
P. Ratanachamnong
Department of Pharmacology, Faculty of Science, Mahidol University, Bangkok, 10400, Thailand
U. Matsathit
Department of Pharmacology, Faculty of Medicine, Srinakharinwirot University, Bangkok, 10110, Thailand
Y. Sanvarinda
Department of Pharmacology, Faculty of Science, Mahidol University, Bangkok, 10400, Thailand
P. Piyachaturawat
Department of Physiology, Faculty of Science, Mahidol University, Bangkok, 10400, Thailand
L. Phivthong-Ngam
Department of Pharmacology, Faculty of Medicine, Srinakharinwirot University, Bangkok, 10110, Thailand
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 4 | Page No.: 234-242







ABSTRACT
Curcuma comosa Roxb. (C. comosa), an indigenous medicinal plant containing phytoestrogens, has been reported to have hypocholesterolemic and anti-inflammatory effects. Hypercholesterolemia and inflammation play crucial roles in the development of atherosclerosis. In the present study we investigated whether the hexane extract of C. comosa inhibits atherosclerotic plaque formation and platelet aggregation in hypercholesterolemic rabbits. Rabbits were fed 0.5% cholesterol (N = 6), 0.5% cholesterol plus 5 mg day-1 of simvastatin (N = 6), 0.5% cholesterol plus 100 mg kg-1 b.wt./day of C. Comosa extract (N = 6) or normal rabbit chow (N = 6) for 12 weeks. Blood samples for cholesterol determination were collected in monthly intervals. At the end of the study period, platelet aggregation ex vivo, endothelium-dependent and -independent vascular function of isolated aortic rings ex vivo and aortic plaques were assessed. Cholesterol feeding to the rabbits for 12 weeks significantly increased plasma lipid profiles, platelet aggregation and impaired endothelium-dependent relaxations to ACh and ADP. Supplement with simvastatin or hexane extract of C. comosa attenuated platelet aggregation, plasma lipid profiles and partly restored endothelium-mediated vascular responses. Both simvastatin and hexane extract of C. comosa also improved aortic intimal thickening by about 76 and 74%, respectively. The results indicate that hexane extract of C. comosa possesses the beneficial effects in hypercholesterolemia in similar manner to simvastatin. The therapeutic potential for the prevention or regression of atherosclerosis should be further investigated.
PDF Abstract XML References Citation
Received: November 04, 2011;
Accepted: February 16, 2012;
Published: March 27, 2012
How to cite this article
P. Ratanachamnong, U. Matsathit, Y. Sanvarinda, P. Piyachaturawat and L. Phivthong-Ngam, 2012. Effects of Curcurma comosa Roxb. on Platelet Aggregation and Atherosclerotic Plaque Development in Hypercholesterolemic Rabbits. International Journal of Pharmacology, 8: 234-242.
DOI: 10.3923/ijp.2012.234.242
URL: https://scialert.net/abstract/?doi=ijp.2012.234.242
DOI: 10.3923/ijp.2012.234.242
URL: https://scialert.net/abstract/?doi=ijp.2012.234.242
INTRODUCTION
Platelets play a key role in homeostasis of the vascular wall. They can also contribute to endothelial dysfunction, modulate various inflammatory responses and also initiate atheromatous plaque formation. Atherosclerotic plaque development involves a complex interaction between the endothelium, inflammatory cytokines and numerous blood elements (Garelnabi, 2010). At the sites of injured endothelium, platelets form a primary platelet plug, followed by the expression or secretion of various molecules that assist in vessel repair in the stages of platelet adhesion, activation and aggregation. Adhering platelets recruit monocytes that transmigrate into the subendothelial space and develop into macrophages and foam cells.
In addition, platelets are a source of inflammatory mediators and growth factors, including adhesion proteins (e.g., fibrinogen, fibronectin, von Willebrand factor, thrombospondin, vitronectin, P-selectin), growth factors (e.g., platelet-derived growth factor, transforming growth factor-β, epidermal growth factor, basic fibroblast growth factor), chemokines (e.g., PF-4, regulated on activation, normal T-cell expressed and secreted, epithelial neutrophil-activating protein 78, macrophage inflammatory protein-1α), cytokines and cytokine-like factors (e.g., interleukin-1β, CD40 ligand, thromboglobulin-β) and coagulation factors (e.g., factor V, factor XI, plasminogen activator inhibitor type 1, plasminogen, protein S) (Jennings, 2009). These chemokines and growth factors are detectable in atherosclerotic plaques (Pitsilos et al., 2003). Moreover, it was reported that activation of circulating platelets correlates with enhanced wall thickness of the carotid artery in humans (Saeed et al., 2004; Fateh-Moghadam et al., 2005), platelet aggregability are increased in animal models of hypercholesterolemia and in human subjects with familial hypercholesterolemia. Activated platelets also modulate the tone and the structure of the vascular wall (Bode-Boger et al., 1998). In hypercholesteremic rabbits, platelets adhere to the endothelium of atherosclerotic sites in the absence of endothelial cell denudation before manifest atherosclerotic lesions develop (Theilmeier et al., 2002). Accumulating evidence suggests that inhibition of platelet function can modulate inflammatory markers, such as CD40 ligand, P-selectin and C-reactive protein and reduce vascular inflammation (Rosenson and Tangney, 1998; Hermann et al., 2001; Saeed et al., 2004). The report from the study of the antiplatelet Trialists’ collaboration meta-analysis showed the benefit of antiplatelet therapy to patients with atherosclerotic disease (Antithrombotic Trialists’ Collaboration, 2002). Antiplatelet drugs, such as aspirin or clopidogrel, have been demonstrated to reduce the incidence of Myocardial Infarction (MI), stroke or vascular death in patients with vascular disease (CAPRIE Steering Committee, 1996; Rosenson and Tangney, 1998; Saeed et al., 2004). However, bleeding risk is a substantial limitation of antiplatelet therapy, because these agents target platelet activation pathways critical for both protective haemostasis and pathologic thrombosis.
Curcuma comosa Roxb. (Zingiberaceae) has been used extensively as indigenous Thai medicinal plant. Its rhizome was found to contain an estrogenic like action and has been traditionally used as an anti-inflammatory agent in postpartum uterine bleeding and uterine inflammation. It has been reported that the extracts of C. comosa possess a hypocholesterolemic (Piyachaturawat et al., 1999), an anti-oxidative effects (Niumsakul et al., 2007). Both hexane and ethanol extract of C. comosa exhibit an anti-inflammatory action by suppression the release of pro-inflammatory cytokines, tumor necrosis factor α (TNF-α) and interleukin-1β, from phorbol-12-myristate-13-acetate (PMA)-stimulated peripheral blood mononuclear cells (PBMC) and human pro-monocytic cell line (U937) (Sodsai et al., 2007). Recently, the hexane extract of C. comosa and its isolated compound, a diaryltheptanoid-D3, have been demonstrated to have vasodilator effect as shown by enhancement of endothelium-dependent vascular relaxation of rat aortic rings (Intapad et al., 2009). Therefore, it was interesting to determine the role of C. comosa on the platelet activity and atherosclerotic plaque development. In the present study, we investigated whether the hexane extract of C. comosa could attenuate platelet aggregation, restore endothelium function and prevent atherosclerotic plaque formation in rabbit atherosclerosis model.
MATERIALS AND METHODS
Materials: C. comosa powder was kindly provided and characterized by Professor Apichart Suksamrarn, Faculty of Science, Ramkamhaeng University. Its main constituents were 1,7-diphenyl-(6E)-6-hepten-3-ol and 1,7-diphenyl-(4E,6E)-4,6-heptadien-3-ol. In brief, dried rhizome of C. comosa was collected from Nakornpathom province. A voucher specimen (BKF No. 97298) was deposited at the Forest Herbarium, Royal Forest Department, Ministry of Agriculture and Cooperatives, Bangkok. The C. comosa rhizomes were sliced and dried at 50-60°C. The dried rhizomes were pulverized before used in animals. Simvastatin was purchased from an accredited drug store (Bangkok, Thailand). Normal diet chow and enriched with 0.5% cholesterol were prepared by Charoen Pokphand Foods Public Company Limited, Thailand.
Animals and study design: Twenty-four male New Zealand White rabbits initially weighing 1.5-2 kg were supplied by the National Laboratory Animal Center of Thailand, Mahidol University, Salaya Campus, Nakornpathom, Thailand. The study protocol was approved by the ethical committee for studies in animals of Srinakharinwirot University, which conformed with the International Guiding Principles for Biomedical Research Involving Animals (CIOMS/WHO). The animals were housed with one animal per cage at the Faculty of Medicine, Srinakharinwirot University, Thailand. All of the animals were kept in a humidity-controlled room at a constant temperature of 25±2°C and maintained on a 12 h alternate light-dark cycle, with free access to food (C.P. Company, Thailand) and drinking water ad libitum throughout the experiment.
After 2 weeks of adaptation, the rabbits were randomly divided into four groups of six animals each. Rabbits in the first group served as a control group (control group) were fed normal rabbit chow throughout the study. The other three groups of rabbits were fed a diet containing 0.5% cholesterol (cholesterol group), or 0.5% cholesterol plus 5 mg day-1 of simvastatin (cholesterol+simvastatin group), or 0.5% cholesterol plus 100 mg kg-1 b.wt./day of hexane extract of C. comosa (cholesterol+C. comosa group) for 12 weeks. Food and water were allowed ad libitum. Body weight, food and water consumption were measured at weekly intervals.
Before the start of the dietary intervention and every 4 weeks thereafter, blood samples were obtained from the central ear artery. Plasma was removed after centrifugation (1,500 g, 10 min) and stored at -20°C before measuring plasma cholesterol concentrations. At the end of the experimental period, the rabbits were killed. The aorta was isolated for the measurement of vascular function and histological examination.
Measurement of plasma cholesterol: Plasma total cholesterol concentrations were quantified by commercially available enzymatic methods (Hoffmann-La Roche Inc. USA).
Platelet aggregation: Platelet Rich Plasma (PRP) was generated from citrated blood by centrifugation at 80 g for 10 min at 25°C. Platelet Poor Plasma (PPP) was prepared from the remaining volume of blood by centrifugation at 2,000 g for 15 min at 25°C.
Platelet aggregation was performed spectrophotometrically on Chronolog aggregometer (500 CA/560CA) as described previously (Bode-Boger et al., 1998). The aggregometer was adjusted before each test so that in each subject the value for light transmission for PRP was 0% and that for PPP was 100%. Aggregation was induced in duplicate using four different concentrations of adenosine diphosphate (ADP; final concentrations of 0.5, 1.0, 2.0 and 4.0 mmol L-1) and was monitored for 4 min. Aggregations were evaluated as area under the aggregation curves (Born, 1962).
Measurement of vascular functions: The aortas were dissected free of adhering fat and connective tissue. The specimens were cut into rings of approximately 3 mm long, four rings per animal. During harvesting, special care was taken to avoid contact with the luminal surface of the rings to preserve the endothelium. The aortic rings were immediately placed into organ baths of 20 mL capacity containing modified Krebs solution (pH 7.4) of the following composition (in mM): Na+ 145.0, K+ 5.95, Ca2+ 1.7, Mg2+ 1.2, Cl¯ 128.15, HCO3¯25.0, H2PO42¯ 1.2, SO42¯ 1.2, glucose 10.6, EDTA 0.025. The buffer was maintained at 37°C and continuously aerated with a gas mixture consisting of 95% O2: 5% CO2. The aortic rings were connected to force transducers (Biegestab SB-24, Japan) for the measurement of isometric tension. Each aortic ring was gradually stretched to a resting tension of 2 g and allowed to equilibrate for 60 min. The rings were then contracted with 1 μM Norepinephrine (NE) and relaxed by 1 μM Acetylcholine (ACh) for testing of endothelial integrity. Thereafter, the vascular rings were repeatedly washed with Krebs buffer until the tension had returned to the previous baseline value. Aortic rings from control animals always exhibited more than 70% relaxation of the NE-induced contraction plateau in response to 1 μM ACh.
Subsequently, cumulative concentration response curves were obtained with the endothelium-dependent relaxant ACh or Adenosine Diphosphate (ADP) and the endothelium-independent relaxant Sodium Nitroprusside (SNP) after precontraction with 1 μM norepinephrine (all drugs concentration ranging from 1 nM to 0.1 mM). Aortic relaxations were expressed as percentage of the precontractile tension induced with 1 μM NE.
Aortic atherosclerotic plaque formation: Segments of the thoracic aorta were fixed in formalin, embedded in paraffin and stained with haematoxylin/eosin for the morphometric measurement of intimal and medial cross-sectional areas by using a planimeter (semi-automatic mode). Four sections of each animal were analyzed and the values were averaged. The observers performing the histomorphological measurements were blinded to the treatment groups.
Statistical analysis: All data were expressed as Mean± SEM. Statistical significances between groups and within a group at multiple time points were analyzed using multiple comparison analysis of variance followed by the Scheffe’ F-test (Fisher and van Belle, 1993). Statistical significance was accepted at the 0.05 level of probability.
RESULTS
Body weight and plasma cholesterol concentrations: The body weights of the rabbits in all experimental groups were regularly increased with no significant differences between the groups at all periods (Table 1). Plasma total cholesterol, LDL-C and HDL-C concentrations were elevated by cholesterol-enriched diet. Supplementation with either simvastatin or C. comosa significantly decreased plasma total cholesterol after the 4-week experimental period and LDL-C after the 8-week experimental period, compared with the control group. There were no significant differences in HDL-C concentrations at any time point between the rabbits fed cholesterol diet and those supplemented with C. comosa. Treatment with simvastatin significantly lowered HDL-C concentrations only at the end of experiments, compared to cholesterol group.
Platelet aggregation: Platelet aggregation in response to ADP was significantly increased in hypercholesterolemic rabbits as compared to controls over the whole concentration range of ADP (p<0.05; Fig. 1). Supplementation with C. comosa inhibited the increased of percent light transmission of platelet aggregation induced by low concentration of ADP (0.5 and 1 μM). In response to higher concentration of ADP (2 and 4 μM), percentage of platelet aggregation was higher than those in control group but still was significantly lower than in hypercholesterolemic rabbits.
| Table 1: | Body weight, plasma total cholesterol, low-density lipoprotein (LDL)-cholesterol and high-density lipoprotein (HDL)-cholesterol concentrations in the experimental groups |
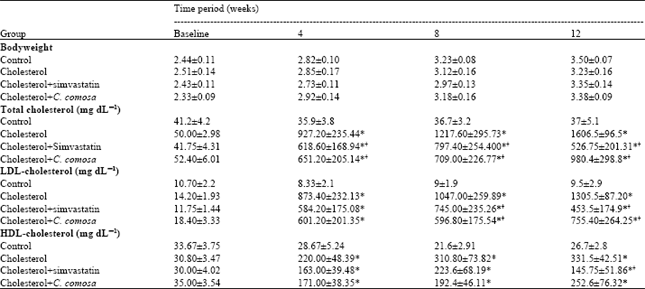 | |
| Data are Mean±SEM, *p<0.05 vs. control, †p<0.05 vs. cholesterol | |
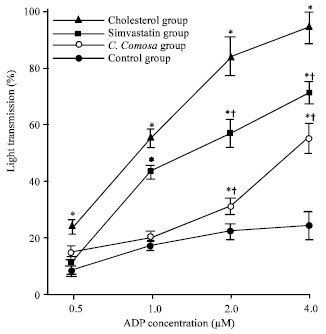 | |
| Fig. 1: | Percent light transmission in platelet aggregation induced by ADP (0.5-4 μM) in platelet rich plasma ex vivo from rabbits in various groups after 12 weeks of the experiments. Data are Mean±SEM, *p<0.05 vs. control, †p<0.05 vs. cholesterol |
Whereas treatment with simvastatin reduced platelet aggregation induced by only higher concentration of ADP (2 and 4 μM).
Endothelial function: Endothelium-dependent relaxations of isolated aortic rings ex vivo in response to Acetylcholine (ACh) and Adenosine Diphosphate (ADP) were significantly attenuated in the cholesterol-fed rabbits as compared to controls (Fig. 2a and b).
| Table 2: | Median effective concentrations (log EC50 values) of relaxation responses of isolated rabbit aortic rings |
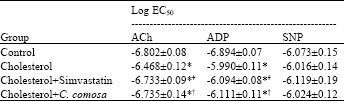 | |
| Data are Mean±SEM, *p<0.05 vs. control, †p<0.05 vs. cholesterol | |
Supplementation with simvastatin or C. comosa partly but not completely restored relaxations to ACh and ADP (p<0.05 vs. cholesterol). In contrast, no significant differences were observed in endothelium-independent relaxations in response to Sodium Nitroprusside (SNP) between all three groups (Fig. 2c).
The EC50 values for the endothelium-dependent and -independent vasodilators are given in Table 2. Sensitivity of isolated aortic rings to ACh and ADP was significantly decreased in cholesterol group as compared to control group (p<0.05) and slightly increased by the supplementation with either simvastatin or C. comosa.
Aortic plaque formation: After 12 weeks of dietary intervention, no intimal thickening of thoracic aorta was observed in the control group. Intima/media ratio in histological cross sections of the thoracic aorta was increased to 1.15±0.18 in the cholesterol group. Both simvastatin and C. comosa administration significantly reduced intimal thickening in the thoracic aorta.
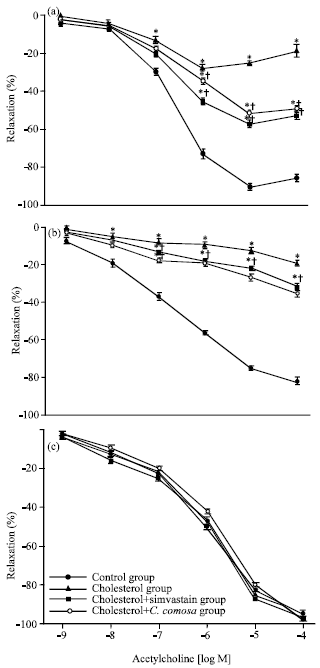 | |
| Fig. 2(a-c): | Endothelium-dependent relaxations induced by (a) ACh, (b) ADP and (c) endothelium-independent relaxations induced by SNP of isolated aortic rings ex vivo from rabbits fed normal rabbit chow, 0.5% cholesterol-enriched diet, 0.5% cholesterol-enriched diet plus 5 mg/day of simvastatin, or plus 100 mg kg-1 b.wt./day of C. comosa (ο) for 12 weeks. Data are Mean±SEM, *p<0.05 vs. control, †p<0.05 vs. cholesterol |
The intima/media ratios were 0.28±0.10 and 0.31±0.07 for the cholesterol plus simvastatin and that plus C. comosa, respectively (Fig. 3).
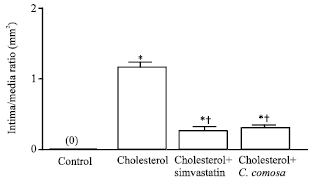 | |
| Fig. 3: | Aortic cross-sectional intima/media ratio in rabbits fed normal rabbit chow, 0.5% cholesterol-enriched diet, or the same diet plus 5 mg day-1 of simvastatin, or plus 100 mg kg-1 b.wt./day of C. comosa for 12 weeks. Data are Mean±SEM, *p<0.05 vs. control, †p<0.05 vs. cholesterol |
DISCUSSION
Our present study suggests that the development of atherosclerotic plaque in cholesterol-fed rabbits could be reduced by daily supplementation with 100 mg kg-1 b.wt./day of the hexane extract of C. comosa or 5 mg day-1 of simvastatin. This beneficial effect is paralleled by (1) decreased plasma concentration of total cholesterol and LDL-C, (2) reduced platelet aggregation and (3) improvement of vascular function.
Atherosclerosis is associated with increased total cholesterol and LDL-C, impaired endothelium-dependent vasodilation, enhanced platelet aggregation, increased oxidative stress and endothelial adhesiveness for monocytes (Jennings, 2009; Garelnabi, 2010). Rabbit fed a high-cholesterol is well known for a good model to study hypercholesterolemia and atherosclerosis (Phivthong-Ngam et al., 1998; Srisawat et al., 2003; Abdelhalim and Alhadlaq, 2008; Chumark et al., 2008). Dietary containing high cholesterol (0.5-1%) increases the levels of plasma total cholesterol and LDL-C in rabbits, concomitantly with increased platelet aggregation, impaired vascular function and aortic atherosclerotic plaque formation (Bode-Boger et al., 1998; Phivthong-Ngam et al., 1998; Srisawat et al., 2003; Chumark et al., 2008). The similar changes were found in this study.
Simvastatin was used as the positive control. It is a hydroxy-3-methylglutaryl coenzyme A reductase inhibitor, a potent cholesterol lowering drug. Cholesterol-lowering drugs are therapeutic options that have been shown to reduce the biological and clinical burden related to atherosclerosis (Rosenson and Tangney, 1998; Anand et al., 2008; Superko and King, 2008). The effectiveness of statins in the prevention of coronary artery disease is not only to reduce cholesterol levels but also to stabilize atherosclerotic plaque, improve paraoxonase activity, improve endothelial function, enhance fibrinolysis and antithrombotic effects (Rosenson and Tangney, 1998; Undas et al., 2001; Shetty et al., 2008). Previous studies reported that statin therapy could upregulate nitric oxide production or activity and reduce oxidative stress (Anand et al., 2008; Superko and King, 2008). The dose of simvastatin used in our study (5 mg day-1) was a low dose that has been reported to reduce plasma total cholesterol in rabbits without toxicity (Alfon et al., 1999). Consistent with these studies, our results showed that simvastatin treatment reduced plasma cholesterol, prevented the platelet aggregation, restored impaired endothelium-dependent vasodilation and attenuated plaque formation in cholesterol-fed rabbits. Antiplatelet activity of simvastatin has been previously demonstrated both in hypercholesterolemic patients and animals (Rosenson and Tangney, 1998). Lee and co-workers recently reported that the antiplatelet activity of simvastatin involved an increase of the cyclic AMP-eNOS/NO-cyclic GMP pathway, followed by inhibition of the PLCγ2-PKC-p38 MAPK-TxA2 cascade (Lee et al., 2010).
C. comosa and its active chemical compound, phloroacetophenone or 2, 4, 6 trihydroxyacetophenone, have been reported to decrease plasma cholesterol. Intragastric administration of C. comosa rhizome ethyl acetate extract to hypercholesterolemic hamsters at a dose of 500 mg/kg/day for 7 day caused decreased both plasma triglyceride and cholesterol levels (Piyachaturawat et al., 1999). Phloroacetophenone also decreased plasma cholesterol in rats (Piyachaturawat et al., 2002a) and decreased both plasma cholesterol and triglyceride level in hypercholesterolemic hamsters (Piyachaturawat et al., 2002b). Hypocholesterolemic effect of C. comosa was also found in our experiments. The dose of C. comosa used in this study (100 mg kg-1 b.wt./day) was much lower than the maximum dose for hypercholesterolemic hamsters as reported by Piyachaturawat et al. (1999) and was not shown to have any toxicity. Phloroacetophenone, an active compound of C. comosa, was found to enhance the activity of hepatic cholesterol-7-α-hydroxylase, the rate-limiting step enzyme for hepatic conversion of cholesterol to bile acid and excreted via biliary secretion (Charoenteeraboon et al., 2005). Lowering effect of C. comosa in this study may be due to an activation of hepatic cholesterol-7-α-hydroxylase enzyme. However, the accurate mechanism of C. comosa on lipid lowering activity should be further investigated.
Platelet aggregability is enhanced in hypercholesterolemic animals and patients (Jennings, 2009). There are many mechanisms considered for the enhanced platelet aggregation. One of them is an increase in the adenosine diphosphate-induced fibrinogen binding to platelets mediated by LDL in a dose-dependent manner (Andrews et al., 1987). LDL (Van Willigen et al., 1994) and oxidized LDL (Relou et al., 2003) have been shown to activate platelets through a protein kinase C (PKC)-dependent mechanism. In addition, oxidative stress which is associated with the hypercholesterolemia (Ferroni et al., 2004; Prasanna and Purnima, 2011) has also been reported to activate PKC (Kooy et al., 1993). Thus, simvastatins and C. comosa may inhibit platelet aggregation by lowering LDL levels in vivo. On the other hand, they may affect platelet function by changing the cholesterol content of platelet membranes, which alters membrane fluidity (Rosenson and Tangney, 1998). Among these mechanisms, Nitric Oxide (NO) also inhibits platelet aggregation in vivo and in vitro. Hypercholesterolemia impairs platelet-derived nitric oxide (PDNO) released from platelets attributable to increased oxidative stress. Platelets possess the L-arginine-NO pathway through constitutive NO synthase in humans (Sase and Michel, 1995). PDNO during platelet aggregation increases the intraplatelet level of cyclic guanosine 3’, 5’-monophosphate and then inhibits platelet aggregation. Thus, PDNO acts as a negative feedback mechanism to inhibit not only platelet aggregation but also recruitment after aggregation (Freedman et al., 1997). It has been found that PDNO was lower in hypercholesterolemic patients (Wolf et al., 1997).
Phytoestrogens presented in C. comosa hexane extract are phenolic compounds which show a similar activity to estrogen (Piyachaturawat et al., 1999). The effect of estrogen on the coagulation system is variable. Platelets incubated with estradiol manifest decreased adherence (Miller et al., 1994), aggregation (Nakano et al., 1998), fibrinogen (Miller et al., 1995) and plasminogen activator inhibitor (Koh et al., 1999), leading to inhibition of platelet aggregation.
Endothelial dysfunction is an important early event in the pathogenesis of atherosclerosis, contributing to plaque initiation and progression. Impaired responses to endothelium-dependent vasodilators have been shown in arteries and arterioles from cholesterol-fed animal (Phivthong-Ngam et al., 1998; Srisawat et al., 2003) and from human associated with hypercholesterolemia and atherosclerosis (Creager et al., 1992). Several lines of evidence suggest that the vascular dysfunction associated with atherosclerosis is due to increased serum cholesterol levels (Sudano et al., 2006). Cholesterol-lowering drugs or diet can attenuate the accumulation of cholesterol and preserve endothelium-dependent relaxation in hypercholesterolemic animal models and in human. One of the major factors responsible for endothelial dysfunction appears to be a reduced stability of the endothelial enzyme generating NO, endothelial nitric oxide synthase (eNOS) (Lokhande et al., 2006). It was demonstrated that statins prevented hypoxia-induced down regulation of eNOS in human endothelial cells by stabilizing eNOS mRNA, leading to an increase in NO production by these cells (Laufs et al., 1997). Statins may influence plaque composition by inhibition of cholesterol accumulation in monocyte-derived macrophages either by reducing the availability of free cholesterol, or by reduction of the synthesis of mevalonate and its derivates which are required for cholesterol esterification (Bernini et al., 1993).
In persons at high risk of cardiovascular events, a greater isoflavone intake is associated with better vascular endothelial function and lower carotid atherosclerotic burden (Ogbuewu et al., 2010; Nurulhuda et al., 2012). It has been reported that genistein reduced the elevated blood pressure and endothelial dysfunction in Spontaneously Hypertensive Rats (SHRs). This latter effect appears to be related to increased eNOS activity associated with increased calmodulin-1 expression and decreased O2•G generation (Vera et al., 2007). Furthermore, phytoestrogens have the ability to upregulate the expression of endothelial nitric oxide synthase (eNOS) protein in the endothelial cells of the blood vessels and hence increase in the production of Nitric Oxide (NO) (Mendelsohn and Karas, 1999). In addition, it has been reported that the hexane extract of C. comosa enhanced endothelial-dependent relaxation of rat aortic rings in response to ACh via the estrogen receptors/Akt/eNOS signaling pathway (Intapad et al., 2009).
Atherosclerosis is a chronic inflammatory disease. Several inflammatory cytokines and growth factors, including platelet-derived growth factor, transforming growth factor-β, interleukin-1 beta (IL-1β), monocyte chemoattractant protein-1 (MCP-1) and tumor necrosis factor-alpha (TNF-α), are detectable in atherosclerotic plaques (Garelnabi, 2010; Pitsilos et al., 2003). Hexane and ethanol extract of C. comosa were shown to suppress the release of cytokines TNF-α and IL-1β through attenuating IκB kinase (IKKβ) expression and NF-κB activation (Sodsai et al., 2007).
CONCLUSIONS
The results of the present study support the beneficial effects of C. comosa in the prevention of the atherosclerotic plaque development in the same manner as simvastatin. Its antiatherosclerotic effects seem to be due to cholesterol-lowering, antiplatelet activity and restoration of endothelial function. The mechanisms underlie these effects and therapeutic approach should be further investigated.
ACKNOWLEDGMENT
This study was supported by Mahidol University and National Research Council of Thailand.
REFERENCES
- Abdelhalim, M.A.K. and H.A. Alhadlaq, 2008. Effects of cholesterol feeding periods on blood haematology and biochemistry of rabbits. Int. J. Biol. Chem., 2: 49-53.
CrossRefDirect Link - Alfon, J., J.F. Guasch, M. Berrozpe and L. Badimon, 1999. Nitric oxide synthase II (NOS II) gene expression correlates with atherosclerotic intimal thickening. Preventive effects of HMG-CoA reductase inhibitors. Atherosclerosis, 145: 325-331.
Direct Link - Anand, V.A., M. Chenniappan, S. Kalavathy, K. Uma, M.P. Saravanan and S. Kumar, 2008. Redeeming measure of atorvastatin in the risk factors of cardiovascular disease. Int. J. Pharmacol., 4: 305-309.
CrossRefDirect Link - Andrews, H.E., J.W. Aitken, D.G. Hassall, V.O. Skinner and K.R. Bruckdorfer, 1987. Intracellular mechanisms in the activations of human platelets by low-density lipoproteins. Biochem. J., 242: 559-564.
Direct Link - Antithrombotic Trialists Collaboration, 2002. Paper Collaborative meta-analysis of randomised trials of antiplatelet therapy for prevention of death, myocardial infarction and stroke in high risk patients. Br. Med. J., 324: 71-86.
CrossRefPubMedDirect Link - Bernini, F., G. Didoni, G. Bonfadini, S. Bellosta and R. Fumagalli, 1993. Requirement for mevalonate in acetylated LDL induction of cholesterolesterification in macrophages. Atherosclerosis, 104: 19-26.
Direct Link - CAPRIE Steering Committee, 1996. A randomised, blinded, trial of clopidogrel versus aspirin in patients at risk of ischaemic events (CAPRIE). Lancet, 348: 1329-1339.
CrossRef - Charoenteeraboon, J., K. Nithipatikom, W.B. Campbell, P. Piyachaturawat, P. Wilairat and P. Rongnoparut, 2005. Induction of human cholesterol 7 alpha-hydroxylase in HepG2 cells by 2,4,6-trihydroxyacetophenone. Eur. J. Pharmacol., 515: 43-46.
CrossRef - Chumark, P., P. Khunawat, Y. Sanvarinda, S. Phornchirasilp and N.P. Morales et al., 2008. The in vitro and ex vivo antioxidant properties, hypolipidaemic and antiatherosclerotic activities of water extract of Moringa oleifera Lam. leaves. J. Ethnopharmacol., 116: 439-446.
CrossRefPubMedDirect Link - Creager, M.A., S.J. Gallagher, X.J. Girerd, S.M. Coleman, V.J. Dzau, and J.P. Cooke, 1992. L-arginine improves endothelium-dependent vasodilation in hypercholesterolemic humans. J. Clin. Invest., 90: 1248-1253.
CrossRef - Fateh-Moghadam, S., Z. Li, S. Ersel, T. Reuter and P. Htun et al., 2005. Platelet degranulation is associated with progression of intima-media thickness of the common carotid artery in patients with diabetes mellitus type 2. Arterioscler. Thromb. Vasc. Biol., 25: 1299-1303.
CrossRef - Ferroni, P., S. Basili, A. Falco and G. Davi, 2004. Antioxidants and redox signaling. Antioxid Redox Signal, 6: 747-756.
CrossRef - Freedman, J.E., J. Loscalzo, M.R. Barnard, C. Alpert, J.F. Keaney and A.D. Michelson, 1997. Nitric oxide released from activated platelets inhibits platelet recruitment. J. Clin. Invest., 100: 350-356.
Direct Link - Garelnabi, M., 2010. Emerging evidences from the contribution of the traditional and new risk factors to the atherosclerosis pathogenesis. J. Med. Sci., 10: 153-161.
CrossRef - Hermann, A., B.H. Rauch, M. Braun, K. Schror and A.A. Weber, 2001. Platelet CD40 ligand (CD40L)-subcellular localization, regulation of expression, and inhibition by clopidogrel. Platelets, 12: 74-82.
Direct Link - Intapad, S., A. Suksamrarn and P. Piyachaturawat, 2009. Enhancement of vascular relaxation in rat aorta by phytoestrogens from Curcuma comosa Roxb. Vasc. Pharmacol., 51: 284-290.
CrossRef - Jennings, L.K., 2009. Role of platelets in atherothrombosis. Am. J. Cardiol., 103: 4A-10A.
Direct Link - Koh, K.K., C. Cardillo, M.N. Bui, L. Hathaway and G. Csako et al., 1999. Vascular effects of estrogen and cholesterol-lowering therapies in hypercholesterolemic postmenopausal women. Circulation, 99: 354-360.
PubMed - Laufs, U., V. La Fata and J.K. Liao, 1997. Inhibition of 3-hydroxy-3-methylglutaryl (HMG)-CoA reductase blocks hypoxia-mediated down-regulation of endothelial nitric oxide synthase. J. Biol. Chem., 272: 31725-31729.
CrossRef - Lokhande, P.D., B.S. Kuchekar, A.R. Chabukswar and S.C. Jagdale, 2006. Nitric oxide: Role in biological system. Asian J. Biochem., 1: 1-17.
CrossRefDirect Link - Niumsakul, S., A. Hirunsaree, S. Wattanapitayakul, N. Junsuwanitch and K. Prapanupun, 2007. An antioxidative and cytotoxic substance extracted from Curcuma comosa Roxb. J. Thai. Trad. Altern. Med., 5: 24-29.
Direct Link - Nurulhuda, M.H., A. Azlan, A. Ismail, Z. Amom and F.H. Shakirin, 2012. Cholesterol-lowering and artherosclerosis inhibitory effect of sibu olive in cholesterol fed-rabbit. Asian. J. Biochem., Vol. 7. In Press.
Direct Link - Phivthong-Ngam, L., S.M. Bode-Boger, R.H. Boger, M. Bohme, R.P. Brandes, A. Mugge and J.C. Frolich, 1998. Dietary L-arginine normalizes endothelin-induced vascular contractions in cholesterol-fed rabbits. J. Cardiovasc. Pharmacol., 32: 300-307.
Direct Link - Pitsilos, S., J. Hunt, E.R. Mohler, A.M. Prabhakar and M. Poncz et al., 2003. Platelet factor 4 localization in carotid atherosclerotic plaques: correlation with clinical parameters. Thromb. Haemost., 90: 1112-1120.
CrossRef - Piyachaturawat, P., J. Charoenpiboonsin, C. Toskulkao and A. Suksamrarn, 1999. Reduction of plasma cholesterol by Curcuma comosa extract in hypercholesterolaemic hamsters. J. Ethnopharmacol., 66: 199-204.
CrossRefDirect Link - Piyachaturawat, P., C. Tubtim, A. Chuncharunee, P. Komaratat and A. Suksamrarn, 2002. Evaluation of the acute and subacute toxicity of a choleretic phloracetophenone in experimental animals. Toxicol. Lett., 129: 123-132.
CrossRef - Piyachaturawat, P., P. Srivoraphan, A. Chuancharunee, P. Komaratat and A. Suksamran, 2002. Cholesterol lowering effects of a choleretic phloracetophenone in hypercholesterolemic hamsters. Eur. J. Pharmacol., 439: 141-147.
CrossRef - Relou, I.A., C.M. Hackeng, J.W. Akkerman and E. Malle, 2003. Low-density lipoprotein and its effect on human blood platelets. Cell. Mol. Life. Sci., 60: 961-971.
Direct Link - Rosenson, R.S. and C.C. Tangney, 1998. Antiatherothrombotic properties of statins: Implications for cardiovascular event reduction. JAMA, 279: 1643-1650.
Direct Link - Saeed, S.A., M.Z.S. Urfy, F.W.A. Khimani and S.O. Saeed, 2004. From evolution of platelets to their role in heart attacks. J. Med. Sci., 4: 47-51.
CrossRefDirect Link - Sase, K. And T. Michel, 1995. Expression of constitutive endothelial nitric oxide synthase in human blood platelets. Life Sci., 57: 2049-2055.
CrossRef - Shetty, J.K., M. Prakash, S. Tripathy, M. Verma and P. Vikram, 2008. Effect of atorvastatin on paraoxonase activity in-patients with hyperlipidemia. Asian J. Biochem., 3: 139-142.
CrossRefDirect Link - Sodsai, A., P. Piyachaturawat, S. Sophasan, A. Suksamrarn and M. Vongsakul, 2007. Suppression by Curcuma comosa Roxb. of pro-inflammatory cytokine secretion in phorbol-12-myristate-13-acetate stimulated human mononuclear cells. Int. Immunopharmacol., 7: 524-533.
CrossRefDirect Link - Srisawat, S., L. Phivthong-Ngam, S. Unchern, U. Chantharaksri, P. Govitrapong and Y. Sanvarinda, 2003. Improvement of vascular function by chronic administration of a cyclo-oxygenase inhibitor in cholesterol-fed rabbits. Clin. Exp. Pharmacol. Physiol., 30: 405-412.
CrossRef - Sudano, I., L.E. Spieker, F. Hermann, A. Flammer and R. Corti et al., 2006. Protection of endothelial function: Targets for nutritional and pharmacological interventions. J. Cardiovasc. Pharmacol., 47: S136-S150.
Direct Link - Miller, V.T., J. LaRosa, V. Barnabei, C. Kessler and G. Levin et al., 1995. Effects of estrogen or estrogen/progestin regimens on heart disease risk factors in postmenopausal women: The Postmenopausal Estrogen/Progestin Interventions (PEPI) trial. J. Am. Med. Assoc., 273: 199-208.
CrossRefDirect Link - Theilmeier, G., C. Michiels, E. Spaepen, I. Vreys, D. Collen, J. Vermylen and M.F. Hoylaerts, 2002. Endothelial von Willebrand factor recruits platelets to atherosclerosis-prone sites in response to hypercholesterolemia. Blood, 99: 4486-4493.
CrossRefPubMedDirect Link - Kooy, M.V.M., J.W.N. Akkerman, S. van Asbeck, L. Borghuis and H.C. van Prooijen, 1993. UVB radiation exposes fibrinogen binding sites on platelets by activating protein kinase C via reactive oxygen species. Br. J. Haematol., 83: 253-258.
CrossRef - Van Willigen, G., G. Gorter and J.W. Akkerman, 1994. LDLs increase the exposure of fibrinogen binding sites on platelets and secretion of dense granules. Arterioscler. Thromb., 14: 41-46.
CrossRef - Vera, R., M. Sanchez, M. Galisteo, I.C. Villar and R. Jimenez et al., 2007. Chronic dysfunction in spontaneously hypertensive rats: Involvement of eNOS, caveolin and calmodulin expression and NADPH oxidase activity. Clin. Sci., 112: 183-191.
Direct Link - Wolf, A., C. Zalpour, G. Theilmeier, B.Y. Wang and A. Ma et al., 1997. Dietary L-arginine supplementation normalizes platelet aggregation in hypercholesterolemic humans. J. Am. Coll. Cardiol., 29: 479-485.
Direct Link - Lee, Y.M., W.F. Chen, D.S. Chou, T. Jayakumar and S.Y. Hou et al., 2010. Cyclic nucleotides and mitogen-activated protein kinases: Regulation of simvastatin in platelet activation. J. Biomed. Sci., Vol. 17.
CrossRef