
Seung-Heon Shin
Department of Otorhinolaryngology, School of Medicine, Catholic University of Daegu, Daegu, South Korea
Mi-Kyung Ye
Department of Otorhinolaryngology, School of Medicine, Catholic University of Daegu, Daegu, South Korea
Jung-Kyu Kim
Department of Otorhinolaryngology, School of Medicine, Catholic University of Daegu, Daegu, South Korea
Kwan-Kyu Park
Department of Pathology, School of Medicine, Catholic University of Daegu, Daegu, South Korea
International Journal of Pharmacology
Year: 2013 | Volume: 9 | Issue: 2 | Page No.: 143-149







ABSTRACT
Bee Venom (BV) has been used as treatment against a variety of inflammatory diseases. Studies have demonstrated anti-inflammatory effect of BV. This study aimed to clarify the potential clinical efficacy of BV concerning the anti-inflammatory effect on bronchial epithelial cell. BEAS-2B cells were exposed to Alternaria alternata and Aspergillus nigra with or without BV. IL-6, IL-8 and granulocyte macrophage colony-stimulating factor were measured to determine the activation of epithelial cells. Nuclear factor kappa B (NF-κB) and activator protein 1 expression and activity were determined with Western blot analysis and ELISA. Cytotoxicity of BV was measured using a CellTiter-96® aqueous cell proliferation assay kit. Cell survival was significantly decreased concentrations at 10 μg mL-1. Fungi induced IL-6 and IL-8 production was effectively inhibited by BV. BV suppressed fungi induced NF-κB expression and Alternaria induced NF-κB activation. BV seems relatively safe and is of potential value for the treatment of airway inflammatory diseases.
PDF Abstract XML References Citation
Received: February 28, 2013;
Accepted: April 23, 2013;
Published: June 05, 2013
How to cite this article
Seung-Heon Shin, Mi-Kyung Ye, Jung-Kyu Kim and Kwan-Kyu Park, 2013. Bee Venom Reduces Fungi Induced Bronchial Epithelial Cells Activation Through Down Regulation of NF-κB. International Journal of Pharmacology, 9: 143-149.
DOI: 10.3923/ijp.2013.143.149
URL: https://scialert.net/abstract/?doi=ijp.2013.143.149
DOI: 10.3923/ijp.2013.143.149
URL: https://scialert.net/abstract/?doi=ijp.2013.143.149
INTRODUCTION
Bee Venom (BV) has been used to treat chronic inflammatory diseases and malignant tumors for long time in Oriental medicine (Hider, 1988). BV contains enzymes, biologically active amines, peptides and nonpeptide components with various pharmacological characteristics. Studies in animal models has demonstrated the capability of BV to modify the immune function in the body with anti-inflammatory and anti-bacterial activities in the absence of side effects (Han et al., 2007). Anti-inflammatory activity is due to the ability of BV to inhibit the expressions of cycloxygenase-2 and phospholipase A2. BV also inhibits the release of inflammatory mediators similar to nonsteroidal anti-inflammatory drugs.
Fungal spores are continuously inhaled and deposit in normal and patient’s airway mucosa without harmful effect. But fungi that are commonly involved in asthmatic reaction with atopic response belong to Alternaria, Cladosporium, Aspergillus, Penicillium and Candida (Knutsen et al., 2012). Fungal elements like proteins and enzymes induce immunologic and inflammatory responses. Proteases from fungi activate epithelial cells through surface receptors, resulting in generation of cytokines and facilitating antigen access either by proteolytic attack or cell desquamation (Kauffman et al., 1995; Shin and Lee, 2010).
The respiratory epithelium is the first mucosal defense organ exposed to the environmental stimulants associated with airway disease. Respiratory epithelial cells provide mechanical barrier to prevent infection and produce chemokines, cytokines and antimicrobial components to eliminate pathogenic microorganisms. These cells also play an active role in inflammation through the production of a variety of proinflammatory cytokines and chemokines for the recruitment of inflammatory cells (Matsuwaki et al., 2012).
Humans are constantly exposed to various fungi, but only a few, especially Alternaria and Aspergillus, are associated with respiratory disease. The proteolytic enzymes released by fungi induce the production of chemoattractive and proinflammatory cytokines by respiratory epithelial cells (Shin and Lee, 2010; Matsuwaki et al., 2012). Until now, no studies have been performed to investigate the anti-inflammatory effect of BV on fungi-activated bronchial epithelial cells. The aim of this study was to evaluate the effect of BV on the production of chemical mediators from activated bronchial epithelial cells and their anti-inflammatory mechanisms.
MATERIALS AND METHODS
Bronchial epithelial cell culture and activation with fungi: The human bronchial epithelial cell line BEAS-2B, which was derived from human bronchial epithelium transformed by adenovirus 12-SV40, was purchased from American Type Culture Collection (Rockville, MD, USA). Cells were cultured with DMEM/F12 medium supplemented with heat-inactivated 10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA), 100 U mL-1 penicillin and 100 μg mL-1 streptomycin at 37°C and 5% CO2. Cells were seeded at 5x104 cells/well in a 24 well tissue culture plate and grown until 80% confluence. At this stage, the BEAS-2B cells were stimulated for up to 24 h with endotoxin free Alternaria alternata (50 μg mL-1) and Aspergillus nigra (25 μg mL-1) (Greer Lab, Lenoir, NC, USA). In our previous experiments with nasal polyp epithelial cells, maximal cytokine production was found at 50 μg mL-1 of Alternaria and 25 μg mL-1 of Aspergillus (Shin and Lee, 2010). The same concentrations of these allergens were used currently. Supernatants and cells were harvested and stored at -70°C until they were assayed. Production of interleukin (IL)-6, IL-8 and granulocyte-macrophage colony stimulating factor (GM-CSF) was quantified using commercially available enzyme-linked immunosorbant assay (ELISA) kits (R and D Systems, Minneapolis, MN, USA).
Preparation and cytotoxicity assay of BV: Pure honeybee (Apis mellifera) venom was obtained from the National Institute of Agricultural Science and Technology, Suwon, Korea. BV was collected using a specialized collector without damaging honeybees by an established electric shock method. BV was dissolved in distilled water and centrifuged at 12,000xg for 10 min to remove insoluble materials. The BV was lyophilized by freeze drying and stored (Han et al., 2007).
BEAS-2B cells were incubated at 37°C, 5% CO2 in 96 well plates in the presence of 0, 0.03, 0.15, 0.5, 1, 5 and 10 μg mL-1 BV. Cytotoxicity was measured using a CellTiter-96® aqueous cell proliferation assay kit (Promega, Madison WI, USA). For this assay, the tetrazolium compound and Owen’s reagent were added to each well and incubated for 4 h at 37°C in a humidified 5% CO2 chamber. Color intensities were assessed with a microplate reader at 490 nm.
Expression and activation of nuclear factor kappa B (NF-κB) and activator protein-1 (AP-1): To perform Western blot analysis for NF-κB and AP-1, NE-PER nuclear extraction reagents (Pierce Chemical, Rockford, IL, USA) was used to isolate proteins. The protein was quantified using a BCATM protein assay reagent kit (Pierce Chemical). The resulting protein was separated on a NuPAGE 4-12% Bis-Tris gel (Invitrogen) and transferred onto a nitrocellulose membrane (Bio-Rad Laboratories, Hercules, CA, USA). The membrane was blocked with membrane blocking solution (Invitrogen) and probed with anti-human NF-κB and c-Jun antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 4°C overnight. After washing, the membranes were incubated with secondary goat anti-rabbit antibody coupled with horseradish peroxidase (HRP) for 1 h at room temperature and developed with SuperSignal West Pico Chemiluminescent substrate (Pierce Chemical). The band intensities were expressed as a percentage of treated versus untreated cells.
Nuclear NF-κB and AP-1 activities were quantified using an ELISA-based TransAM kit (Active Motif, Carlsbad, CA, USA) according to the manufacturer’s protocol. In brief, the nuclear protein extracts were added to wells of a 96-well plate that was immobilized with oligonucleotide containing NF-κB and AP-1. NF-κB and AP-1 contained in the nuclear extracts bound specifically to the oligonucleotide during incubation for 1 h at room temperature. NF-κB antibody (1:1000 dilution) and AP-1 antibody (1:500 dilution) were then added to each well for 1 h followed by HRP-conjugated secondary antibody for 1 h. After adding 100 μL of detection reagent for up to 15 minutes, the colorimetric reaction was stopped. NF-κB and AP-1 activities were determined by reading the absorbance on a spectrophotometer at 450 nm with a reference wavelength of 655 nm.
Statistical analysis: All experiments were performed at least five times and produced comparable results. Results are presented as the Mean±SD. Statistical significance was determined using one-way Analysis of Variance (ANOVA) followed by a Dunnett’s test for multiple comparisons to determine the overall effect of BV pretreatment on BEAS-2B cells. One factor repeated measure analysis was performed to determine the cytotoxic effect and cytokine production effect of BV (SPSS ver. 14.0; SPSS Inc., Chicago, IL, USA). A p-value of 0.05 or less was considered significant.
RESULTS
Cytotoxic effect of BV: In order to determine the optimal dose and treatment time of cytotoxic effect of BV on BEAS-2B, an established viability assay utilizing MTT was carried out. The cells were treated with BV at concentration of 0.03, 0.15, 0.5, 5 and 10 μg mL-1 for 24, 48 and 72 h.
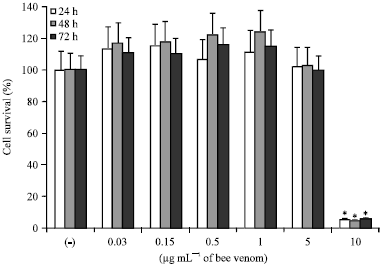 | |
| Fig. 1: | Cytotoxic effect of bee venom (BV) on bronchial epithelial cells (BEAS-2B) at various concentrations and times with CellTiter-96® aqueous cell proliferation assay. Cell survival was significantly decreased at BV concentrations at 10 μg mL-1, *p<0.05 compared to negative control, n = 5 |
The viability of cells treated with BV at a concentration of 10 μg mL-1 were 5.8±2.3% for 24 h, 4.43±1.4% for 48 h and 5.8±2.2% for 72 h. Although the concentration of BV influenced the survival of epithelial cells, BV exposure time did not significantly influence the cell survival (Fig. 1).
Effects of BV on the stimulants induced cytokine production: When the BEAS-2B cells were stimulated with Alternaria (IL-6; 413.7±117.1 pg mL-1, IL-8; 2736.2±148.8 pg mL-1 and GM-CSF; 383.3±149.2 pg mL-1) and Aspergillus (IL-6; 506.4±142.7 pg mL-1, IL-8; 2947.1±422.58 pg mL-1 and GM-CSF; 452.6±133.3 pg mL-1), cytokine production was significantly increased. IL-6 and IL-8 production were significantly inhibited by BV. However, GM-CSF production was not inhibited (Fig. 2). For example, 24.1-36.7% of Alternaria induced IL-6 production was inhibited by BV concentrations of 1, 0.5, 0.1 and 0.01 μg mL-1 and 22.1-24.1% of Alternaria induced IL-8 production using BV concentrations of 0.1 and 0.01 μg mL-1.
Effects of BV on the fungi induced expression and activation of NF-κB and AP-1: When BEAS-2B cells were stimulated with Alternaria and Aspergillus, nuclear NF-κB expression was significantly increased for 24 h. The increased NF-κB expression was significantly inhibited by 0.1 and 0.01 μg mL-1 BV. Alternaria induced NF-κB expression was inhibited at 3 and 24 h after stimulation and Aspergillus induced NF-κB expression was inhibited at 3, 6, 12 and 24 h. When the BEAS-2B cells were treated with fungi, AP-1 expression was not significantly increased (Fig. 3).
After stimulation with fungi for 24 h, NF-κB and AP-1 activations were evaluated in an ELISA-based assay kit using positive control (Jukat nuclear extract for NF-κB and K-562 nuclear extract for AP-1). Alternaria induced nuclear NF-κB activity was significantly inhibited when BEAS-2B cells were treated with BV. However nuclear AP-1 activity was not influenced by fungi and BV (Fig. 4).
DISCUSSION
Asthma is the lower airway inflammatory disease with airway hypersensitivity and airflow limitation. Their recent incidence, prevalence and severity have continuously increased. Although the etiology of asthma is complex and multifactorial, fungi play an important role in the pathogenesis of asthma. Severe asthma and acute exacerbation of asthma have been associated fungal sensitivity or increased concentration of fungi (Zureik et al., 2002). Airborne fungi and their elements induce immune and inflammatory response from respiratory epithelial cells. The interaction of fungi with respiratory epithelial cell receptors, by protease-activated receptor or toll-like receptors, leads to the production of inflammatory chemical mediators, induction of respiratory burst and inflammatory cell recruitment.
In this study, Alternaria and Aspergillus were used to activate the bronchial epithelial cells, because these organisms are common pathogens found in respiratory secretions and in respiratory tract diseases. Alternaria and Aspergillus extracts have a high protease dependent activation and are highly potent in inducing cytokine production from upper and lower airway epithelial cells (Kauffman et al., 1995; Shin and Lee, 2010).
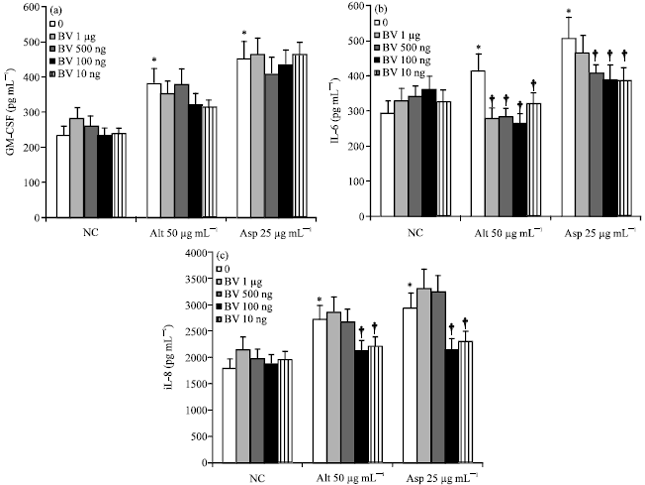 | |
| Fig. 2(a-c): | Influence of bee venom on fungi induced cytokine production by bronchial epithelial cells (BEAS-2B). When BEAS-2B cells were treated with various concentrations (0.01, 0.1, 0.5 and 1 μg mL-1) of BV, Alternaria 50 μg mL-1 (Alt 50) and Aspergillus 25 μg mL-1 (Asp 25) induced IL-6 and IL-8 production was significantly inhibited. NC: negative control, Alt: Alternaria, Asp: Aspergillus, BV: bee venom, *p<0.05 compared with non-stimulated group †<0.05 compared to Alternaria or Aspergillus stimulated groups, n = 7 |
Alternaria and Aspergillus induced IL-6, IL-8 and GM-CSF production from BEAS-2B cells. These proinflammatory cytokine influence the pathogenesis and symptom development of asthma. The expressions of inflammatory mediators are regulated by the activation of protein kinase C, mitogen-activated kinase, NF-κB and AP-1 (Fujioka et al., 2004). When BEAS-2B cells were stimulated with fungi, cytokine productions were significantly increased with up-regulation of intranuclear NF-κB. NF-κB regulates the expression of cytokines, growth factors and effector enzymes in response to ligation of many receptors involved in immune responses. However, presently, AP-1 expression was not influenced by exposure to the fungi.
The data concerning transcription factor expression and activation in respiratory epithelial cells initially prompted the consideration, that NF-κB might be the most important factor associated with cytokine production. Alternaria induced NF-κB expression was stronger than Aspergillus and continuously increased for 24 h. NF-κB activity was significantly increased by Alternaria but not by Aspergillus. Alternaria might more strongly induce the production of chemical mediators and might be associated with the aggravation of respiratory symptoms.
BV has anti-inflammatory, anti-nociceptive and anti-cancer activities (Jung et al., 2007; Han et al., 2007; Lee et al., 2001; Shin et al., 2013). The anti-inflammatory effect is particularly well established and various mechanisms have been suggested. Peptides of BV have anti-inflammatory effects due to their ability to inhibit prostaglandin synthesis system or stabilize mast cells. BV also inhibits the activity of cyclooxygenase (COX-2) and expression of COX-2 mRNA in a dose-dependent manner, without having any cytotoxic effects (Nam et al., 2003).
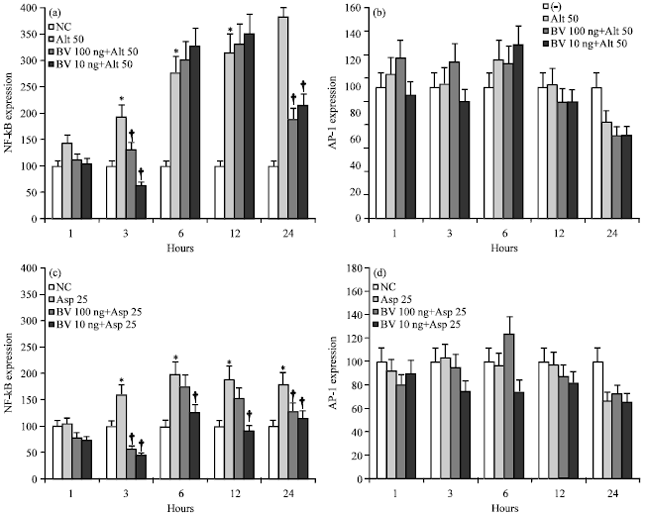 | |
| Fig. 3(a-d): | Expression of NF-κB and AP-1 in bronchial epithelial cells (BEAS-2B) stimulated by Alternaria 50 μg mL-1 (Alt 50) and Aspergillus 25 μg mL-1 (Asp 25). NF-κB expression was significantly increased by fungi at various time intervals and Bee Venom (BV) significantly inhibited NF-κB expression. Although fungi did not influence the AP-1 expression, BV tended to inhibit AP-1 expression. NC: Negative control, *p<0.05 compared with non-stimulated group, †<0.05 compared to Alternaria or Aspergillus stimulated group, n = 5 |
When BEAS-2B cells were pre-treated with various concentration of BV, the fungi induced production of IL-6 and IL-8 were significantly inhibited. The anti-inflammatory effect was stronger at lower concentrations of BV (10 and 100 ng mL-1) than at higher concentrations (1 μg mL-1 and 500 ng mL-1). These results were similar to previous experiment with nasal epithelial cells (Shin et al., 2013). Lower than 100 ng mL-1 of BV had anti-inflammatory property on upper and lower airway inflammation. At higher concentrations of BV, enzymatic components that enhance inflammatory responses might more actively or strongly influence the immune response of bronchial epithelial cells. For the clinical use of BV, the optimal concentration that contains maximal anti-inflammatory effects without harmful or cytotoxic effects should be determined.
NF-κB and AP-1 are key transcription factors associated with induction and regulation of chemical mediators involved in inflammation. Although NF-κB and AP-1 transcription factors are regulated by different mechanisms, the signaling pathways leading to activation of NF-κB and AP-1 overlap, which suggests that they work cooperatively (Fujioka et al., 2004). At lower concentrations, BV inhibited fungi induced IL-6 and IL-8 production and NF-κB expression and activity. These inhibitory effects are consistent with the decrease in the release of IκB detected in the cytosol through the suppression of IκB phosphorylation and the decrease in the translocation of the p50 subunit of NF-κB (Park et al., 2007). This means that BV inhibits the DNA binding activity of NF-κB by inhibiting IκB phosphorylation, thereby preventing p50 translocation, resulting in a decrease in the expression of the inflammatory gene.
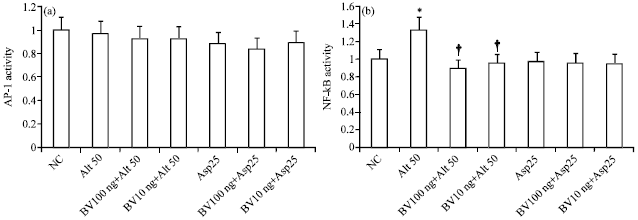 | |
| Fig. 4(a-b): | Intensity of NF-κB and AP-1 activity in bronchial epithelial cells (BEAS-2B) stimulated by Alternaria 50 μg mL-1 (Alt 50) and Aspergillus 25 μg mL-1 (Asp 25). Bee venom (BV) significantly inhibited Alternaria induced NF-κB activation. AP-1 activity was not significantly changed with fungi and BV treatment. NC: Negative control, *p<0.05 compared with non-stimulated group, †<0.05 compared to Alternaria stimulated group, n = 5 |
However, GM-CSF production was not inhibited with BV. Thus, NF-κB may be involved in the IL-6 and IL-8 release, with another transcription factor, such as mitogen-activated protein kinase, being associated with the production of GM-CSF. These results were similar to previous experiment with nasal epithelial cells (Shin et al., 2013). Lower concentrations of BV inhibited intranuclear NF-κB expression and activity in upper and lower airway inflammation. Fungi did not influence the AP-1 expression; however, BV tends to suppress the AP-1 expression. Lee et al. (2009) suggested that BV abolishes the expression and translation of PKC-α, phosphorylation of MEK/ERK and expression and DNA binding activity of AP-1/NF-κB (Lee et al., 2009).
CONCLUSION
The present findings elucidate the anti-inflammatory mechanism of activated bronchial epithelial cells with BV. Alternaria and Aspergillus can induce cytokine production from BEAS-2B cells and enhance the expression and activity of NF-κB. BV suppresses the production of cytokines, especially IL-6 and IL-8, with inhibition of the expression and activity of NF-κB. Although further study is needed for determination the more detail anti-inflammatory mechanism of BV, an optimal effect convention of BV and the effects in-vivo, BV contributes to the development of a new therapeutic strategy for airway inflammatory diseases.
ACKNOWLEDGMENT
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2010-0023163).
REFERENCES
- Fujioka, S., J. Niu, C. Schmidt, G.M. Sclabas and B. Peng et al., 2004. NF-κB and AP-1 connection: Mechanism of NF-κB-dependent regulation of AP-1 activity. Mol. Cell Biol., 24: 7806-7819.
CrossRefDirect Link - Han, S.M., K.G. Lee, J.H. Yeo, H.Y. Kweon and S.O. Woo et al., 2007. Effect of honey bee venom on microglial cells nitric oxide and tumor necrosis factor-α production stimulated by LPS. J. Ethnopharmacol., 111: 176-181.
CrossRef - Hider, R.C., 1988. Honeybee venom: A rich source of pharmacologically active peptides. Endeavour, 12: 60-65.
CrossRef - Jung, H.J., K.K. Park, J.H. Park, K.H. Kim and C.H. Cho et al., 2007. Effects of whole bee venom on expression of TNF-α, nitric oxide, TGF-β1 in collagen-induced arthritis model. Korean J. Apicult., 22: 181-188.
Direct Link - Knutsen, A.P., R.K. Bush, J.G. Demain, D.W. Denning and A. Dixit et al., 2012. Fungi and allergic lower respiratory tract diseases. J. Allergy Clin. Immunol., 129: 280-291.
CrossRef - Lee, J.H., Y.B. Kwon, H.J. Han, W.C. Mar and H.J. Lee et al., 2001. Bee venom pretreatment has both an antinociceptive and anti-inflammatory effect on carrageenan-induced inflammation. J. Vet. Med. Sci., 63: 251-259.
CrossRefPubMedDirect Link - Lee, K.G., H.J. Cho, Y.S. Bae, K.K. Park and J.Y. Choe et al., 2009. Bee venom suppresses LPS-mediated NO/iNOS induction through inhibition of PKC-α expression. J. Enthnopharmacol., 123: 15-21.
CrossRef - Matsuwaki, Y., K. Wada, T. White, H. Moriyama and H. Kita, 2012. Alternaria fungus induces the production of GM-CSF, interleukin-6 and interleukin-8 and calcium signaling in human airway epithelium through protease-activated receptor 2. Int. Arch. Allergy Immunol., 158: 19-29.
CrossRefPubMedDirect Link - Nam, K.W., K.H. Je, J.H. Lee, H.J. Han, H.J. Lee, S.K. Kang and W. Mar, 2003. Inhibition of COX-2 activity and proinflammatory cytokines (TNF-α and IL-1β) production by water-soluble sub-fractionated parts from bee (Apis mellifera) venom. Arch. Pharm. Res., 26: 383-388.
CrossRefDirect Link - Park, H.J., D.J. Son, C.W. Lee, M.S. Choi and U.S. Lee et al., 2007. Melittin inhibits inflammatory target gene expression and mediator generation via interaction with IκB kinase. Biochem. Pharmacol., 73: 237-247.
CrossRef - Shin, S.H. and Y.H. Lee, 2010. Airborne fungi induce nasal polyp epithelial cell activation and toll-like receptor expression. Int. Arch. Allergy Immunol., 153: 46-52.
CrossRefDirect Link - Shin, S.H., M.K. Ye, J.K. Kim and K.K. Park, 2013. Bee venom at different concentrations modulates the aeroallergen-induced activation of nasal polyp epithelial cells. Pharmacology, 91: 39-47.
CrossRefDirect Link - Zureik, M., C. Neukirch, B. Leynaert, R. Liard, J. Bousquet and F. Neukirch, 2002. Sensitisation to airborne moulds and severity of asthma: Cross sectional study from European community respiratory health survey. Br. Med. J., 325: 411-414.
CrossRefPubMedDirect Link