriol B, are also potent inhibitors of NO and IL-6 production in RAW 264.7 cells. The exact mechanisms operating the inhibition activity for compounds 1 and 2 are not fully known at this point. It could be inferred that the inhibition of NO production seems to be due to the down-regulation of iNOS protein and mRNA expression which might be associated with the attenuation of IL-6 formation. Ultimately, it is suggested that that C. turczaninowii extracts, containing carpinontriol A and carpinontriol B, appear to be potential anti-inflammatory candidates for use in pharmaceuticals and cosmetics.
ACKNOWLEDGMENTS
This study was supported by a program of Business for Academic-industrial Cooperative establishment funded by Korea Small and Medium Business Administration in a year of 2011 (2011-00045876).
REFERENCES
- Chang, C.S. and K.S. Chang, 2010. Typification of Corylopsis coreana (Hamamelidaceae) and Carpinus laxiflora var. longispica (Betulaceae). J. Jpn. Bot., 85: 270-276.
Direct Link - Jachak, S.M., 2007. PGE synthase inhibitors as an alternative to COX-2 inhibitors. Curr. Opin. Investi. Drugs, 8: 411-415.
PubMed - Jeon, J.I. and J.S. Jang, 2000. Foliar flavonoids of genus Carpinus in eastern Asia primarily based on native taxa to Korea. Kor. J. Plant Tax., 30: 139-153.
Direct Link - Kang, S.H., J.E. Koo, H.J. Hong, V.B. Mathema and Y.S. Koh, 2012. Anti-inflammatory activity of Carpinus tschonoskii leaves extract in R848-stimulated bone marrow-derived macrophages and dendritic cells. J. Bacteriol Virol., 42: 77-82.
CrossRef - Lee, J.S., H.J. Kim, H.K. Parkand Y.S. Lee, 2002. New diarylheptanoids from the stems of Carpinus cordata. J. Nat. Prod., 65: 1367-1370.
PubMed - Moon, J.Y., E.J. Yang, S.S. Kim, J.Y. Kang, G.O. Kim, N.H. Lee and C.G. Hyun, 2011. Sasa quelpaertensis phenylpropanoid derivative suppresses lipopolysaccharide-induced nitric oxide synthase and cyclo-oxygenase-2 expressions in RAW 264.7 cells. Yakugaku Zasshi, 131: 961-967.
PubMed - Yang, E.J., E.Y. Yim, G. Song, G.O. Kim and C.G. Hyun, 2009. Inhibition of nitric oxide production in lipopolysaccharide-activated RAW 264.7 macrophages by Jeju plant extracts. Toxicology, 2: 245-249.
PubMed - Yang, E.J., J.Y. Moon, M.J. Kim, D.S. Kim and C.S. Kim et al., 2010. Inhibitory effect of Jeju endemic seaweeds on the production of pro-inflammatory mediators in mouse macrophage cell line RAW 264.7. J. Zhejiang. Univ. Sci. B., 11: 315-322.
PubMed - Yoon, W.J., Y.M. Ham, S.S. Kim, B.S. Yoo and J.Y. Moon et al., 2009. Suppression of pro-inflammatory cytokines, iNOS and COX-2 expression by brown algae Sargassum micracanthum in RAW 264.7 macrophages. EurAsia J. BioSci., 3: 130-143.
CrossRef - Yoon, W.J., N.H. Lee and C.G. Hyun, 2010. Limonene suppresses lipopolysaccharide-induced production of nitric oxide, prostaglandin E2 and pro-inflammatory cytokines in RAW 264.7 macrophages. J. Oleo. Sci., 59: 415-421.
PubMedDirect Link - Yoon, W.J., J.Y. Moon, J.Y. Kang, G.O. Kim, N.H. Lee and C.G. Hyun, 2010. Neolitsea sericea essential oil attenuates LPS-induced inflammation in RAW 264.7 macrophages by suppressing NF-kappaB and MAPK activation. Nat. Prod. Commun., 5: 1311-1316.
PubMed - Yoon, W.J., J.Y. Moon, G. Song Y.K. Lee and M.S. Han et al., 2010. Artemisia fukudo essential oil attenuates LPS-induced inflammation by suppressing NF-κB and MAPK activation in RAW 264.7 macrophages. Food Chem. Toxicol., 48: 1222-1229.
CrossRef - Zhang, R., K.A. Kang, M.J. Piao, J.W. Park and T.K. Shin et al., 2007. Cytoprotective activity of Carpinus tschonosk against H2O2 induced oxidative stress. Nat. Prod. Sci., 13: 118-122.
Direct Link - Zeilhofer, H.U. and K. Brune, 2006. Analgesic strategies beyond the inhibition of cyclooxygenases. Trends Pharmacol. Sci., 27: 467-474.
CrossRefPubMedDirect Link