
D. Sivaraman
Department of Pharmacology and Toxicology, C.L. Baid Metha College of Pharmacy, Jyothi Nagar, Thoraipakkam, Chennai 97, Tamil Nadu, India
P. Muralidharan
Department of Pharmacology and Toxicology, C.L. Baid Metha College of Pharmacy, Jyothi Nagar, Thoraipakkam, Chennai 97, Tamil Nadu, India
P. Panneerselvam
Department of Pharmaceutical Chemistry, C.L. Baid Metha College of Pharmacy, Jyothi Nagar, Thoraipakkam, Chennai 97, Tamil Nadu, India
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 4 | Page No.: 212-223







ABSTRACT
Alzheimer’s disease is a primary degenerative disease of the central nervous system. The progression of Alzheimer’s disease will ultimately lead to dementia, behavioral and cognitive impairments. Alzheimer’s is the commonest cause of dementia, a group of progressive condition which involves especially short-term memory loss, poor concentration, poor sense of time and space, difficulty in finding words or understanding other people, difficulty in perceiving and interpreting surroundings, mood changes and emotional upsets. The present study was designed to determine the ameliorating effect of ethanolic leaf extract of Ficus hispida Linn. (EEFH) on (amyloid beta) Aβ 25-35-induced cognitive deficits and oxidative stress in mice. Animals were treated with EEFH for periods of 4 weeks dose-dependently (200 and 400 mg kg-1) then received a single intracerebroventricular (i.c.v.) injection of Aβ 25-35 (10 μg mouse-1). Behavioral changes in the mice were evaluated using passive avoidance, Y-maze, Hole board and water-maze tests. Anti-oxidant enzymes and neuro-transmitter levels were also been estimated. EEFH at the dose of 400 mg kg-1 significantly ameliorated the cognitive and memory deficits caused by i.c.v. injection of Aβ 25-35. EEFH attenuated the Aβ-induced increase in brain levels of thiobarbituric acid reactive substances. There was an increase in glutathione peroxidase, glutathione reductase and super oxide dismutase activity in EEFH treated groups. The acetyl cholinesterase activity in the brain was lower in EEFH supplemented groups than the Aβ-injected group. EEFH treated group showed a significant alteration in behavior when compare to negative control in Y maze, Plus-maze and also in water maze tests. These findings suggest EEFH exerts a protective effect against cognitive deficits induced by Aβ 25-35 accumulation in Alzheimer’s disease, because of its potential antioxidant property.
PDF Abstract XML References Citation
Received: November 04, 2011;
Accepted: February 16, 2012;
Published: March 27, 2012
How to cite this article
D. Sivaraman, P. Muralidharan and P. Panneerselvam, 2012. Ameliorating Effect of Ethanol Leaf Extract of Ficus hispida Linn.
on Amyloid beta Aβ (25-35) Induced Cognitive Deficits and Oxidative Stress
in Alzheimer’s Mice. International Journal of Pharmacology, 8: 212-223.
DOI: 10.3923/ijp.2012.212.223
URL: https://scialert.net/abstract/?doi=ijp.2012.212.223
DOI: 10.3923/ijp.2012.212.223
URL: https://scialert.net/abstract/?doi=ijp.2012.212.223
INTRODUCTION
Alzheimer’s is characterized by selective neuronal cell death, the presence of extra cellular amyloid deposits in the core of neuritic plaques and the formation of intraneuronal neurofibrillary tangles in the brain of affected individuals. Alzheimer’s disease is a progressive neurodegenerative disease clinically characterized by dementia and neurobehavioral deterioration (Farbood et al., 2009). Neurochemically, these deficits are associated with dramatic losses of cortically projecting cholinergic neurons and by a reduction in the presynaptic markers of the cholinergic system, particularly in the areas of the brain related to memory and learning (Iqbal and Grundke-Iqbal, 2008).
The major component of the amyloid plaque core is the pathologically deposited amyloid-beta-peptide (Aβ), which is derived from Amyloid Protein Precursor (APP). Alterations in APP processing to secrete higher levels of the longer Aβ 42 peptide and a clinical correlation between elevated levels of amyloid β-peptide in the brain and cognitive decline has recently been reported (Ghosh et al., 2008). The pathological features of Alzheimer’s disease include deposition of senile plaques in different brain zones formed by aggregates of fibrillar Aβ peptide (Aβ), a neurotoxic metabolic product (Alinovi et al., 2006; Sadik et al., 2001).
Alzheimer’s Disease (AD) affects up to 15% of people over age 65 years and nearly half of people aged 85 years. Selective neuronal and synaptic loss in the hippocampus and cerebral cortex correlates with clinical symptoms and therefore, a clearer understanding of the mechanisms that cause neuronal death and dysfunction should lead to a greater understanding of the underlying pathophysiology of the disease and unveil potential therapeutic opportunities (LaFerla and Oddo, 2005; Hebert et al., 2003). Prevalence rates for more than 20 million people worldwide including 4.5 million Americans. This number is expected to double every 20 years as world populations continue to age. AD is now considered to be the third major cause of death in developed countries, after cardiovascular disease and cancer (Ma and Gang, 2008).
The total worldwide cost of dementia care is estimated to be US$315.4 billion annually and 77% of these costs occurred in the world’s more developed regions. The course of AD often takes a decade or more to progress, bringing with it a severe load to patients, families and society (Sato et al., 2002).
The course of AD often takes a decade or more to progress, bringing with it a severe load to patients, families and society. There is currently no known cure for AD. However, several drugs for treatment of AD symptoms have been approved by the US Food and Drug Administration (FDA) in recent years. Stem cell technology has emerged in the face of extraordinary advances for prevention, diagnosis and treatment of human diseases such as heart disease, diabetes, cancer and diseases of the nervous system, such as Parkinson’s disease and Alzheimer’s disease (AD). It offers great promise for tissue regeneration, cell replacement and gene therapy but clinical applications remain limited (Saeed and Mesaik, 2005).
Current AD therapies are merely palliative and only temporarily slow cognitive decline and treatments that address the underlying pathologic mechanisms of AD are still lacking. In this review, we focus on the current aspects of AD ranging from the key risk factors for AD, the underlying pathogenic events and the novel medications including disease-modifying properties (Khairallah and Kassem, 2011). Researches done by FDA and other associations have claimed that herbs contain natural composition of various compounds that affects our internal system (brain, heart and mind). In conventional medicine, those formulations are synthesized by chemical associations and are given in different doses to the person. We should also keep in mind that around 30% of the ingredients that are added in conventional medicine are extracted from natural resources like plants and herbs (Winston, 2005).
The leads of Central Nervous System (CNS) active medicinal plants which have emerged besides Rauwolfia serpentina, Mucuna pruriens for Parkinson’s disease, Ocimum sanctum as an anti stress agent, Withania somnifera as anxiolytic, Centella asiatica and Bacopa monnieri for learning and memory disorders. Bacopa monnieri, Benincasa hispida (Roy et al., 2008), Areca catechu (Jaiswal et al., 2011) and Ginkgo biloba for Alzheimer’s disease. The study related to Alzheimer’s Disease (AD) is focused towards the traditionally used rejuvenating and neurotonic agents. The recent trends in the pharmacological studies are based on the biochemical and molecular mechanism which leads to the development of CNS active principles from the herbal drugs (Anekonda and Reddy, 2005; Vinutha et al., 2007; Hussian and Manyam, 1997; Moustafa et al., 2007).
Ficus hispida Linn. belongs to the family Moraceae is a widely distributed and most commonly found in the interior and coastal regions distributed throughout India, Srilanka, Myanmar and Southern region of the Republic china. A mixture of honey and the juice of these fruit is a good antihemorrhagic (Peraza-Sanchez et al., 2002) but the barks and leaves are of particular interest from a medicinal point of view as an antidiarrhoeal (Mandal and Kumar, 2002), antidiabetic (Gosh et al., 2004) and as cardio protective (Shanmugarajan et al., 2007) among others.
The current study was undertaken to evaluate the ameliorating effect of ethanolic leaf extract of Ficus hispida Linn (EEFH) , till now no pharmacological evaluation has been done on FH especially in leaf for its anti-alzhemerian activity. This prompted us to pursue the activity and was examined for their efficacy.
MATERIALS AND METHODS
Plant material: The fresh Ficus hispida Linn (FH) leaves were collected from (Perannakavur, Changlepet, Tamil Nadu, India) western Ghats of South India during June 2008. The plant was identified and authenticated by Dr. G.V.S. Murthy, Joint Director, Botanical Survey of India, Southern Circle, Coimbatore (BSI/SC/5/23/08-09/Tech-738). The specimen voucher was deposited in the Department of Pharmacology and Toxicology, C.L. Baid Metha College of Pharmacy, Chennai, Tamil Nadu, India.
Preparation of the ethanolic extract of FH: The fresh leaves of FH were collected and washed with running water and shade dried at room temperature. One kilogram of the dried leaf was made in to coarse powder. The powder was passed through a 60 No. mesh sieve. The grounded power was extracted with ethanol in water bath at room temperature. The solvent was then removed by filtration and fresh solvent was added to the plant material (Ravishankara et al., 2002). The extract process was twice repeated. The combined filtrates were then evaporated under reduced pressure to give a dark green viscous mass. The extract was stored at 0-4°C. The percentage yield was 20.2%w/w.
Phytochemical screening: The freshly prepared ethanol leaf extract of FH (EEFH) was qualitatively tested for the presence of chemical constituents. Phytochemical screening of the extract was performed using the following reagents and chemicals: Alkaloids with Mayer’s, Hager’s and Dragendorffs reagent; Flavonoids with the use of sodium acetate, ferric chloride, amyl alcohol; Phenolic compounds and tannins with lead acetate and gelatin; carbohydrate with Molish’s, Fehling’s and Benedict’s reagent; proteins and amino acids with Millon’s, biuret and xanthoprotein test. Saponins was tested using hemolysis method; Gum was tested using Molish’s reagent and Ruthenium red; Coumarin by 10% sodium hydroxide and quinones by Concentrated Sulphuric acid. These were identified by characteristic color changes using standard procedures (Trease and Evans, 1989).
These were identified by characteristic color changes using standard procedures.
The screening results were as follows: Alkaloids +ve; carbohydrates +ve; proteins and amino acids +ve; saponins +ve, steroids -ve; sterols +ve; phenols +ve; flavonoids +ve; gums and mucilage +ve; glycosides +ve; saponins +ve; terpenes +ve and tannins -ve. Where +ve and –ve indicates the presence and absence of compounds.
Animals: Young adult Swiss albino mice of either sex, weighing (18-20 g) were obtained from animal house of C.L. Baid Metha College of Pharmacy, Chennai, Tamil Nadu, India. Animals were kept in raised mesh bottom cages to prevent coprophagy. The animals were maintained in colony cages at 25±2°C, relative humidity 50-55% maintained under 12:12 h light and dark cycle. The animals were fed with Standard animal feed (Hindustan Lever Ltd., Bangalore, India) and water ad libitum, animals were acclimatized to the laboratory conditions prior to experimentation. All the experiments were conducted between 10-00 and 17.00 h and were in accordance with the ethical guidelines of the International association for Study of Pain (Zimmermann, 1983). All experiments were carried out according to the guidelines for care and use of experimental animals and approved by Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA). the study was approved by the Institutional Animal Ethical Committee. C.L. Baid Metha College of Pharmacy, Chennai, Tamil Nadu, India.
Acute toxicity study: This was performed for the extracts to ascertain safe dose by the acute oral toxic class method by the Organization of Economic Cooperation and Development (OECD). A single administration of starting dose of 2000 mg kg-1 body weight p.o. of the EEFH was administered to three female mice and the mice were observed for three days to evaluate considerable changes in body weight and other signs of toxicity. There was no considerable change in body weight before and after treatment and no sign of toxicity were observed. When the experiment was repeated again with same dose level, 2000 mg kg-1 b.wt. p.o. of plant extract for 7 more days and observed for fourteen days no change was observed from the experiments.
Induction of amnesia
Intracerebroventricular injection of Aβ peptide: The administration of a Aβ 25-35 was performed by identifying the bregma pointing the skull, each mouse was injected at bregma with a 50 μL Hamilton micro syringe fitted with a 26-gauge needle that was inserted to a depth of 2.4 mm in brief, the needle was inserted unilaterally 1 mm to the right of the middle point equidistant from each eye, at an equal distance between the eyes and ears and perpendicular to the plane of the skull. Mice exhibited normal behavior with in 1 min after injection. The injection volume was 10 μL; the memory impairment due to the injection was examined on the second day of i.c.v. injection by step down inhibitory avoidance (Laursen and Belknap, 1986).
Experimental design: Animals were divided into four groups of each ten animals:
| Group I: | Animals injected by 1% w/v CMC |
| Group II: | Animal injected with β-Amyloid (25-35) 10 μL by intracerebroventricular injection (i.c.v.) |
| Group III: | Animals injected by Aβ peptide and treated with 200 mg kg-1 (p.o.) EEFH |
| Group IV: | Animals injected by Aβ peptide and treated with 400 mg kg-1 (p.o.) EEFH |
Amnesia is induced by intracerebroventricular injection (i.c.v.) of Aβ peptide, i.c.v. Injection for the II, III and IV groups were performed on the 21st day of the pre treated and continued for 7 days. Control animals are injected with only 1% w/v CMC.
In vivo pharmacological examination (behavioral studies)
Step down inhibitory avoidance: The apparatus consist of 50x25x25 cm acrylic box, whose floor consisted of parallel 1.0 cm apart. A 7.0 cm wide, 2.5 cm high, 25.0 cm long platform occupied the centre floor. In the training session, immediately after stepping down placing their four paws on the grid the animals received a 0.4 MA, 2.0 sec scrambled foot shock. In test session no foot shock was given and step-down latency was used as a measure of retention (to a ceiling of 300 sec) one-trial step-down inhibitory avoidance in rats and mice involves the activation of two separate memory types, a Short-Term Memory (STM) system and a Long-Term Memory (LTM) system. Therefore, retention tests were carried out 90 min after training to evaluate STM and 7 days after training to evaluate long-term memory. The same mice were used for both tests, as testing for STM has been found not to affect LTM retention scores in previous studies (Dhingra et al., 2006).
Open field habituation: In order to control for possible effects on locomotor activity, animals were explored twice, with a 24 h interval, to a 40x50x60 cm open field whose brown linoleum floor was divided into 12 equal squares by white lines. In both sessions, the animals were placed in the rear left square and left to explore it freely for 5 min during which time the number of lines crossing, head dippings and rearings were counted (Saitoh et al., 2006; Yamada et al., 2005).
Water maze task: The experimental apparatus consisted of a circular water tank (diameter-100 cm; height-35 cm), containing water at 28°C to a depth of 15 cm and rendered opaque by adding titanium dioxide. A platform (diameter 4.5 cm; height 14.5 cm) was submerged 1 cm below the water surface and placed at the midpoint of one quadrant. After several trials, the test was conducted after injection of β amyloid peptide. In each training trial, the time required to escape on to the platform was recorded (Pietropaolo et al., 2008).
Y-maze task: Y-maze task is used to measure the spatial working through the spontaneous alternation of behaviour. The maze is made of black painted wood. Each arm is 40 cm long, 13 cm high, 3 cm wide at the bottom, 10 cm wide at the top and converges at an equal angle. Each mouse is placed at the end of one arm and allowed to move freely through the maze during an 8 min session. Mice tend to explore the maze systematically, entering each arm in turn. The ability to alternate requires that the mice know which arm they have already visited. The series of arm entries, including possible returns into the same arm, are recorded visually. Alternation is defined as the number of successive entries into the three arms, on overlapping triplet sets. The percentage of alternation is calculated as the ratio of actual alternations, defined as the total number of arm entries minus two and multiplied by 100 (Hiramatsu and Inoue, 2000).
Estimation of neuro transmitter and metabolic enzymes
Estimation of acetylcholinesterase enzyme: Twenty milligram of brain tissue per mL of phosphate buffer (pH 8, 0.1 m) was homogenized in a Potter-Elvehjem homogenizer. A 0.4 mL aliquot of brain homogenate was added to a cuvette containing 2.6 mL of 0.1 m phosphate buffer (pH 8). One hundred microliter of the DTNB reagent was added to the photo cell. The absorbance was measured at 412 nm then acetylthiocholine iodide was added. Changes in absorbance were recorded and the change in absorbance per minute was calculated (Vinutha et al., 2007). The enzyme activity is expressed as μmol/min/mg tissue.
Estimation of monoamine oxidase (MAO) enzyme: The assay mixture contain 4 mm of serotonin as specific substrates for MAO-A, 250 μL solution of the mitochondrial fraction and 100 mm sodium phosphate buffer (pH 7.4) up to the final volume of 1 mL, the reaction was allowed to proceed at 37°C for 20 min and stopped by adding 1 m HCl (200 μL), the reaction product was extracted with 5 mL of butyl acetate, the organic phase was measured at wavelength of 280 nm in a spectrometer. Blank samples were prepared by adding 1 m HCl (200 μL) prior to the reaction and worked subsequently in the same manner (Muralidharan et al., 2009).
Estimation of dopamine: On the day of experiment mice were sacrificed, whole brain was dissected out and separated the subcortical region (including the striatum). Weighed a specific quantity of tissue and was homogenized in 3 mL HCl Butanol in a cool environment. The sample was then centrifuged for 10 min at 2000 rpm. Supernatant phase was removed and added to an Eppendorf reagent tube containing heptane and HCl. After 10 min, shake the tube and centrifuged under same conditions to separate two phases. Upper organic phase was discarded and the aqueous phase was used for dopamine assay (Schlumpf et al., 1974).
Dopamine assay: To 0.02 mL of the HCl phase, 0.4 mL HCl and 0.01 mL EDTA/Sodium Acetate buffer (pH 6.9) were added, followed by 0.01 mL iodine solution for oxidation. The reaction was stopped after 2 min by the addition of sodium thiosulphate in 5 M Sodium hydroxide. 10 M Acetic acid was added 1.5 min later. The solution was then heated to 100°C for 6 min. When the sample again reaches room temperature, excitation and emission spectra were read (330 to 375 nm) in a spectrofluorimeter. Compared the tissue values (fluorescence of tissue extract minus fluorescence of tissue blank) with an internal reagent standard (fluorescence of internal reagent standard minus fluorescence of internal reagent blank). Tissue blanks for the assay were prepared by adding the reagents of the oxidation step in reversed order (sodium thiosulphate before iodine). Internal reagent standards were obtained by adding 0.005 mL double distilled water and 0.1 mL HCl Butanol to dopamine standard.
Estimation of serotonin: Three milligram of mice brain is homogenized in 0.1 mL of HCl-n butanol for 1 min in glass homogenizer. The sample was centrifuged for 10 min at 2000 rpm. The supernatant phase was removed and added to Eppendorf reagent tubes containing 0.2 mL of heptane and 0.025 mL of HCl 0.1 M. After 10 min of vigorous shaking the tube was centrifuged under the same conditions as above in order to separate the two phases. The aqueous phase was taken and o-phthaldialdehyde was added. The fluorophore was developed by heating to 100°C for 10 min. After the sample reach the equilibrium with ambient temperature the intensity readings at 360-470 nm was taken in microcuvette (Kepe et al., 2006).
Estimation of glutamate: Weighed quantity of brain portion was homogenized with perchloric acid and centrifuge for 10 min at 3000 rpm. Adjust supernatant fluid to pH 9 with phosphate solution. Allow to stand 10 min, in an ice bath and then filter through a small, fluted filter paper. Allow to warm to room temperature, dilute and take 1.0 mL for the assay (Sivaraman et al., 2010).
Estimation of total protein: Total protein was estimated in brain homogenate by using suitable reagents method (Lowry et al., 1951).
Estimation of antioxidant enzyme
Estimation of superoxide dismutase (SOD): The assay mixture contained 1 mL of pyrogallol-tris-DEPTA, 0.2 mL of suitably diluted tissue and 0.8 mL of water. The rate of pyrogallol autoxidation is taken from the increase in absorbance at 420 nm. The activity of SOD was expressed as units/minutes/mg protein. One unit of the enzyme is defined as the amount of enzyme which inhibits the rate of pyrogallol auto oxidation by 50%. The SOD is expressed as units/min/mg protein (Byers et al., 2006).
Estimation of catalase (CAT): Homogenize the tissue with phosphate buffer at 1 to 4°C and centrifuged. Stirred the sediment with cold phosphate buffer and allowed to stand in the cold condition with occasional shaking. Repeat the extraction once or twice, supernatants were combined and used for assay. H2O2-phosphate buffer was taken in one cuvette, added 0.01- 0.04 mL sample and read against a control cuvette containing enzyme solution without H2O2 phosphate buffer at 240 nm. It was noted for a decrease in the optical density from 0.450 to 0.400. This value was used for the calculations (Pari and Latha, 2004).
Estimation of glutathione peroxidase (GPx): The reaction mixture consisting of 0.2 mL each of EDTA, sodium azide and 0.4 mL of phosphate buffer, 0.1 mL of suitably diluted tissue was incubated at 37°C at different time intervals. The reaction was arrested by the addition of 0.5 mL of TCA and the tubes were centrifuged at 2000 rpm. To 0.5 mL of supernatant, 4 mL of Disodium hydrogen phosphate and 0.5 mL DTNB were added and the color developed was read at 420 nm immediately (Resende et al., 2008). The activity of GPx is expressed as μmoles of glutathione oxidized/minutes/mg protein.
Estimation of glutathione reductase (GR): The reaction mixture containing 1 mL of phosphate buffer, 0.5 mL of EDTA. 0.5 mL of oxidized glutathione and 0.2 mL of NADPH was made up to 3 mL with water. After the addition of suitably diluted tissue, the change in optical density at 340 nm was monitored for 2 min at 30 sec intervals (Sapakal et al., 2008). The activity of GR is expressed as nmoles of NADPH oxidized/minute/mg protein.
Estimation of lipid peroxidation: About 0.2 mL of tissue homogenate, acetic acid, sodium dodecyl sulphate and thiobarbituric acid were added. The volume of the mixture was made up to 4.0 mL with distilled water and then heated at 95°C in a water bath for 60 min. After incubation the tubes were cooled to room temperature and final volume was made to 5.0 mL in each tube, n-butanol: pyridine mixture was added and the contents were vortexed thoroughly for 2 min. After centrifugation at 3000 rpm for 10 min, the organic upper layer was taken and its optical density read at 532 nm against an appropriate blank without the sample (Alper et al., 1998). The levels of lipid peroxides were expressed as nmoles of Malondialdehyde (MDA)/min/mg protein in brain homogenate.
Histopathological examination: The mice from each group were anesthetized by i.p. (intraperitoneal) injection of thiopentone (100 mg kg-1),The brain was removed with out any injury after opening the skull. The collected sample were washed with normal saline and fixed in 10% neutral formalin for 48 h for further histological observation. Paraffin section were taken at 5 μm thickness processed in alcohol-xylene series and was stained with Haematoxylin-eosin dye. The section were examined microscopically for histopathological changes especially in the hippocampus and in the cortex (Takanaga et al., 2003).
Statistical analysis: All values are expressed as Mean±SEM. Data were analyzed by non-parametric ANOVA followed by Dunnett’s multiple comparison tests and other data were evaluated using Graph Pad PRISM software. A p-value<0.05 was considered significantly different.
RESULTS
Effect of EEFH on step down inhibitory avoidance: There was a significant difference between the control group (I) and the negative control group (II) in the step down latency in the STM and LTM. The EEFH treated groups show significant tendency to increase the retention of STM and LTM. The low dose and high dose shown significance (p<0.05) when compared with the negative control was shown in Fig. 1.
Effect of EEFH on water maze: There is an increase in escape latency in negative control group when compared with the control group (p<0.01) of the two groups of amnesia induced animals, both showed decreased time to escape on to the escape platform. The group treated with 200 mg kg-1 EEFH group 400 mg kg-1 treated showed the significance of (p<0.01 and p<0.01), respectively as shown in Fig. 2.
Effect of EEFH on exploratory behavior: The amnesia induced group negative control indicated decrease in the exploratory behavior i.e., head dippings, rearings, line crossings by the (p<0.05, p<0.05 and p<0.05), respectively in comparison with the control group I. The results presented by the treated groups shows significance (p<0.05, p<0.05 and p<0.01) increase in the head dippings and line crossings in respect of 200 mg kg-1 when compare with the negative control group. The 400 mg kg-1 treated group shown significance of (p<0.01, p<0.01 and p<0.01) head dippings, rearings and line crossings, respectively as shown in Fig. 3.
Effect of EEFH on Y-maze: The amnesia induced group indicated decrease in the alternation of behaviour by the (p<0.01) in comparison with the control group I. The results presented by the treatment groups shows significance by (p<0.01 and p<0.01) increase in the alternation of behaviour in respect of 200 mg kg-1 of EEFH and 400 mg kg-1 of EEFH when compared with negative control group as shown in Fig. 4.
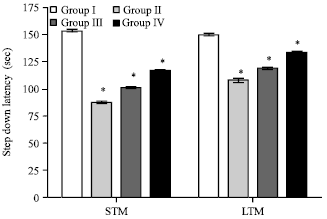 | |
| Fig. 1: | Effect of EEFH on step down inhibitory avoidance (*p<0.05) |
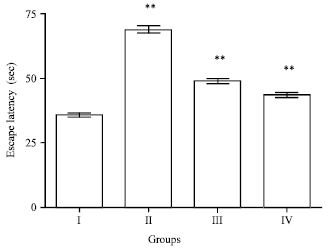 | |
| Fig. 2: | Effect of EEFH on water maze (**p<0.01) |
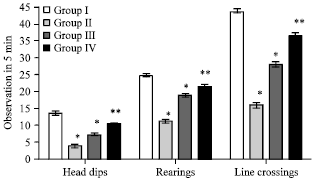 | |
| Fig. 3: | Effect of EEFH on exploratory behavior (*p<0.05, **p<0.01) |
Effect of EEFH on AchE (activities of acetylcholine esterase) activity: Injection of Aβ 25-35 significantly (p<0.01) increased the AchE activity when compared with control group. In the treated group there was a significance (p<0.01) reduction in enzyme levels on both 200 and 400 mg kg-1 of EEFH treated mice as shown in Fig. 5.
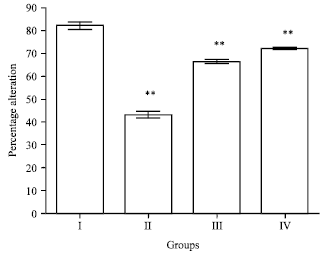 | |
| Fig. 4: | Effect of EEFH on Y-maze (**p<0.01) |
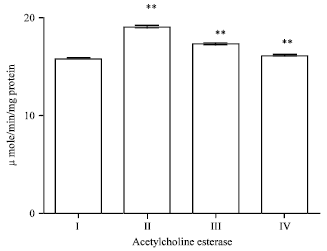 | |
| Fig. 5: | Effect of EEFH on AchE activity (**p<0.01) |
Effect of EEFH on MAO (monoamine oxide) -A: The enzyme activity in Aβ 25-35 induced group had significantly increased (p<0.01) when compared with control group I. The activity of MAO-A in 200 mg kg-1 of EEFH treated animals showed a significant reduction (p<0.01). The activity of MAO-A in 400 mg kg-1 of EEFH treated animals significant reduction (p<0.01) in comparison negative control group as shown in Fig. 6.
Effect of EEFH on dopamine: The enzyme activity of Aβ 25-35 induced group had decreased (p<0.01), respectively when compared with control group I. The dopamine activity in 200 and 400 mg kg-1 of EEFH treated groups showed a significance increase in (p<0.01 and p<0.01) on comparison with the negative control as shown in Fig. 7.
Effect of EEFH on serotonin: The enzyme activity in Aβ 25-35 induced group had decreased (p<0.01), respectively.
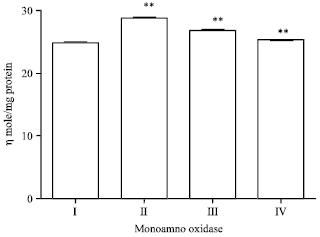 | |
| Fig. 6: | Effect of EEFH on MAO-A (**p<0.01) |
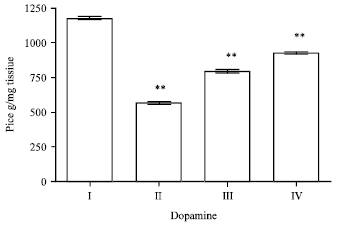 | |
| Fig. 7: | Effect of EEFH on dopamine (**p<0.01) |
When compared with control group. The serotonin activity in 200 and 400 mg kg-1 of EEFH treated groups showed a significance increase in (p<0.01 and p<0.01) on comparison with the negative control as shown in Fig. 8.
Effect of EEFH on glutamate level: There was an increase in the glutamate level in Aβ 25-35 induced group (p<0.01), respectively. The glutamate activity in 200 and 400 mg kg-1 EEFH treated groups showed a significance increase decreased in (p<0.01 and p<0.01) on comparison with the negative control as shown in Fig. 9.
Effect of EEFH on protein level: In the present study the protein level of Aβ 25-35 induced group show significant (p<0.05) increase when compared with control group. A significant decrease (p<0.01) in the level of protein was observed for 200 and 400 mg kg-1 of EEFH treated group as the results shown in Fig. 10.
Effect of EEFH on superoxide dismutase: SOD levels in the brain was significantly reduced (p<0.01) in Aβ 25-35 induced group when compared to control group.
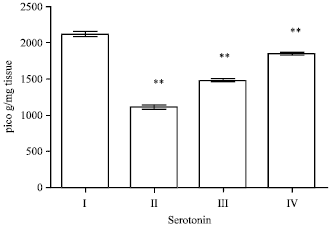 | |
| Fig. 8: | Effect of EEFH on serotonin (**p<0.01 ) |
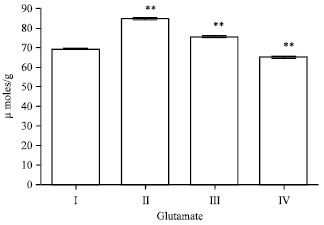 | |
| Fig. 9: | Effect of EEFH on glutamate level (**p<0.01 ) |
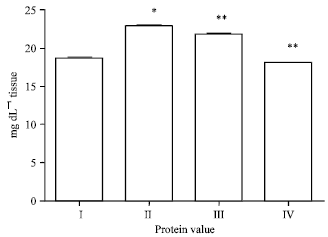 | |
| Fig. 10: | Effect of EEFH on protein Level (*p<0.05, **p<0.01) |
Treatment with EEFH at 200 and 400 mg kg-1 dose level showed significant increase (p<0.01 and p<0.01), respectively when compared with negative control group as the results were shown in Fig. 11.
Effect of EEFH on glutathione peroxidase: The GPx in the dementia induced mice (group II) shown significant (p<0.01) reduction in the enzyme activity when compared with control group.
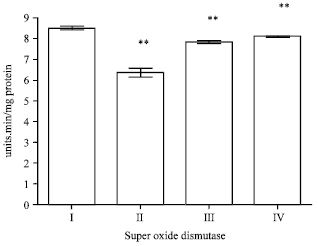 | |
| Fig. 11: | Effect of EEFH on Super oxide dismutase (**p<0.01) |
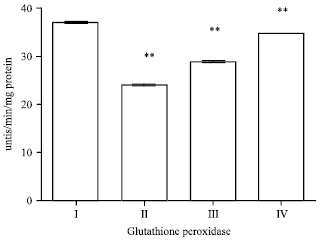 | |
| Fig. 12: | Effect of EEFH on Glutathione Peroxidase (**p<0.01) |
The treatment with EEFH at 200 and 400 mg kg-1 shown the significance (p<0.01 and p<0.01), respectively when compared with negative control group as shown in Fig. 12.
Effect of EEFH on glutathione reductase: The glutathione reductase activity of Aβ 25-35 induced group shown significant (p<0.01) decrease in the activity when compared with control group. The two groups with 200 and 400 mg kg-1 doses of EEFH treated animals showed significance increase (p<0.01) when compared with negative control group. The results are shown in Fig. 13.
Effect of EEFH on catalase level: There was a decrease in the catalase level in Aβ 25-35 induced group when compared with the control group. The two groups with 200 and 400 mg kg-1 doses of EEFH treated animals showed significant increase (p<0.01) when compared with negative control group. The results are shown in Fig. 14.
Effect of EEFH on lipid peroxidation level: There was an increase in the lipid peroxidation level in Aβ 25-35 induced group when compared with the control group.
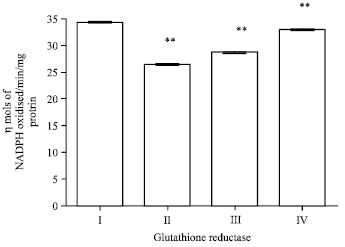 | |
| Fig. 13: | Effect of EEFH on glutathione reductase (**p<0.01) |
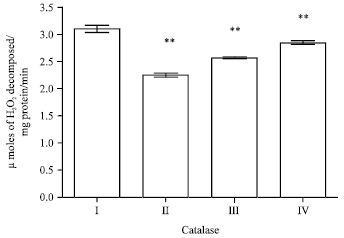 | |
| Fig. 14: | Effect of EEFH on catalase level (**p<0.01) |
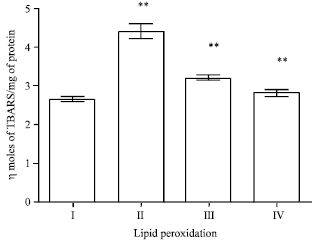 | |
| Fig. 15: | Effect of EEFH on lipid peroxidation level (**p<0.01) |
The two groups with 200 and 400 mg kg-1 doses of EEFH treated animals showed significant decrease (p<0.01) when compared with negative control group. The results are shown in Fig. 15.
Effect of EEFH on hippocampai neuronal degeneration: There was a increase in neuronal degeneration and decrease in the number of neuronal cells in hippocampal region in Aβ 25-35 induced group (negative control) group when compared with the control group .The group treated with 200 and 400 mg kg-1 EEFH showed the significant decrease in brain cell edema and improves the neuronal configuration when compared with negative control group as shown in Fig. 16.
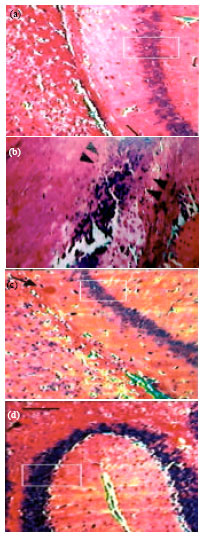 | |
| Fig. 16(a-d): | Effect of EEFH on hippocampai neuronal degeneration: (a) Control group, (b) Aβ 25-35 Treated group, (c) EEFH 200 mg kg-1 Treated group and (d) EEFH 400 mg kg-1 Treated group |
DISCUSSION
The present study has revealed the neuroprotective effect of EEFH on Aβ 25-35 induced cognitive deficits in mice. Ficus hispida Linn. is a medicinal plant with antioxidant properties. Intracerebroventricular injection of Aβ 25-35 induced impairment of memory assessed by passive avoidance and Morris water tests. Aβ 25-35 has the potential to induced oxidative stress in the brain cholinergic hypo function, elevation of AchE and MAO (Akiyama et al., 2003). Moreover, it has been reported that it induces the production of hydrogen peroxide and lipid peroxide in hippocampal neurons of the mice brain. And the induction of 4-hydroxy-2-nonenal and 8-hydroxy-2-deoxyguanosine (a marker of oxidative damage to DNA) immunoreactivities following infusion of Aβ 1-42 in mice brain. In the present study, we have found a significant decrease in the level of antioxidant enzymes and the elevated of AchE and MAO in mice brain after a single injection with Aβ 25-35 (D’Almeida et al., 1996). Furthermore in balance each antioxidant enzyme were also observed and demonstrated that continuous i.c.v. infusion of Aβ 1-42 in mice resulted in a significant decrease in protein expression of SOD, GPx and glutathione-s-transferase in mice brain. Glutathione (GSH) is the major non-protein thiols antioxidant in mammalian cells and it is considered to be the main intracellular redox buffer. GSH protects cellular protein-thiols against irreversible loss, thus preserving protein function.
The position that GSH takes part in the redox-sensitive signaling cascade has been receiving growing support. One of the most importance GSH-dependent detoxifying processes involved is Glutathione Peroxidase (GPx) which plays a central role in the removal of hydrogen and organic peroxides and leads to the formation of oxidized Glutathione (GSSG). GSSG is reduced back to its thiols form (GSH) by the ancillary enzyme Glutathione reductase (GRD), leading to the consumption of NADPH which is mainly produced in the pentose phosphate pathway. GSH also takes part in xenobiotic conjugation with the assistance of several glutathione S-transferase isoenzymes. GSH conjugates or GSSG can be eliminated from the cell by the family of ATP dependent transporter pumps. The inhibition GSH synthesis leads to an increase in Aβ induced cell death and intracellular Aβ accumulation (Polidori, 2003).
It’s been suggested that antioxidant might contribute to the prevention of Alzheimer’s disease. Antioxidant such as beta carotene and vitamins C, E and A may protect cells from the type of damage that leads to aging in the brain and tissues (Saeed et al., 2005). Both vitamin C and E are antioxidant which are likely reduce oxidative stress and injury in the central nervous system, this may reduce the Aβ plaque deposition in the neuronal cells (Cole et al., 2005).
Presently in present study, we found that treatment with EEFH ameliorated cognitive deficits in Aβ 25-35 injected mice, especially shows increase in step down inhibitory avoidance and in the exploratory behaviour. Step-down inhibitory avoidance task is a classic task of memory with a strong aversive component. EEFH exerted significant effects in short-term retention and increase in long-term retention of this task. We also examined the effect of EEFH on the open-filed test as it is presented in which allowed us to study the effect of the treatment on the general locomotor activity of the animals, results found that animals treated with EEFH had an increase in the number of head dippings, line crossings and rearings in the open filed. These results of our experiments have been consistent with the favorable effect on locomotor activity. In the water-maze test, consumption of EEFH decreases the escape latency almost to normal levels in the dose dependent manner. It is possible that neuroprotection plays a role in that favorable effect of EEFH on Aβ 25-35 induced cognitive deficits.
The AchE activity has been shown to be increased around Aβ plaques in Alzheimer’s brain. The calcium influx followed by oxidative stress is involved in the increase in activity of AchE induced by Aβ 25-35 peptide, decreasing cell membrane order and ultimately leading to the exposure of more active enzyme (Ban et al., 2006). The observation that Aβ peptide increases AchE activity indicates that it can be possible to ameliorate cholinergic function, by inhibiting Aβ induce increase in AchE activity. The AchE activity in the brain was increased in mice treated with Aβ 25-35 when compared with the normal. In addition, the Aβ 25-35 induced increase in AchE was attenuated by EEFH treatment.
MAO is an important enzyme in the metabolism of a wide range of monoamine neurotransmitters, including noradrenalin, dopamine and 5-hydroxytryptamine. MAO exists in two forms, A and B. MAO-A in the metabolism of the major neurotransmitter monoamines. MAO-A inhibitors have been accepted to treat depression (Kaakkola and Wurtman, 1992). MAO-A and MAO-B in the brain have been implicated in the etiology of Alzheimer’s disease. Elevations in MAO-A in Alzheimer neurons have been linked to increase in neurotoxic metabolites and neuronal loss. Free radical generation by Aβ or other noxious stimuli could contribute to an imbalance between the production of nitric oxide and oxygen radicals and participate in oxidative stress. In conclusion, we suggest that markedly EEFH improves cognitive deficits induced by Aβ 25-35 and this effect is mediated by the antioxidant properties of EEFH.
CONCLUSION
In conclusion from the observation, the neuroprotective activity of the plant Ficus hispida Linn. on Alzheimer’s type of dementia is due to the inhibiting activity against AchE, MAO, glutamate, lipidperoxidation there by it elevates Dopamine, Serotonin and free radical scavenging activity. Further studies have to be anticipated on the various level of extracts and isolated principles of the plant for various studies with immunomodulatory changes with respect to the brain mitochondrial level DNA damage.
ACKNOWLEDGMENT
The authors are grateful to Dr. S. Venkataraman (Director of C.L. BAID METHA Foundation for Pharmaceutical Education and Research, Chennai) and for his technical and secretarial assistance. The authors have no conflict of interest to report.
REFERENCES
- Akiyama, T., T. Yamazaki, H. Mori and K. Sunagawa, 2003. Inhibition of cholinesterase elicits muscarinic receptor-mediated synaptic transmission in the rat adrenal medulla. Auton. Neurosci., 107: 65-73.
PubMedDirect Link - Alper, G., M. Cinar, C. Can, G. Mentes, B. Ersoz and A. Evinc, 1998. The effects of vitamin E on catalase activies in various rat tissues. Turk. J. Med. Sci., 28: 127-131.
Direct Link - Moustafa, A.M.Y., F.S. Kouam, K. Aisha, E. Asma and A. Shamsher et al., 2007. Phytochemical investigation and biological evaluation of Schinus terebinthifolius. Res. J. Phytochem., 1: 1-11.
CrossRefDirect Link - Anekonda, T.S. and P.H. Reddy, 2005. Can herbs provide a new generation of drugs for treating Alzheimer's disease? Brain Res. Rev., 50: 361-376.
CrossRefPubMedDirect Link - Ban, J.Y., S.O. Cho, S.B. Koh, K.S. Song, K. Bae and Y.H. Seong, 2006. Protection of amyloid β protein (25-35)-induced neurotoxicity by methanol extract of Smilacis chinae rhizome in cultured rat cortical neurons. J. Ethnopharmacol., 106: 230-237.
PubMed - Roy, C., T.K. Ghosh and D. Guha, 2008. Dose dependent activity of Benincasa hispida on colchicine induced experimental rat model of alzheimer's disease. Int. J. Pharmacol., 4: 237-244.
CrossRefDirect Link - Cole, G.M., G.P. Lim, F. Yang, B. Teter and A. Begum et al., 2005. Prevention of Alzheimer's disease: Omega-3 fatty acid and phenolic anti-oxidant interventions. Neurobiol. Aging, 26: 133-136.
CrossRef - D'Almeida, V., R. Camarini, L.A. Azzalis, V.B. Junqueira and E.A. Carlini, 1996. Chronic fenfluramine treatment of rats with different Ages: Effects on brain oxidative stress-related parameters. J. Biochem. Toxicol., 11: 197-201.
PubMed - Dhingra, D., M. Parle and S.K. Kulkarni, 2006. β-Alanine protects mice from memory deficits induced by ageing, scopolamine, diazepam and ethanol. Indian J. Pharma. Sci., 68: 216-221.
CrossRefDirect Link - Farbood, Y., A. Sarkaki and M. Badavi, 2009. Preventive effect of grape seed hydroalcholic extract on dementia type of alzheimer's disease in aged male rats. Int. J. Pharmacol., 5: 257-262.
CrossRefDirect Link - Ghosh, A.K., S. Gemma and J. Tang, 2008. Β-Secretase as a therapeutic target for Alzheimer's disease. Neurother. J. Am. Soc. Exp. NeuroTher., 5: 399-408.
PubMed - Sadik, G., K. Takeda, H. Kaji, M. Taoka and T. Shinoda, 2001. In vitro generation of amyloid A4 peptide from amyloid protein precursor through nonspecific proteolysis. Pak. J. Biol. Sci., 4: 289-292.
CrossRefDirect Link - Gosh, R., K. Sharatchandra, S. Rita and I.S. Thokchom, 2004. Hypoglycemic activity of Ficus hispida (bark) in normal and diabetic albino rats. Indian J. Pharmacol., 36: 222-225.
Direct Link - Hebert, L.E., P.A. Scherr, J.L. Bienias, D.A. Bennett and D.A. Evans, 2003. Alzheimer disease in the US population: Prevalence estimates using the 2000 census. Arch. Neurol., 60: 1119-1122.
CrossRefPubMedDirect Link - Hiramatsu, M. and K. Inoue, 2000. Improvement by low doses of nociceptin on scopolamine-induced impairment of learning and/or memory. Eur. J. Pharmacol., 395: 149-156.
CrossRefPubMedDirect Link - Hussian, G. and B.V. Manyam, 1997. Mucuna pruriens proves more effective than L-DOPA in Parkinson's disease animal model. Phytother. Res., 11: 419-423.
CrossRefDirect Link - Kepe, V., J.R. Barrio, S.C. Huang, L. Ercoil and P. Siddarth et al., 2006. Serotonin 1A receptors in the living brain of Alzheimer's disease patients. PNAS, 103: 702-707.
CrossRefDirect Link - LaFerla, F.M. and S. Oddo, 2005. Alzheimer's disease: Aβ, tau and synaptic dysfunction. Trends Mol. Med., 11: 170-176.
CrossRefPubMedDirect Link - Laursen, S.E. and J.K. Belknap, 1986. Intracerebroventricular injections in mice. Some methodological refinements. J. Pharmacol. Methods, 16: 355-357.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Ma, X. and D.R. Gang, 2008. In vitro production of huperzine A, a promising drug candidate for Alzheimer's disease. Phytochemistry, 69: 2022-2028.
CrossRefPubMedDirect Link - Mandal, S.C. and C.K.A. Kumar, 2002. Studies on anti-diarrhoeal activity of Ficus hispida. leaf extract in rats. Fitoterapia, 73: 663-667.
CrossRefPubMedDirect Link - Khairallah, M.I. and L.A.A. Kassem, 2011. Alzheimer's disease: Current status of etiopathogenesis and therapeutic strategies. Pak. J. Biol. Sci., 14: 257-272.
CrossRefDirect Link - Muralidharan, P., G. Balamurugan and V. Babu, 2009. Cerebroprotective effect of Glycyrrhiza glabra Linn. root extract on hypoxic rats. Bangladesh J. Pharmacol., 4: 60-64.
Direct Link - Pari, L. and M. Latha, 2004. Protective role of Scoparia dulcis plant extract on brain antioxidant status and lipidperoxidation in STZ diabetic male Wistar rats. BMC Complementary Altern. Med., Vol. 4.
CrossRefDirect Link - Pietropaolo, S., Y. Sun, R. Li, C. Brana, J. Feldon and B.K. Yee, 2008. The impact of voluntary exercise on mental health in rodents: A neuroplasticity perspective. Behav. Brain Res., 192: 42-60.
CrossRefPubMedDirect Link - Polidori, M.C., 2003. Antioxidant micronutrients in the prevention of age-related diseases. J. Postgrad Med., 49: 229-235.
Direct Link - Jaiswal, P., P. Kumar, V.K. Singh and D.K. Singh, 2011. Areca catechu L.: A valuable herbal medicine against different health problems. Res. J. Med. Plant, 5: 145-152.
CrossRefDirect Link - Ravishankara, M.N., N. Shrivastava, H. Padh and M. Rajani, 2002. Evaluation of antioxidant properties of root bark of Hemidesmus indicus R. Br. (Anantmul). Phytomedicine, 9: 153-160.
CrossRefPubMedDirect Link - Resende, R., P.I. Moreira, T. Proenca, A. Deshpande, J. Busciglio, C. Pereira and C.R. Oliveira, 2008. Brain oxidative stress in a triple-transgenic mouse model of Alzheimer disease. Free Radical Biol. Med., 44: 2051-2057.
CrossRefPubMedDirect Link - Saitoh, A., M. Yamada, M. Yamada, S. Kobayashi, N. Hirose, K. Honda and J. Kamei, 2006. ROCK inhibition produces anxiety-related behaviors in mice. Psychopharmacology, 188: 1-11.
CrossRefPubMedDirect Link - Sapakal, V.D., T.S. Shikalgar, R.V. Ghadge, R.S. Adnaik, N.S. Naikwade and C.S. Magdum, 2008. In vivo screening of antioxidant profiles: A review. J. Herbal Med. Toxicol., 2: 1-8.
Direct Link - Sato, A., Y. Sato and S. Uchida, 2002. Regulation of cerebral cortical blood flow by the basal forebrain cholinergic fibers and aging. Auton. Neurosci., 96: 13-19.
Direct Link - Schlumpf, M., W. Lichtensteiger, H. Langemann, P.G. Waser and F. Hefti, 1974. A fluorometric micromethod for the simultaneous determination of serotonin, noradrenaline and dopamine in milligram amounts of brain tissue. Biochem. Pharmacol., 23: 2437-2446.
CrossRefPubMedDirect Link - Shanmugarajan, T.S., M. Arunsundar, I. Somasundaram, E. Krishnakumar, D. Sivaraman and V. Ravichandiran, 2008. Cardioprotective effect of Ficus hispida Linn. on cyclophosphamide provoked oxidative myocardial injury in a rat model. Int. J. Pharmacol., 4: 78-87.
Direct Link - Saeed, S.A., M.Z.S. Urfy, T.M. Ali, F.W. Khimani and A.U.H. Gilani, 2005. Antioxidants: Their role in health and disease. Int. J. Pharmacol., 1: 226-233.
CrossRefDirect Link - Saeed, S.A. and M.A. Mesaik, 2005. New vistas in the therapeutic uses of stem cells. J. Med. Sci., 5: 350-357.
CrossRefDirect Link - Takanaga, A., T. Hayakawa, K. Tanaka, K. Kawabata, S. Maeda and M. Seki, 2003. Immunohistochemical characterization of cardiac vagal preganglionic neurons in the rat. Auton. Neurosci., 106: 132-137.
PubMed - Vinutha, B., D. Prashanth, K. Salma, S.L. Sreeja and D. Pratiti et al., 2007. Screening of selected Indian medicinal plants for acetylcholinesterase inhibitory activity. J. Ethnopharmacol., 109: 359-363.
CrossRef - Winston, D., 2005. An introduction to herbal medicine. Am. Extra Pharmacopoeia, 2005: 1-8.
Direct Link - Yamada, M., T. Chiba, J. Sasabea, M. Nawa and H. Tajima et al., 2005. Implanted cannula-mediated repetitive administration of Aβ25-35 into the mouse cerebral ventricle effectively impairs spatial working memory. Behav. Brain Res., 164: 139-146.
CrossRef - Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain in conscious animals. Pain, 16: 109-110.
CrossRefPubMedDirect Link - Peraza-Sanchez, S.R., H. Chai, Y.G. Shin, T. Santisuk and V. Reutrakul et al., 2002. Constituents of the leaves and twigs of Ficus hispida. Planta Med., 68: 186-188.
CrossRefPubMedDirect Link