
Tajdar Husain Khan
Department of Pharmacology, College of Pharmacy, Salman Bin Abdulaziz University,
P.O. Box 173, Al-Kharj 11942, Kingdom of Saudi Arabia
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 5 | Page No.: 428-433







ABSTRACT
Dietary factors are considered important environmental risk determinants for various diseases. Present study shows the modulatory influence of methanolic extract of Aegle marmelos (AM) on Benzo(a)Pyrene (B(a)P) (125 mg kg-1) induced genotoxicity in Swiss albino mice. The effect of AM was studied by in vivo bone marrow chromosomal aberration and micronuclei induction test. Two doses of AM (25 and 50 mg kg-1 b.wt.) were given orally for seven days prior to the administration of B[a]P. The efficacy of plant extract was also evaluated by an in vitro cytochrome P450 (CYP), DNA-sugar damage in calf thymus DNA and Fe++/ascorbate induced lipid peroxidation (p<0.001) in microsome of mice. Significant increase in the antioxidant activity (p<0.001) and a concomitant decrease (p<0.001) in the malondialdehyde level were observed at three concentrations of plant extract (2.5, 5.0 and 7.5 mg kg-1 b. wt.). Administration of AM, showed inhibitory effects on the genotoxicity of B[a]P in terms of chromosomal aberration and micronucleus formation. The chemopreventive effect of AM on the inhibition of CYP activity and DNA integrity mediate the possible mechanism of inhibition of genotoxicity.
PDF Abstract XML References Citation
Received: February 18, 2012;
Accepted: March 12, 2012;
Published: June 13, 2012
How to cite this article
Tajdar Husain Khan, 2012. Modulatory Effect of Aegle marmelos against Benzo (a) Pyrene B(a)P Induced Genotoxicity in Swiss Albino Mice. International Journal of Pharmacology, 8: 428-433.
DOI: 10.3923/ijp.2012.428.433
URL: https://scialert.net/abstract/?doi=ijp.2012.428.433
DOI: 10.3923/ijp.2012.428.433
URL: https://scialert.net/abstract/?doi=ijp.2012.428.433
INTRODUCTION
Reactive Oxygen Species (ROS) can cause cell damage and initiate various degradation processes (Momtaz and Abdollahi, 2012). Oxidative stress is the result of imbalance between the formation of reactive oxygen species (ROS) and antioxidants in the body which can lead to oxidative damage to biomolecules resulting in lipid peroxidation, mutagenesis and carcinogenesis. Oxidative damage to the DNA plays an important role in carcinogenesis (Hussain et al., 2003; Khan, 2012). Free radical induced lipid peroxidation has been involved in pathogenesis of numerous pathological disorders including cancer (Hristozov et al., 2001). ROS has been associated in numerous diseases, including malignancy, aging and neurodegenerative disorders (Balsano and Alisi, 2009; Momtaz and Abdollahi, 2012).
A promising way to control carcinogenesis or mutagenesis is to prevent the formation of free radical i.e., reactive metabolite from the inert compounds which on interacting or binding covalently with the cellular macromolecule like DNA, lipids or protein may lead to its damage and if not checked or not control may lead to cancer (Ziech et al., 2011; Aggarwal et al., 2009). The peroxidation of lipids, the cross-linking and inactivation of proteins and mutations in DNA are typical consequences of free radicals. Membranes lipids are the main targets of free radicals and they initiate lipid peroxidation chain reaction. A result of lipid peroxidation, membrane fluidity decreased which alters membrane properties (Aggarwal et al., 2009).
Herbs are gaining additional focus because of their less toxicity and high efficacy against a number of ailments. Epidemiological studies have shown that fruits, vegetables, spices, tea and medicinal herbs rich in antioxidants and other micronutrients protect against diverse forms of chemically induced carcinogenesis, inhibit DNA-damage, mutagenesis and lipid peroxidation (Ziech et al., 2011; Birt et al., 2001) Aegle marmelos, known as bael grows in tropical and subtropical parts of the world. Various parts of the AM are used in Indian system of medicine for treatment of many diseases, including diarrhoea, dysentery and dyspeptic symptoms (Shoba and Thomas, 2001; Sharma et al., 2007). Marmelosin, isolated from the AM, has been reported to have anti-helminthic, anti-bacterial, antioxidant activity and anticarcinogenic (Khan and Sultana, 2009; Patil et al., 2010; Khan and Sultana, 2011).
The exposures to genotoxicants have been associated to the expression of biological effects and to increased risk for cancer (Holland et al., 2011). Genotoxicity may cause a long-term effect on the sustainability of particular populations (Bhattacharya, 2011). Genotoxic substances may bring about changes in normal DNA integrity leading to pathological conditions (Holland et al., 2011). DNA damage is unequivocally the main cause of mutagenesis (Preston and Hoffmann, 2001). A frequently occurring form of DNA damage is the DNA strand breaks, divided into single strand breaks (ssBs) and double-strand breaks (dsBs). B(a)P is prevalent in environment, food, ambient and indoor air. Some of its metabolites produced are highly reactive to DNA and may lead to covalent binding causing DNA adduct formation. It has been reported to cause mutations, chromosome aberrations, chromatid exchanges and cancer. Some B(a)P reactive intermediates form alkali-labile sites on DNA (Holland et al., 2011; Paget et al., 2012).
MATERIALS AND METHODS
Chemicals: Calf thymus DNA and Benzo(a)pyrene were obtained from Sigma chemicals Co (St Louis, MO) and all other chemicals were of the highest purity and commercially available.
Animals: Swiss albino mice (20-25 g) were obtained from Central Animal House of Hamdard University, New Delhi, India. They were allowed to acclimatize for one week before the experiments and were given free access to standard laboratory feed (Hindustan Lever Ltd., Bombay, India).
Plant material: The plant material was procured from wholesale spice and herbs market Khari Baoli, old Delhi. Professor Mohammed Iqbal, Medicinal Plant Division, Department of Environmental Botany, Hamdard University, New Delhi verified the identity of plant material. The plant material was chopped and coarsely powdered to a mesh size of 1 mm as described by Antonio and Brito (1998).
Preparation of extract: Powdered plant material was repeatedly extracted in 4000 mL round bottom flask with 2000 mL methanol. The methanolic extracts was cooled at room temperature, filtered and evaporated to dryness under reduced pressure in a rotatory evaporator (Buchi Rotavapor).
In vitro lipid peroxidation: The assay of microsomal lipid peroxidation was done according to the method of Wright et al. (1981). To assess the potential of plant extracts to inhibit lipid peroxidation four different concentration of plant extract were selected. Group I served as a control group in which the reaction mixture consisted of 0.58 mL phosphate buffer (0.1 M, pH 7.4), 0.2 mL ascorbic acid (100 mM) and 0.02 mL ferric chloride (100 mM). In-group II, III, IV, V and IV in addition to the complete control reaction mixture different concentration of plant extract were also added. This reaction mixture was then incubated at 37°C in a shaking water bath for 1 h. The reaction was stopped by the addition of 1 mL of TCA (10%). Following addition of 1.0 mL TBA (0.67%), all the tubes were placed in a boiling water bath for a period of 20 min. The tubes were then centrifuged at 2500 xg for 10 min. The amount of malondialdehyde (MDA) formed in each of the sample was assessed by measuring the optical density of the supernatant at 535 nm. The results were expressed as the nmol MDA formed h-1 g-1 tissue at 37°C by using a molar extinction coefficient of 1.56x105 M-1 cm -1.
DNA sugar damage assay: The DNA sugar damage was assayed by the method of Halliwell and Gutteridge (1981). Group I served as a control group. In control the reaction mixture consisted of 0.5 mL calf thymus DNA (1 mg mL-1 of 0.15 M NaCl), 0.5 mL phosphate buffer (0.1 M, pH 7.4) and 0.05 mL of FeCl3 (100 μM in final concentration). In other groups i.e., Group II, III, IV, V and VI in addition to the above mixture different concentrations of plant extracts were added. The reaction mixture was incubated for 1 h at 37°C in a water bath shaker. After the incubation was over, 1 mL TBA (0.67%) was added to the reaction mixture and then it was kept in boiling water bath for 25 min. The TBA reacting species so generated forms an adduct showing a characteristic absorption at 535 nm which was monitored using spectrophotometer.
Cytochrome P450: Assay of Cyt - P450 content was done by the method of Omura and Sato (1964). To assess the efficacy of AM to inhibition of cytochrome P450 content, 4 groups were chosen. Group I served as control, in which a pinch of sodium dithionate was added to 2 mL of sample. This was then divided equally between matched cuvettes. In addition to the above-mentioned reaction mixture different concentration of plant extract was also added. The contents of the test cuvette were gently bubbled with carbon monoxide for about one minute and the OD was taken at 450 and 490 nm.
Treatment schedule: AM extract was suspended in normal saline and B(a)P in corn oil. B(a)P and Aegle marmelos was administered orally. Each group consisted of five animals. In group I (vehicle control) animals were given normal saline (0.9%) orally. The animals of group II served as positive control and were administered single oral dose of B(a) P (125 mg kg-1 b.wt.). Animals of group III and V were pretreated with 50 mg kg-1 b.wt. of Aegle marmelos while group IV were pretreated with 25 mg kg-1 b.wt. of Aegle marmelos for seven consecutive days. The above-mentioned doses of Aegle marmelos were selected based on the preliminary studies by the investigator. On day 8, the animals of group II, IV and V were administered a single oral dose of B(a)P (125 mg kg-1).
Chromosomal aberration test: Mice were sacrificed by light ether anesthesia 30 h after treatment with B(a)P. About 90 min prior to killing, a single i.p. dose of colchicine (4 mg kg-1) was administered to the animals. The time of sacrifice was decided based on preliminary experiments as optimal for scoring of aberrations. The slides of bone marrow cells were prepared and stained according to the routine schedule for metaphase plate analysis i.e., hypotonic-acetic acid-methanol-flame-drying-Giemsa (Preston et al., 1987). At least 100 well-spread intact metaphases were scored per animal under 100xoil immersion using a light microscope (Olympus BX 50). The type of chromosomal aberration (CA) included chromatid and chromosome breaks and chromosomal rearrangements. All aberrations were considered as equal regardless of the number of breaks involved, gaps were not included. A single observer did blind scoring.
Micronucleus test: For this test, mice were sacrificed 28 h, after treatment with B(a)P. The mouse bone marrow micronucleus test was carried out according to the method of Schmid (1975). The air-dried slides were stained with May-Grunwald and Giemsa as described by Schmid (1975) made permanent and coded. A total of 2500 polychromatic erythrocytes (PCEs) were scored per animal by the same observer for determining the frequencies of micronucleated polychromatic erythrocytes (MnPCEs).
RESULTS
In vitro: Free radical generated by the iron/ascorbate system was inhibited by the addition of plant extracts doses. This inhibition in the formation of malondialdehyde was dose dependent. By the addition of AM extract there was an inhibition in iron ascorbate induced microsomal lipid peroxidation starting from 23-53% respectively as comparison to control group as shown in Fig. 1 and Table 1.
| Table 1: | Percent inhibition values of A. marmelos on lipid peroxidation, cytochrome P-450 content and DNA sugar damage |
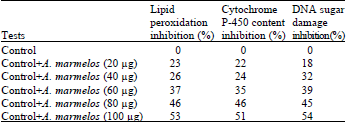 | |
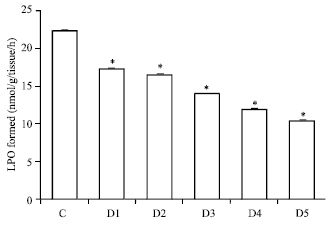 | |
| Fig. 1: | Efficacy of Aegle marmelos in inhibiting iron/ascorbate induced microsomal LPO AM concentration; D1 = 20 μg, D2 = 40 μg, D3 = 60 μg, D4 = 80 μg, D5 = 100 μg, p<0.005 when compared with control |
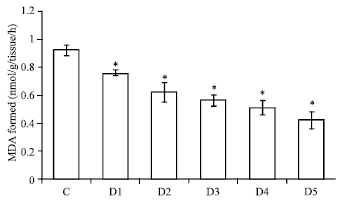 | |
| Fig. 2: | Efficacy of Aegle marmelos towards hydroxyl radical induced DNA-sugar damage AM concentration, D1 = 20 μg, D2 = 40 μg, D3 = 60 μg, D4 = 80 μg, D5 = 100 μg, p<0.005 when compared with control |
All the test groups inhibited DNA sugar damage dose dependently. There was a dose dependent inhibition in DNA sugar damage ranging from 18-54%, respectively. The complete control was assumed to possess 100% DNA damage and 0% inhibition capacity as shown in Fig. 2 and Table 1. The maximum inhibition was observed in-group having highest concentration of plant extract.
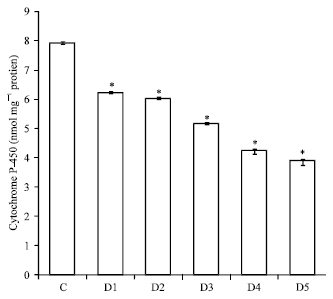 | |
| Fig. 3: | Efficacy of Aegle marmelos on in vitro cytochrome P-450 contents AM concentration, D1 = 20 μg, D2 = 40 μg, D3 = 60 μg, D4 = 80 μg, D5 = 100 μg, p<0.005 when compared with control |
| Table 2: | Effect of Aegle marmelos on chromosomal aberrations induced by B(a)P in mice |
 | |
| B’: Chromatid breaks, B”: Chromosomal breaks, R: Rearrangements, aTotal 500 metaphase plates were observed per group (n = 5 animals) for scoring chromosomal aberrations, *Significantly different from the control group (p<0.001), #Significantly different from the only B(a)P treated group (p<0.05) | |
| Table 3: | Effect of Aegle marmelos on micronuclei induced by B(a)P in mice |
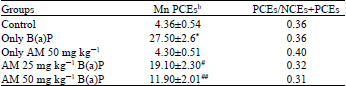 | |
| PCEs: Polychromatic erythrocytes, NCEs: Normochromatic erythrocytes, MnPCEs: micronucleated polychromatic erythrocytes, bMean±SE of 2500 PCEs, *Significantly different from the control group (p<0.001), #Significantly different from the B(a)P treated group (p<0.05), ##Significantly different from the B[a]P treated group (p<0.001) | |
According to our data with AM there is a dose dependent inhibition in the level of cytochrome P450 activity starting from 22-51%. Maximum inhibition was seen in plant having 100 μg concentration of plant extract as shown in Fig. 3 and Table 1.
In vivo: B(a)P treatment alone produced gaps and DNA strand breaks in the cells, However cells with multiple chromosomal aberration and exchanges were observed infrequently as they are not considered as good indicator of chromosomal damage. Table 2 shows the reduction in frequency of chromosomal aberrations induced in mouse bone marrow cells following oral administration of the two doses of Aegle marmelos to the mice for 7 days before B(a)P treatment as compared with the group treated with clastogen, i.e., B(a)P alone. Significant reduction in the frequency of chromosomal aberrations was observed at 25 and 50 mg kg-1 b.wt. doses of AM treatment. Significantly higher incidence of chromosomal aberrations was observed when we compare control group with only B(a)P treated group. In all cases, however, the aberration frequency was lower in the animals given the clastogen alone.
The data presented in Table 3 shows the protective effect of AM against B(a)P induced mutagenicity as assessed by the bone marrow micronucleus test. B(a)P produced significant micronuclei formation when compared with the control group (p<.001) There was 37-57% inhibition of the B(a)P-induced micronuclei by the pretreatment with Aegle marmelos (p<0.001). B(a)P produces micronucleus formation and found significant difference when compared to the control (p<0.001).
DISCUSSION
Cancer and other chronic diseases may be related with mutations produced by environmental agents; therefore, minimizing the exposure to harmful agents has been recommended as a way to prevent these diseases (Aggarwal et al., 2009). Unfortunately, it is not easy to eliminate the source of genotoxic agent completely in modern society. Therefore, the identification and application of well-known antimutagens is essential for improving human health (Siddique and Afzal, 2008). Considerable emphasis had been laid down on the use of dietary constituents to prevent mutagen induced cytogenic damage and DNA damage due to their non-toxic effects. Many plants and their isolated compounds have been tested to determine their antimutagenic potential; like green and black teas are known to be strong mutagenic and carcinogenic inhibitors (Bitiren et al., 2010; Santana-Rios et al., 2001).
Oxidative stress causes a series of chain reaction which is known as lipid peroxidation. Lipid peroxidation causes serious membrane damage and may even lead to cell death (Yassa et al., 2008). It is now widely known that the mutagenic capacity of free radical is due to the direct interaction of hydroxyl radicals (. OH) with DNA. Such interactions induce numerous lesions in DNA that cause deletions, mutations and other genetic effects. Characterization of this damage to DNA has indicated that both the sugar and the base moieties are susceptible to oxidation, causing base degradation, single strand breakage and cross linking to protein (Aggarwal et al., 2009; Momtaz and Abdollahi, 2012). In this study, in vitro lipid peroxidation assay exposes that AM suppress free radical induced lipid peroxidation dose dependently which further may inhibit the process of free radical induce DNA damage.
The deoxyribose assay showed that on interaction of these free radicals with transition metals the hydroxyl radical is produced by Fenton’s reaction which is responsible for DNA damage (Ayene et al., 2007). The dose dependent decrease in the calf thymus DNA sugar damage strengthens our investigation. Our investigation further reveals that AM block the activation of carcinogens by inhibiting Cyt P450 activity which further may inhibit the process of tumorigenesis. Cyt P450 is the major enzyme involved in metabolism of many drugs and xenobiotics. CYP activates Polycyclic Aromatic Hydrocarbon (PAHs) into ultimate metabolites which covalently bind to DNA, a key event in the initiation of carcinogenesis (Srinivasan et al., 2008; Abdel-Latif and Sadek, 1999).
The chromosomal aberrations are produced by error in DNA molecule. Chromosomal aberrations are analyzed in mitotic metaphases from proliferative tissue such as bone marrow tissue. In micronuclei induction test, the clastogenic effects were measured indirectly by counting small nuclei in interphase cells formed by acentric chromosomal fragment or whole chromosomes. In this study, Aegle marmelos have been proven to reduce the micronuclei induction and chromosomal aberration dose dependently.
CONCLUSIONS
Thus, the results from the in vitro and in vivo experiments are quite promising for the use of AM as a protective agent. The anti-clastogenic effects of Aegle marmelos can be attributed to inhibition of lipid peroxidation, DNA sugar damage and cytochrome p450 content in dose dependent manner. It is concluded that Aegle marmelos can be used as a major chemopreventive agent against B[a]P. induced mutagenicity.
ACKNOWLEDGMENT
Dr. Tajdar Husain Khan is thankful to the Indian Council for Medical Research (ICMR), New Delhi, India, for providing the funds to carry out this study.
REFERENCES
- Abdel-Latif, H. and I.A. Sadek, 1999. Curcumin and its effect on cytochrome P450 and GST in toad liver tumor induced by DMBA. Pak. J. Biol. Sci., 2: 1193-1196.
CrossRefDirect Link - Ayene, I.S., C.J. Koch and R.E. Krisch, 2007. DNA strand breakage by bivalent metal ions and ionizing radiation. Int. J. Radiation Biol., 83: 195-210.
PubMed - Balsano, C. and A. Alisi, 2009. Antioxidant effects of natural bioactive compounds. Curr. Pharm. Des., 15: 3063-3073.
PubMed - Bhattacharya, S., 2011. Natural antimutagens: A review. Res. J. Med. Plant, 5: 116-126.
CrossRefDirect Link - Birt, D.F., S. Hendrich and W. Wang, 2001. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther., 90: 157-177.
CrossRef - Bitiren, M., D. Musa, A. Ozgonul, M. Ozaslan and A. Kocyigit et al., 2010. Protective effects of green tea (Camelia sinensis), Hypericum perforatum and Urtica dioica on hepatic injury and lymphocyte DNA damage induced by carbon tetrachloride in wistar rats. Int. J. Pharmacol., 6: 241-248.
CrossRefDirect Link - Halliwell, B. and J.M.C. Gutteridge, 1981. Formation of a thiobarbituric-acid-reactive substance from deoxyribose in the presence of iron salts: The role of superoxide and hydroxyl radicals. FEBS Lett., 128: 347-352.
CrossRefPubMedDirect Link - Hristozov, D., V. Gadjeva, T. Vlaykova and G. Dimitrov, 2001. Evaluation of oxidative stress in patients with cancer. Arch. Physiol. Biochem., 109: 331-336.
CrossRefPubMedDirect Link - Hussain, S.P., L.J. Hofseth and C.C. Harris, 2003. Radical causes of cancer. Nat. Rev. Cancer, 3: 276-285.
CrossRefDirect Link - Khan, T.H., 2012. Soy diet diminish oxidative injure and early promotional events induced by CCl4 in rat liver. Int. J. Pharmacol., 8: 30-38.
CrossRefDirect Link - Khan, T.H. and S. Sultana, 2009. Antioxidant and hepatoprotective potential of Aegle marmelos Correa. against CCl4-induced oxidative stress and early tumor events. J. Enzyme Inhib. Med. Chem., 24: 320-327.
CrossRefPubMedDirect Link - Khan, T.H. and S. Sultana, 2011. Effect of Aegle marmelos on DEN initiated and 2-AAF promoted hepatocarcinogenesis: A chemopreventive study. Toxicol. Mech. Methods, 21: 453-462.
CrossRef - Momtaz, S. and M. Abdollahi, 2012. A comprehensive review of biochemical and molecular evidences from animal and human studies on the role of oxidative stress in aging: An epiphenomenon or the cause. Asian. J. Anim. Vet. Adv. 7: 1-19.
CrossRef - Omura, T. and R. Sato, 1964. The carbon monoxide binding pigment of liver microsomes. 1- Evidence for its hematoprotein nature. J. Biol. Chem., 239: 2370-2378.
PubMed - Paget, V., M. Lechevrel, V. Andre, J. Le Goff and D. Pottier et al., 2012. Benzo[a]pyrene, aflatoxine B1 and acetaldehyde mutational patterns in TP53 gene using a functional assay: Relevance to human cancer aetiology. PLoS ONE, Vol. 7, No. 2.
CrossRefDirect Link - Patil, D.N., A.R. Kulkarni and B.S. Patil, 2010. Fruit gum of Aegle marmelos as pharmaceutical aid. Int. J. Pharmacol., 6: 68-71.
CrossRefDirect Link - Preston, R.J., B.J. Dean, S. Galloway, H. Holder, A.F. McFee and M. Shelby, 1987. Mammalian in vivo cytogenetic assay: Analysis of chromosome aberration in bone marrow cell. Mutation Res., 189: 157-165.
PubMed - Santana-Rios, G., G.A. Orner, A. Amantana, C. Provost, S.Y. Wu and R.H. Dashwood, 2001. Potent antimutagenic activity of white tea in comparison with green tea in the Salmonella assay. Mutat. Res., 495: 61-74.
PubMed - Sharma, B., S.K. Satapathi and P. Roy, 2007. Hypoglycemic and hypolipidemic effect of Aegle marmelos (L.) leaf extract on streptozotocin induced diabetic mice. Int. J. Pharmacol., 3: 444-452.
CrossRefDirect Link - Shoba, F.G. and M. Thomas, 2001. Study of antidiarrhoeal activity of four medicinal plants in castor-oil induced diarrhoea. J. Ethnopharmacol., 76: 73-76.
CrossRefDirect Link - Siddique, Y.H. and M. Afzal, 2008. A review on the genotoxic effects of some synthetic progestins. Int. J. Pharmacol., 4: 410-430.
CrossRefDirect Link - Srinivasan, P., S. Suchalatha, P.V. Babu, R.S. Devi, S. Narayan, K.E. Sabitha and C.S.S. Devi, 2008. Chemopreventive and therapeutic modulation of green tea polyphenols on drug metabolizing enzymes in 4-Nitroquinoline 1-oxide induced oral cancer. Chem. Biol. Interact., 172: 224-234.
CrossRefPubMedDirect Link - Yassa, N., H. Razavi Beni and A. Hadjiakhoondi, 2008. Free radical scavenging and lipid peroxidation activity of the shahani black grape. Pak. J. Biol. Sci., 11: 2513-2516.
CrossRefPubMedDirect Link - Ziech, D., R. Franco, A. Pappa and M.I. Panayiotidis, 2011. Reactive Oxygen Species (ROS)-induced genetic and epigenetic alterations in human carcinogenesis. Mutation Res., 711: 167-173.
CrossRefPubMedDirect Link - Antonio, M.A. and A.R.M.S. Brito, 1998. Oral anti-inflammatory and anti-ulcerogenic activities of a hydroalcoholic extract and partitioned fractions of Turnera ulmifolia (Turneraceae). J. Ethnopharmacol., 61: 215-228.
CrossRefPubMedDirect Link