
Maysam Mard-Soltani
Department of Biology, Faculty of Science, Shahid Chamran University, Ahvaz, Iran
Mahnnaz Kesmati
Department of Biology, Faculty of Science, Shahid Chamran University, Ahvaz, Iran
Lotfolah Khajehpour
Department of Biology, Faculty of Science, Shahid Chamran University, Ahvaz, Iran
Abdolrahman Rasekh
Department of Statistics, Faculty of Mathematics and Computer Science, Shahid Chamran University, Ahvaz, Iran
Abdolhosein Shamshirgar-Zadeh
Medical Education Development Center, Dezfoul University of Medical Sciences, Dezfoul, Iran
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 5 | Page No.: 344-354







ABSTRACT
Many studies have shown that anxiety-like behaviors are influenced by peripheral and central parameters including hormones and neurotransmitters in the different areas of Central Nervous System (CNS). Many investigations have proven the anxiolytic effect of androgens in different methods. Also, there are many reports about the modulating role of the amygdala complex, especially Basolateral Amygdala (BLA), through adrenergic system on anxiety. Among various types and subtypes of adrenergic receptors, β-1 adrenoceptors (β-1 ARs) of BLA account for the anxiety. The purpose of this study was to examine the relationship between testosterone and β-1 ARs in the BLA as an anxiety regulating key of CNS. Using Elevated plus Maze (EPM), the anxiety-like behaviors of four groups of intact adult male Wistar rat were assessed in the presence of different doses of testosterone (0, 20, 30 and 40 mg kg-1, Intraperitoneal (i.p.). Then, the effects of intra-BLA microinjection of different doses of betaxolol, a selective β-1 ARs antagonist, (0, 0.025, 0.1 and 0.4 μg rat-1), were evaluated in the other four groups. Finally, the interaction between the ineffective dose of testosterone via i.p. and betaxolol via intra-BLA was investigated. The results obtained revealed that testosterone (i.p.) and betaxolol (intra-BLA) alone had anxiolytic effects on the male Wistar rats in a dose-dependent manner. Our findings also showed that the anxiolytic effects of testosterone (i.p.) were reinforced by the intra-BLA injection of betaxolol. Co-administration betaxolol and testosterone also showed the synergistic actions on anxiolytic effects in the adult male Wistar rats. Our results interestingly proposed that the interaction between testosterone and β-1 ARs is in part related to common mechanism and other neurotransmitters include gamma-aminobutyric acid (GABA) and serotonin.
PDF Abstract XML References Citation
Received: November 20, 2011;
Accepted: March 27, 2012;
Published: June 13, 2012
How to cite this article
Maysam Mard-Soltani, Mahnnaz Kesmati, Lotfolah Khajehpour, Abdolrahman Rasekh and Abdolhosein Shamshirgar-Zadeh, 2012. Interaction between Anxiolytic Effects of Testosterone and β-1 Adrenoceptors of Basolateral Amygdala. International Journal of Pharmacology, 8: 344-354.
DOI: 10.3923/ijp.2012.344.354
URL: https://scialert.net/abstract/?doi=ijp.2012.344.354
DOI: 10.3923/ijp.2012.344.354
URL: https://scialert.net/abstract/?doi=ijp.2012.344.354
INTRODUCTION
Various humoral and neurochemical substances are involved in anxiety-like behaviors (Samad et al., 2005; Raj et al., 2006; Boyle et al., 2006; Velisek, 2006; Edinger and Frye, 2006; Sadeghi et al., 2007; Blum et al., 2007; Taiwe et al., 2010; Naghibi et al., 2011; Mahbub-E-Sobhani et al., 2011). Anxiety are reduced by anxiolytic drugs and controlled by two large limbic system structures, the amygdala and hippocampus (David, 1999; Wang et al., 2011). The amygdala complex seems to be in charge of mediating anxiety in both humans and animals (Killcross et al., 1997). This complex has a crucial role in regulating anxiety and emotional responses (Wang et al., 2011). The lateral and Basolateral Amygdala (BLA) are known to be of great importance for integrative processes and are the primary input nuclei of amygdala (Heldt and Ressler, 2006; Shibata et al., 1989). Adrenergic system and other neurotransmitters play an important role in the BLA on modulating the anxiety-like behaviors (Quirarte et al., 1997; Roozendaal et al., 2006; Shibata et al., 1989). Among various types and subtypes of adrenergic receptors, β-Adrenoceptors (β-ARs) of BLA are involved in the anxiety-like behaviors (Rudoy and Van Bockstaele, 2007; Fu et al., 2008). However, some studies suggest that various subtypes of β-ARs including β-1, β-2 and β-3 may exert different effects on anxiety (Ordway et al., 1991; Pandey et al., 1995). Some studies have shown that selective β-2 ARs antagonist appears to be ineffective in treating acute anxiety while it is effective in treating chronic anxiety. Other investigations have shown that β-1 ARs expression in rats’ amygdala increased in cocaine withdrawal-induced anxiety (Rudoy and Van Bockstaele, 2007; Fu et al., 2008).
On the other hand, the anxiolytic effect of testosterone has been found in many studies on rodents. Aikey et al. (2002) indicated that testosterone reduces anxiety-like behaviors rapidly in male house mice. Clark and Henderson (2003) showed that testosterone propionate decreased anxiety in adult male rat in Elevated Plus-maze (EPM).
There is a suggestion showing that the mechanism of testosterone in producing anxiolytic effects may be related, in part, to its metabolism. Testosterone is metabolized into Dihydrotestosterone (DHT) by 5α-reductase which activates the androgen responses. Then, the DHT metabolized into 3α-androstenediol (3α-diol) by 3α-hydroxysteroid dehydrogenase. Testosterone and DHT enjoy a high affinity for intracellular androgen receptors (Verhoeven et al., 1975; Cunningham et al., 1979; Christiansen and Knussmann, 1987; Frye et al., 2002, 2007) but 3α-Diol does not (Roselli, 1991). Androgen regimes, that elevate 3α-Diol concentrations significantly, increase the duration of open arm time in the EPM (Gee, 1988; Frye et al., 1996; Frye and Reed, 1998; Monjo et al., 2003; Fernandez-Guasti and Martinez-Mota, 2005). GABAA/benzodiazepine Receptors (GBRs) complexes have a key role in modulation of anxiety. 3α-Diol is a potent modulator at GABAA/benzodiazepine receptors (GBRs), whereas both testosterone and DHT have a weak activity in this regard (Gee, 1988; Frye et al., 1996; Frye and Reed, 1998; Fernandez-Guasti and Martinez-Mota, 2005). Some studies suggest that testosterone reduces serotonin level in the brain area and anti-anxiety effects of testosterone are possibly done by reducing serotonin level (Birger et al., 2003). Up-regulation of serotonin in the brain area such as BLA causes anxiety (Van der Wee et al., 2008; Akimova et al., 2009).
Until now, the relationship between the anxiolytic effects of testosterone and BLA β-1 ARs is not clearly understood. Accumulating evidence suggests that ARs subtypes display a differential regulation by testosterone (i.e. up-regulation of β-1 ARs and down-regulation of β-2 and β-3 ARs) (Monjo et al., 2003). It has been found that β-1 ARs mRNA levels in different tissue are evaluated when all the sexual hormones are injected (Monjo et al., 2003). We therefore hypothesized that existed interaction between anxiolytic effects of testosterone and β-1 ARs of BLA. This study has been designed to examine the effect of testosterone in the presence and absence of BLA β-1 ARs on anxiety-like behaviors in the EPM and to reveal the interaction between them in intact adult male rats.
MATERIALS AND METHODS
All procedures were carried out in accordance with the Institutional Guidelines for Animal Care and Use of Laboratory Animals and approved by the Biology Department of Shahid Chamran University (Ahvaz, Khuzestan Province, Iran).
Animals: Intact male Wistar rats (purchased from Jondi Shapour University of Ahwaz, Khuzestan province, Iran), aged 13±2 weeks and weighing 200±20 g, at the time of surgery were used. The animals were housed four/cage in a colony room with a constant 12 h reverse-light/dark cycle (7:00 AM-19:00 PM light off) at 22±1°C and relative humidity of 30-50%. The animals had free access to food and water except during the time of behavioral test session. All animals were allowed to adapt with the laboratory conditions for at least 1 week before the surgery. The rats were handled about 5 min each day prior to behavioral testing. In our study, rats are always tested in their dark phase (between 9:00 AM and 14:00 PM), when rodents are most active and have consistent differences in their endogenous concentrations of androgens. Each animal was used once only. Eight animals were used in each group of the experiments and all animals had stereotaxic surgery stress.
Stereotaxic surgery: The rats were anesthetized intraperitoneally (i.p.) using ketamine hydrochloride (50 mg kg-1) and xylazine (4 mg kg-1) and placed in a stoelting stereotaxic instrument (Stoelting Co, Illinois, USA). The animals head was restrained in a stereotaxic apparatus. Two Guide cannulas (14 mm stainless steel, 21 gauge) were implanted bilaterally into BLA (from bregma 2.8 mm caudally, 5 mm laterally and from skull surface 6.8 mm ventrally). A stylus (15 mm stainless steel, 27 gauge) was placed in the guide cannulae to prevent coagulation of clogging. After recovery from anesthesia, the rats were returned to their home cage for one week before behavioral test session.
Drugs: Testosterone [testosterone enanteat 250 mg mL-1, Abureihan Co., Tehran, Iran] was dissolved in sesame oil (testosterone vehicle manufactured by Barvich Co., Tehran, Iran). The final testosterone concentrations were 0, 20, 30 and 40 mg/animal kg. The selective β-1 ARs antagonist, betaxolol (betaxolol hydrochloride, Tocris Bioscience, IO Center Moorend Farm Avenue, Bristol BS 11, OL, UK) was dissolved in 0.9% sodium chloride saline (as betaxolol vehicle) and the final concentrations were 0, 0.025, 0.1 and 0.4 μg rat-1 (Cecchi et al., 2007).
Drug injection: For intra-BLA betaxolol or betaxolol vehicle injections, the stylus was withdrawn into the guide cannula and replaced by the injection unit (15 mm stainless steel 27 gauge tubing), terminating at 1.5 mm below the tip of the guide cannula (Cecchi et al., 2007). Each injection unit has been connected to a 2 μL Hamilton syringe by polyethylene tubing. The animals received an injection of 1 μL rat-1 intra-BLA (0.5 μL/per cannula) at a 60 sec period. The inner cannula was left in place for an additional 60 sec to allow diffusion of the drug or vehicle and to reduce the possibility of reflux. 15 min after intra-BLA microinjection, testosterone or vehicle was injected via i.p. (ElAttar et al., 1964). Then, the behavioral testing was done 45 min post i.p. injection.
Behavioral testing: The EPM paradigm was used effectively to assess anxiety and exploratory behaviors (Walf and Frye, 2007; Matuszewich et al., 2007). The EPM consisted of four arms (50 cm long and 10 cm wide) elevated 73 cm off the ground. Two arms were enclosed by 30 cm high walls and the other two arms were exposed. As described by Pellow and File (1986) and Dunn et al. (1989), the rats were placed at the junction of the open and closed arms of the EPM and observed for 5 min. The number of entries and the amount of time spent on the open and closed arms were assessed using a video device (Sony Handycam HDR-CX110 Camcorder-1080i). The rats were considered to be in either the closed or the open arms of the EPM and the open arm time was recorded only when the rats had all four paws on the open arms of the EPM. Numbers of total arm entries reflect the motor component of the exploratory activity (locomotors activity). The apparatus was cleaned with alcohol after each rat was tested.
Experimental design:
| • | Experiment 1: Effects of testosterone alone on the anxiety-like behaviors in adult male rats: In this stage, four groups of animals (control or 0, T20, T30 and T40) received betaxolol vehicle (normal saline 1 μL rat-1 intra-BLA) and then 15 min post-intra-BLA injections, each group received testosterone 0, 20, 30 and 40 mg kg-1, i.p. ,respectively. The behavioral test session was performed 45 min after the i.p. injection and the percent of open arm time (OAT%), the percent of open arm entries (OAE%) and locomotors activity were assessed (Fig. 2) |
| • | Experiment 2: Effects of betaxolol alone on the anxiety-like behaviors in adult male rats: In this stage, four groups of animals (control or 0, B0.025, B0.1 and B4) received betaxolol 0, 0.025, 0.1 and 0.4 μg rat-1 intra-BLA, respectively. Then, 15 min later, the all these groups received testosterone vehicle (1 mL kg-1 sesame oil via i.p.). The behavioral test session was performed 45 min after the i.p., injection and OAT%, OAE% and locomotors activity were assessed (Fig. 3) |
| • | Experiment 3: Interaction of betaxolol and testosterone on the anxiety-like behaviors: In this stage, using the one-way ANOVA follow up by Turkeys’ test of prior experiments, one group of rats (BT group) received an ineffective dose of betaxolol (0.025 μg rat-1 via intra-BLA) and, 15 min later, an ineffective dose of testosterone (20 mg kg-1, via i.p.). The behavioral test session was performed 45 min post i.p. injection and OAT%, OAE% and locomotors activity were assessed. This group was compared with control, T20 and B0.025 groups (Fig. 4) |
Verification of cannula placement: After completion of all experimental sessions, the animals were killed with an overdose of chloroform. Subsequently, 0.5 μL per cannula of ink (0.1% aquatic methylene blue solution) was injected intra-BLA by a 15 mm stainless steel 27 gauge which was projected a further 1.5 mm ventral to the tip of the guide cannula to aid in histological verification. The animals' brain was removed and fixed in 10% formalin 10 days before sectioning. All sections were examined to determine the location of the cannula aimed for BLA. The cannula placement was verified using the Atlas of Paxinos and Watson (1998). The data from rats with cannula placement outside the BLA were excluded from the analyses.
Statistical analyses: Statistical analyses were performed using the Statistical Package for the Social Science (SPSS-PC, version 15. SPSS, Inc., Chicago, IL). The data were expressed as Mean±SEM. The data analysis for the experiments 1, 2 and 3 was performed by one-way analysis of variance (ANOVA). Then a post-hoc analysis (Tukey) was performed for assessing specific group comparisons. The differences between the experimental groups at each point were considered as statistically significant (p<0.05).
RESULTS
Histology: Figure 1 shows the approximate point of drug injection intra-BLA. The histological results were plotted on the representative section taken from the rat brain (2.80 mm posterior to bregma; Paxinos and Watson (1998). The data from the animals with the injection sites located outside the BLA were not used in the analysis.
Effect of testosterone alone on the anxiety-like parameters: Figure 2 shows the effects of i.p. injection of different doses of testosterone (0, 20, 30 and 40 mg kg-1) on the anxiety-related parameters in the EPM.
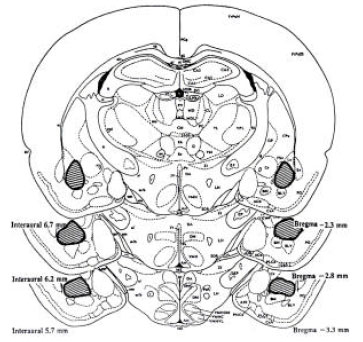 | |
| Fig. 1: | Schematic illustration of coronal section of the rat brain (2.80 mm posterior to bregma; Paxinos and Watson (1998) showing the approximate location of the BLA sites in the study |
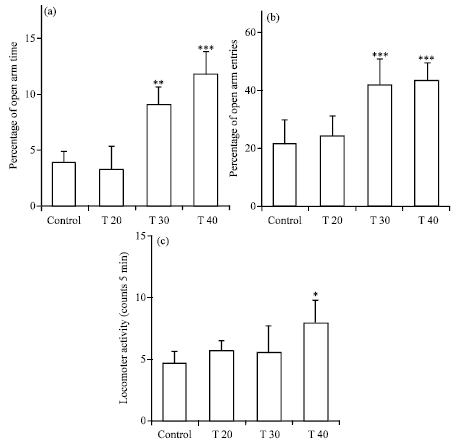 | |
| Fig. 2(a-c): | Anxiolytic effects of testosterone on the performance of rats in the EPM. (a) OAT%, (b) OAE% and (c) Locomotor activity (number of total arm entries). Betaxolol vehicle (1 μL rat-1 Saline via intra-BLA) was microinjected 15 min before the i.p., infusion of different doses of testosterone (0, 20, 30 and 40 mg kg-1). The behavioral test session was performed 45 min after i.p., injection. The data were expressed as Mean±SEM. *p<0.05, **p<0.01 and ***p<0.001 compared with control group. n = 8 for all groups |
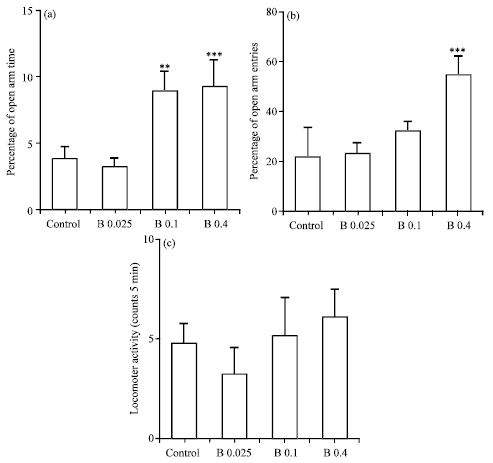 | |
| Fig. 3(a-c): | Anxiolytic effects of betaxolol on the performance of rats in the EPM. (a) OAT%, (b) OAE% and (c) Locomotor activity (No. of total arm entries). Different doses of betaxolol (0, 0.025, 0.1 and 0.4 μg rat-1) were infused 15 min before the i.p. injection of testosterone vehicle (1 mL kg-1 sesame oil via i.p.). The behavioral test session was performed 45 min after i.p. injection. The data were expressed as Mean±SEM, ** p<0.01 and ***p<0.001 compared with control group. n = 8 for all groups |
A one-way ANOVA follow up by Turkeys’ test revealed that anxiety-related parameters were altered by testosterone in comparison with the control group: OAT% [T20 = 3.38±2, p<0.05; T30 = 9.14±1.6, p<0.01; T40 = 11.85±2, p<0.001] (Fig. 2a), OAE% [T20 = 24.4±7, p>0.05; T 30 = 42±9, p<0.001; T40 = 43.42±6, p<0.001] (Fig. 2b) and locomotor activity [T20 = 4.57±1.95, p>0.05; T30 = 5.57±2.2, p>0.05; T40 = 8.00±1.7, p<0.05] (Fig. 2c). These results revealed that testosterone has an anxiolytic effect in a dose-dependent manner. Control group received betaxolol vehicle (1 μL rat-1 saline via intra-BLA) and testosterone vehicle (1 mL kg-1 sesame oil via i.p.). The anxiety-related parameters in control group: OAT% = 4±0.91; OAE% = 22±6; locomotor activity = 4.71±0.95. The data were expressed as Mean±SEM n = 8 for all groups.
Effects of intra-BLA injection of betaxolol on the anxiety-like parameters: Figure 3 shows the intra-BLA injection effects of betaxolol (0, 0.025, 0.1 and 0.4 μg rat-1 via intra-BLA) on the anxiety-related parameters. One-way ANOVA follow up by Turkey’s test revealed that betaxolol also alters anxiety-related parameters in the EPM in a dose dependent manner and reduces the animals’ anxiety. In comparison with the control group, betaxolol significantly altered the OAT% [B0.025 = 3.14±0.75, p>0.05; B0.1 = 8.86±1.50, p<0.01; B0.4 = 9.16±2.00, p<0.001] (Fig. 3a) and the OAE% [B0.025 = 23.25±4.00, p>0.05; B0.1 = 31.14±3.50, p>0.05; B0.4 = 55.71±8.00, p<0.001] (Fig. 3b) but it had no effects on the animals’ locomotor activity [B0.025=3.28±1.26, P>0.05; B0.1 = 5.14±1.86, p>0.05; B0.4 (7,56) = 6.00±1.47, p>0.05] (Fig. 3c). As mentioned earlier, control group received betaxolol vehicle (1 μL rat-1 saline via intra-BLA) and testosterone vehicle (1 mL kg-1 sesame oil via i.p.). The data were expressed as Mean±SEM n = 8 for all groups.
Interaction of testosterone i.p. injection and betaxolol intra-BLA injection on the anxiety-like parameters: Figure 4 shows the effects of betaxolol (0.025 μg rat-1 via intra-BLA) on the anxiolytic effects of testosterone (20 mg kg-1 via i.p.) in the anxiety-related parameters.
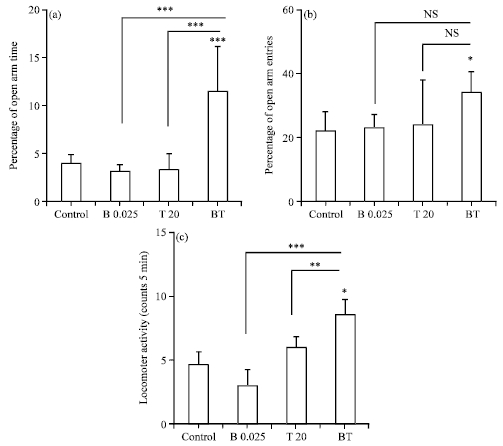 | |
| Fig. 4(a-c): | Interaction of testosterone and betaxolol on the performance of rats in the EPM. (a) OAT%, (b) OAE% and (c) Locomotor activity (No. of total arm entries). BT group received an ineffective dose of betaxolol (0.025 μg rat-1 intra-BLA) and an ineffective dose of testosterone (20 mg kg-1, 15 min later intra-BLA injection via i.p.). The behavioral test session was performed 45 min after i.p. injection. The data were expressed as Mean±SEM *p<0.05, **p<0.01 and ***p<0.001 and BT group compared with the control, T20 and B0.025. The anxiety-related parameters of T20 and B0.025 groups were expressed in previous sections. n = 8 for all groups |
A one-way ANOVA follow up by Turkey’s test revealed that the BT group in comparison with the control, T20 and B0.025 groups has a significant difference on the anxiety-related parameters in the EPM. In the previous experiments, betaxolol (0.025 μg rat-1 via intra-BLA) and testosterone (20 mg kg-1 via i.p.) that had no effect alone on the anxiety-like behaviors, when used together, have an interactive and significant anxiolytic effect on the OAT%, OAE% and locomotor activity. Co-administration of ineffective dose of betaxolol and testosterone (BT group) in comparison with control group significantly altered the OAT% [BT = 12.14±4.68, p<0.001] (Fig. 4a) and OAE% [BT = 34.00±6.80, p<0.05] (Fig. 4b) and locomotion activity [BT = 8.28±1.97, p<0.01] (Fig. 4c). The data were expressed as Mean±SEM.
DISCUSSION
The EPM was employed as an animal model for studying the anxiety-like behaviors (Sullivan et al., 2009; Woode et al., 2009, 2010; Zuloaga et al., 2011; Skurlova et al., 2011). Our findings showed that testosterone has an anxiolytic effect with dose dependent manner in the intact adult male rats. Also the inhibition of BLA β-1 ARs by betaxolol significantly decreased the anxiety-like behaviors in the EPM.
The results of this study also demonstrated that there is an interaction and synergic actions between testosterone and BLA β-1 ARs antagonist. We suggest that the anxiolytic effect of testosterone might have occurred in BLA, because microinjection of betaxolol into BLA reinforces the anxiolytic effects of testosterone.
In support of our findings, the anxiolytic effects of testosterone have been shown in many investigations on rodents. Aikey et al. (2002) indicated that testosterone rapidly reduces anxiety but not locomotor activity in male house mice, Clark and Henderson (2003) showed that testosterone propionate decreases anxiety in adult male rats in the EPM. Some studies suggest that androgen administration can have anxiolytic effects in both male and female rats (Forman et al., 1989; Bitran et al., 1993; Molina et al., 1994).
Testosterones mechanism to produce analgesia and to enhance cognitive performance may be in part due to its metabolism. Testosterone is metabolized into DHT by 5α-reductase which is, in turn metabolized by 3α-hydroxysteroid dehydrogenase into 3α-Diol, as described in more detail in the introduction part. Testosterone and DHT have a high affinity for intracellular androgen receptors (Verhoeven et al., 1975; Cunningham et al., 1979; Christiansen and Knussmann, 1987; Frye et al., 2002, 2007) but 3α-Diol does not (Roselli, 1991). 3α-Diol is a potent modulator at GBRs, whereas testosterone and DHT have only weak activity at GBRs (Gee, 1988; Frye et al., 1996; Frye and Reed, 1998; Fernandez-Guasti and Martinez-Mota, 2005). Androgens can change GBR pharmacodynamics, as gonadectomy decreases sensitivity to diazepam-induced sedation (Svensson et al., 2000). The similar effects of testosterone, DHT and 3α-Diol to produce analgesia and anxiolytic effects suggest that their actions may stem from their metabolism or production of 3α-Diol and subsequent actions at GBRs (Frye and Lacey, 2001). For example, administration of 3α-Diol to Ovariectomized (OVX) rats produced analgesia in the tail-flick model (Frye et al., 1996). Androgen regimes that increase 3α-Diol concentrations significantly increase the OAT% in the EPM (Monjo et al., 2003). Our findings also confirmed the anxiolytic effects of testosterone in the intact adult male rats.
In addition, the results of this study indicated that administration of antagonist β-1 ARs in the BLA has anxiolytic effects and does not have any influence upon locomotor activity. It has been also confirmed that the amygdala receives sever noradrenergic innervations (Clayton and Williams, 2000) and down regulation of adrenergic system in long or short term may occur after stressor cassation. For example, stress can severely and lastingly impair the function of α-1 ARs in the BLA (Braga et al., 2004). β-1 ARs mainly exist in postsynaptic membrane and known to be coupled to Gs (Stimulatory G protein), such that stimulation of these receptors increases cyclic intracellular adenosine monophosphate (cAMP) levels via a direct activation of adenylate cyclase. Immunofluorescence studies have also demonstrated that β-1 ARs were located in the cell membrane and the number of immunoreactive neurons was significantly increased in the amygdala of the fear conditioned rats (Beitner-Johnson and Millhorn, 1998; Rosen and Schulkin, 1998).
Fu et al. (2008), using western blotting analysis, showed up-regulation of β-1 ARs in the amygdala after fear training and anxiety condition. Also it was confirmed that microinjection of metoprolol as antagonist β-1 ARs in the rats BLA inhibits up-regulation of β-1 ARs.
Furthermore, our results obviously revealed that there exists an interaction and synergic action between testosterone and BLA β-1 ARs on the anxiety-like behaviors. The administration of either the lowest dose of testosterone (i.p.) or betaxolol (intra-BLA) alone did not have any effects on the anxiety-related parameters. Co-administration of testosterone and betaxolol attenuated the anxiety-like behaviors in the BT group in comparison with control, B0.025 and T20 groups. This proves the synergistic actions of testosterone and β-1 ARs antagonist on the anxiety. There are miscellaneous reports showing the intervention of androgen hormones, especially testosterone in ARs regulation. This interaction between testosterone and BLA β-1 ARs probably depends on the common intracellular signaling pathway or common neurochemical system in the BLA. One mechanism of synergic actions of testosterone and betaxolol on the anxiety-like parameters might be occurring through modulation of serotonergic and GABAergic systems. For example, activity of projection neurons in the BLA is under strong inhibitory control of GABAergic synaptic transmission (Royer et al., 1999; Szinyei et al., 2000) and intensive anxiety attenuates inhibitory GABAergic control in the BLA (Braga et al., 2002; Rodriguez Manzanares et al., 2005). Other studies have indicated that the dorsal raphe nucleus serotonergic system is a critical modulator of the GABAergic system in the BLA and dysregulation of this system has been recognized in stress and anxiety disorders (Southwick et al., 1999; Van Praag, 2004).
Some studies have shown that up-regulation of serotonin in the brain regions (including the amygdala) caused anxiety and the serotonin levels are strongly enhanced in the BLA during stressful experiences (Amat et al., 1998; Minor and Hunter, 2002). Therefore, manipulation of serotonergic system within the BLA has a critical influence on emotional and anxiety-related parameters (Kim et al., 2005; Van Nobelen and Kokkinidis, 2006). It seems that both testosterone and betaxolol decreases the concentration of serotonin in various brain nuclei including the BLA.
In support of this finding, other studies have shown that β-AR antagonists inhibit serotonin uptake by pulmonary vascular cells in culture and this condition attenuates serotonin function (Lee and Fanburg, 1991). Further, serotonin level in the pineal gland decreases due to the effect of specific β-1 and β-2 AR antagonists (Brownstein et al., 1973). In this study, betaxolol, as a selective β-1 ARs antagonist, probably tends to have affinity for serotonin BLA receptors. Other studies have confirmed that testosterone reduces serotonin level in the brain regions and anxiolytic effects of testosterone are possibly taken place by reducing serotonin level (Birger et al., 2003). According to the findings of this study, testosterone and selective β-1 ARs antagonist likely reduce serotonin function or concentration in the BLA and thus testosterone hormone and betaxolol have an interaction and anxiolytic effects. However, the effect of specific β-1 ARs antagonist occurs in the BLA and betaxolol increases the anxiolytic effects of testosterone in this region.
The second mechanism that it can also be suggested is the role of GABAergic system alone on this synergic effect. Some studies have shown that adrenergic systems, especially α-1A ARs, generally impair facilitation of GABAergic transmission in the BLA (Braga et al., 2004). Further β-1 ARs mediate the facilitation of Purkinje cell in response to GABA in the cerebellum of rats (Yeh and Woodward, 1983). Therefore, betaxolol as β-1 ARs antagonist probably attenuates this system and stops the inhibitory effect of GABAergic system in the BLA. But testosterone and particularly its metabolites have a high affinity to GBRs and reinforce the GABAergic system. Thus it is likely that in contrast to testosterone, selective β-1 ARs antagonists naturally inhibit GABAergic system. It is in contrast to our finding. However, β-1 ARs have a high heterogeneity in different brain regions and the precise mechanism between the all subtypes of β-1 ARs in the BLA and GABAergic system is not clearly understood. Further study is needed to investigate the modulatory effects of GABAergic system in this synergic action.
Finally, we conclude the findings of this study confirmed the synergistic actions of testosterone and betaxolol, β1-AR antagonist, on the anxiolytic effects in the BLA. Also, our results interestingly proposed that the interaction between testosterone and β-1 ARs is probably in part related to common neurotransmitter systems include serotonergic or GABAergic systems. Further study is needed to investigate the precise mechanism of synergistic action of betaxolol and testosterone in the BLA.
ACKNOWLEDGMENT
The financial and technical support of Shahid Chamran University of Ahvaz, Iran, is acknowledged.
REFERENCES
- Aikey, J.L., J.G. Nyby, D.M. Anmuth and P.J. James, 2002. Testosterone rapidly reduces anxiety in male house mice (Mus musculus). Horm. Behav., 42: 448-460.
CrossRefDirect Link - Akimova, E., R. Lanzenberger and S. Kasper, 2009. The serotonin-1A receptor in anxiety disorders. Biol. Psychiatry, 66: 627-635.
CrossRefDirect Link - Amat, J., P. Matus-Amat, L.R. Watkins and S.F. Maier, 1998. Escapable and inescapable stress differentially alter extracellular levels of 5-HT in the basolateral amygdala of the rat. Brain Res., 812: 113-120.
CrossRefDirect Link - Beitner-Johnson, D. and D.E. Millhorn, 1998. Hypoxia induces phosphorylation of the cyclic AMP response element-binding protein by a novel signaling mechanism. J. Biol. Chem., 273: 19834-19839.
CrossRefDirect Link - Birger, M., M. Swartz, D. Cohen, Y. Alesh, C. Grishpan and M. Kotelr, 2003. Aggression: The testosterone-serotonin link. Isr. Med. Assoc. J., 5: 653-658.
PubMedDirect Link - Bitran, D., C.K. Kellogg and R.J. Hilvers, 1993. Treatment with an anabolic-androgenic steroid affects anxiety-related behavior and alters the sensitivity of cortical GABAA receptors in the rat. Horm. Behav., 27: 568-583.
CrossRefDirect Link - Blum, K., T.J.H. Chen, B.W. Downs, B. Meshkin and S.H. Blum et al., 2007. Synaptamine (SG8839),TM an amino-acid enkephalinase inhibition nutraceutical improves recovery of alcoholics, a subtype of Reward Deficiency Syndrome (RDS). Trends Applied Sci. Res., 2: 132-138.
CrossRefDirect Link - Boyle, M.P., B.J. Kolber, S.K. Vogt, D.F. Wozniak and L.J. Muglia, 2006. Forebrain glucocorticoid receptors modulate anxiety-associated locomotors activation and adrenal responsiveness. J. Neurosci., 26: 1971-1978.
CrossRefDirect Link - Braga, M.F.M., V. Aroniadou-Anderjaska, R.M. Post and H. Li, 2002. Lamotrigine reduces spontaneous and evoked GABAA receptor-mediated synaptic transmission in the basolateral amygdala: Implications for its effects in seizure and affective disorders: Lamotrigine and inhibition in the amygdala. Neuropharmacology, 42: 522-529.
CrossRefDirect Link - Braga, M.F.M., V. Aroniadou-Anderjaska, S.T. Manion, C.J. Hough and H. Li, 2004. Stress impairs α1A adrenoceptor-mediated noradrenergic facilitation of GABAergic transmission in the basolateral amygdala. J. Neuropsychopharmacol., 29: 45-58.
CrossRef - Brownstein, M., J.M. Saavedra and J. Axelrod, 1973. Control of pineal N-acetylserotonin by a beta adrenergic receptor. Mol. Pharmacol., 9: 605-611.
PubMedDirect Link - Cecchi, M., N. Capriles, S.J. Watson and H. Akil, 2007. β1 adrenergic receptors in the bed nucleus of stria terminalis mediate differential responses to opiate withdrawal. Neuropsychopharmacology, 32: 589-599.
CrossRefDirect Link - Christiansen, K. and R. Knussmann, 1987. Androgen levels and components of aggressive behavior in men. Horm. Behav., 21: 170-180.
CrossRefDirect Link - Clark, A.S. and L.P. Henderson, 2003. Behavioral and physiological responses to anabolic-androgenic steroids. Neurosci. Biobehav. Rev., 27: 413-436.
CrossRefDirect Link - Clayton, E.C. and C.L. Williams, 2000. Adrenergic activation of the nucleus tractus solitarius potentiates amygdala norepinephrine release and enhances retention performance in emotionally arousing and spatial tasks. J. Behav. Brain Res., 112: 151-158.
PubMedDirect Link - Cunningham, G.R., D.J. Tindall and A.R. Means, 1979. Differences in steroid specificity for rat androgen binding protein and the cytoplasmic receptor. Steroids, 33: 261-276.
CrossRefDirect Link - Dunn, R.W., R. Corbett and S. Fielding, 1989. Effects of 5-HT1A receptor agonists and NMDA receptor antagonists in the social interaction test and the elevated plus maze. Eur. J. Pharmacol., 169: 1-10.
CrossRefDirect Link - ElAttar, T., W. Dirscherl and K.O. Mosebach, 1964. The initial metabolism of 4-14C-testosterone in rat III. The sequence of appearance of testosterone metabolites in the liver. Acta Endocrinol., 45: 527-534.
PubMedDirect Link - Fernandez-Guasti, A. and L. Martinez-Mota, 2005. Anxiolytic-like actions of testosterone in the burying behavior test: Role of androgen and GABA-benzodiazepine receptors. Psychoneuroendocrinology, 30: 762-770.
Direct Link - Forman, L.J., V. Tingle, S. Estilow and J. Cater, 1989. The response to analgesia testing is affected by gonadal steroids in the rat. Life Sci., 45: 447-454.
CrossRefDirect Link - Frye, C.A. and E.H. Lacey, 2001. Posttraining androgens` enhancement of cognitive performance is temporally distinct from androgens` increases in affective behavior. Cogn. Affect. Behav. Neurosci., 1: 172-182.
PubMedDirect Link - Frye, C.A. and T.A. Reed, 1998. Androgenic neurosteroids: Anti-seizure effects in an animal model of epilepsy. Psychoneuroendocrinology, 23: 385-399.
PubMedDirect Link - Frye, C.A., A. Babson and A.A. Walf, 2007. Self-administration of 3α-androstanediol increases locomotion and analgesia and decreases aggressive behavior of male hamsters. Pharmacol. Biochem. Behav., 86: 415-421.
PubMedDirect Link - Frye, C.A., C.M. McCormick, C. Coopersmith and M.S. Erskine, 1996. Effects of paced and non-paced mating stimulation on plasma progesterone, 3 α-diol and corticosterone. Psychoneuroendocrinology, 21: 431-439.
PubMedDirect Link - Frye, C.A., M.E. Rhodes, A. Walf and J.P. Harney, 2002. Testosterone enhances aggression of wild-type mice but not those deficient in type I α-reductase. Brain Res., 948: 165-170.
PubMedDirect Link - Fu, A., X. Li and B. Zhao, 2008. Role of β1-adrenoceptor in the basolateral amygdala of rats with anxiety-like behavior. Brain Res., 1211: 85-92.
PubMedDirect Link - Gee, K.W., 1988. Steroid modulation of the GABA/benzodiazepine receptor-linked chloride ionophore. Mol. Neurobiol., 2: 291-317.
CrossRefDirect Link - Killcross, S., T.W. Robbins and B.J. Everitt, 1997. Different types of fear-conditioned behaviour mediated by separate nuclei within amygdala. Nature, 388: 377-380.
Direct Link - Kim, J.J., J.W. Koo, H.J. Lee and H.S. Han, 2005. Amygdalar inactivation blocks stress-induced impairments in hippocampal long-term potentiation and spatial memory. The J. Neurosci., 25: 1532-1539.
Direct Link - Lee, S.L. and B.L. Fanburg, 1991. Beta adrenergic antagonists inhibit serotonin uptake by pulmonary vascular cells in culture. J. Pharmacol. Exp. Ther., 257: 895-900.
Direct Link - Matuszewich, L., J.J. Karney, S.R. Carter, S.P. Janasik, J.L. O`Brien and R.D. Friedman, 2007. The delayed effects of chronic unpredictable stress on anxiety measures. Physiol. Behav., 90: 674-681.
PubMedDirect Link - Minor, T.R. and A.M. Hunter, 2002. Stressor controllability and learned helplessness research in the United States: Sensitization and fatigue processes. Integr. Physiol. Behav. Sci., 37: 44-58.
CrossRefDirect Link - Molina, N., M.T. Bedran-de-Castro and J.C. Bedran-de-Castro, 1994. Sex-related differences in the analgesic response to the rat tail immersion test. Braz. J. Med. Biol. Res., 27: 1669-1672.
PubMedDirect Link - Monjo, M., A.M. Rodriguez, A. Palou and P. Roca, 2003. Direct effects of testosterone, 17 beta-estradiol and progesterone on adrenergic regulation in cultured brown adipocytes: Potential mechanism for gender-dependent thermogenesis. Endocrinology, 144: 4923-4930.
PubMedDirect Link - Naghibi, B., V. Sheibani, M. Bagherinia, G. Dehghan-Nudeh and F. Sharififar, 2011. Anti anxiety effect of ghavoot: A traditional nutrient preparation. Int. J. Biol. Chem., 5: 322-326.
CrossRefDirect Link - Ordway, G.A., C. Gambarana, S.M. Tejani-Butt, P. Areso, M. Hauptmann and A. Frazer, 1991. Preferential reduction of binding of 125I-iodopindolol to beta-1 adrenoceptors in the amygdala of rat after antidepressant treatments. J. Pharmacol. Exp. Ther., 257: 681-690.
PubMedDirect Link - Pandey, S.G., X. Ren, J. Sagen and G.N. Pandey, 1995. β-adrenergic receptor subtypes in stress-induced behavioral depression. Pharmacol. Biochem. Behav., 51: 339-344.
CrossRefDirect Link - Pellow, S. and S.E. File, 1986. Anxiolytic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: A novel test of anxiety in the rat. Pharmacol. Biochem. Behav., 24: 525-529.
CrossRefDirect Link - Quirarte, G.L., B. Roozendaal and J.L. McGaugh, 1997. Glucocorticoid enhancement of memory storage involves noradrenergic activation in the basolateral amygdala. Proc. Natl. Acad. Sci. USA, 94: 14048-14053.
CrossRefDirect Link - Rodriguez Manzanares, P.A., N.A. Isoardi, H.F. Carrer and V.A. Molina, 2005. Previous stress facilitates fear memory, attenuates GABAergic inhibition and increases synaptic plasticity in the rat basolateral amygdala. J. Neurosci., 25: 8725-8734.
CrossRefDirect Link - Roozendaal, B., G.K. Hui, I.R. Hui, D.J. Berlau, J.L. McGaugh and N.M. Weinberger, 2006. Basolateral amygdala noradrenergic activity mediates corticosterone-induced enhancement of auditory fear conditioning. Neurobiol. Learn. Memory, 86: 249-255.
CrossRefDirect Link - Roselli, C.E., 1991. Sex differences in androgen receptors and aromatase activity in microdissected regions of the rat brain. Endocrinology, 128: 1310-1316.
PubMedDirect Link - Rosen, J.B. and J. Schulkin, 1998. From normal fear to pathological anxiety. Psychol. Rev., 105: 325-350.
PubMedDirect Link - Royer, S., M. Martina and D. Pare, 1999. An inhibitory interface gates impulse traffic between the input and output stations of the amygdala. J. Neurosci., 19: 10575-10583.
PubMedDirect Link - Rudoy, C.A. and E.J. van Bockstaele, 2007. Betaxolol, a selective β1-adrenergic receptor antagonist, diminishes anxiety-like behavior during early withdrawal from chronic cocaine administration in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry, 31: 1119-1129.
CrossRefDirect Link - Sadeghi, Z., M. Eidi, A. Eidi and M. Bahar, 2007. Effects of practical examination stress on serum ACTH and cortisol concentrations in normal and anxious students. J. Med. Sci., 7: 489-491.
CrossRefDirect Link - Samad, N., T. Parveen, S. Haider and D.J. Haleem, 2005. Attenuation of restraint-induced anorexia and anxiogenic behavior by serotonin-1A agonists in rats. J. Med. Sci., 5: 289-293.
CrossRefDirect Link - Heldt, S.A. and K.J. Ressler, 2006. Localized injections of midazolam into the amygdala and hippocampus induce differential changes in anxiolytic-like motor activity in mice. Behav. Pharmacol., 17: 349-356.
Direct Link - Skurlova, M., A. Stofkova and J. Jurcovicov, 2011. Anxiety-like behavior in the elevated-plus maze tests and enhanced IL-1 β, IL-6, NADPH oxidase-1 and iNOS mRNAs in the hippocampus during early stage of adjuvant arthritis in rats. Neurosci. Lett., 487: 250-254.
PubMedDirect Link - Mahbub-E-Sobhani, N. Haque, U. Salma and A. Ahmed, 2011. Immune modulation in response to stress and relaxation. Pak. J. Biol. Sci., 14: 363-374.
CrossRef - Southwick, S.M., S. Paige, C.A. Morgan, J.D. Bremner, J.H. Krystal and D.S. Charney, 1999. Neurotransmitter alterations in PTSD: Catecholamines and serotonin. Semin Clin. Neuropsychiatry, 4: 242-248.
PubMedDirect Link - Sullivan, R.M., A. Duchesne, D. Hussain, J. Waldron and F. Laplante, 2009. Effects of unilateral amygdala dopamine depletion on behaviour in the elevated plus maze: Role of sex, hemisphere and retesting. Behav. Brain Res., 205: 115-122.
CrossRefDirect Link - Svensson, A.I., B. Soderpalm and J.A. Engel, 2000. Gonadectomy enhances shock-induced behavioral inhibition in adult male rats: Implications for impulsive behavior. Pharmacol. Biochem. Behav., 65: 731-736.
CrossRefDirect Link - Szinyei, C., T. Heinbockel, J. Montagne and H.C. Pape, 2000. Putative cortical and thalamic inputs elicit convergent excitation in a population of GABAergic interneurons of the lateral amygdala. J. Neurosci., 20: 8909-8915.
PubMedDirect Link - Taiwe, G.S., E.N. Bum, T. Dimo, E. Talla and N. Weiss et al., 2010. Antidepressant, myorelaxant and anti-anxiety-like effects of Nauclea latifolia smith (Rubiaceae) roots extract in murine models. Int. J. Pharmacol., 6: 364-371.
CrossRefDirect Link - Van der Wee, N.J., J.F. van Veen, H. Stevens, I.M. van Vliet, P.P. van Rijk and H.G. Westenberg, 2008. Increased serotonin and dopamine transporter binding in psychotropic medication-naive patients with generalized social anxiety disorder shown by 123I-β-(4-iodophenyl)-tropane SPECT. J. Nucl. Med., 49: 757-763.
CrossRefPubMedDirect Link - Van Nobelen, M. and L. Kokkinidis, 2006. Amygdaloid GABA, not glutamate neurotransmission or mRNA transcription controls footshock-associated fear arousal in the acoustic startle paradigm. Neuroscience, 137: 707-716.
CrossRefDirect Link - Velisek, L., 2006. Prenatal exposure to betamethasone decreases anxiety in developing rats: Hippocampal neuropeptide y as a target molecule. Neuropsychopharmacology, 31: 2140-2149.
PubMedDirect Link - Raj, K.K.V., B. Narayana, N.S. Kumari, B.V. Ashalatha and B.K. Sarojini, 2006. Neuropsychobehavioral effects and anticancer activity of some substituted triazolo[4,3-a][1,4] benzodiazepines. J. Pharmacol. Toxicol., 1: 471-477.
CrossRefDirect Link - Walf, A.A. and C.A. Frye, 2007. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protocols, 2: 322-328.
CrossRefDirect Link - Woode, E., E. Boakye-Gyasi, G.K. Ainooson, C. Ansah and M. Duwiejua, 2009. Anti-nociceptive effects and the mechanism of Palisota hirsuta K. Schum. Leaf extract in murine models. Int. J. Pharmacol., 5: 101-113.
CrossRefDirect Link - Woode, E., E. Boakye-Gyasi, N. Amidu, C. Ansah and M. Duwiejua, 2010. Anxiolytic and antidepressant effects of a leaf extract of Palisota hirsute K. schum. (Commelinaceae) in mice. Int. J. Pharmacol., 6: 1-17.
CrossRefDirect Link - Yeh, H.H. and D.J. Woodward, 1983. Beta-1 adrenergic receptors mediate noradrenergic facilitation of Purkinje cell responses to gamma-aminobutyric acid in cerebellum of rat. Neuropharmacology, 22: 629-639.
Direct Link - Zuloaga, D.G., C.L. Jordan and S.M. Breedlove, 2011. The organizational role of testicular hormones and the androgen receptor in anxiety-related behaviors and sensorimotor gating in rats. Endocrinology, 152: 1572-1581.
PubMedDirect Link - Wang, D.V., F. Wang, J. Liu, L. Zhang, Z. Wang and L. Lin, 2011. Neurons in the amygdala with response-selectivity for anxiety in two ethologically based tests. PLoS One, Vol. 6.
CrossRef - Edinger, K.L. and C.A. Frye, 2006. Intrahippocampal administration of an androgen receptor antagonist, flutamide, can increase anxiety-like behaviour in intact and DHT-replaced male rats. Horm. Behav., 50: 216-222.
CrossRefDirect Link