
J. Boateng
Nutritional Biochemistry and Carcinogenesis laboratory, Alabama A and M University, Normal, AL, USA
M. Verghese
Nutritional Biochemistry and Carcinogenesis laboratory, Alabama A and M University, Normal, AL, USA
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 3 | Page No.: 152-160







ABSTRACT
Consumption of fruits and the other dietary antioxidants are considered beneficial due to the protection they afford in the pathogenesis associated with oxidative stress. The aim of this study was to evaluate the antioxidative effects of selected fruit extracts (Plums, Apples, Grapes and Cranberries) on human lung fibroblasts (CCD-25LU) exposed to tert-butyl hydroperoxide (tBHP) oxidative stress. Lactate Dehydrogenase (LDH) was used to assess cytotoxicity (cell integrity) and antioxidant enzymes catalase (CAT), glutathione-s-transferase (GST), glutathione peroxidases (GPx) and concentrations of reduced glutathione (GSH) were determined. Results showed that LDH release by cells pretreated with fruits extracts were significantly (p<0.05) lower compared to cells treated with tBHP alone. Antioxidant enzymes (CAT, GST and GPx) in cells pretreated with fruit extracts were increased by 2-4 folds compared to cell exposed to tBHP alone. GSH levels which were significantly (p<0.05) reduced after exposure to tBHP were restored by pretreatment with fruit extracts. Fruits extracts used in this study protected CCD-25LU against oxidative stress induced by tBHP and reduced cell damage. Consumption of fruits may therefore play a significant role in protection against oxidative induced lung diseases.
PDF Abstract XML References Citation
Received: August 31, 2011;
Accepted: February 18, 2012;
Published: March 26, 2012
How to cite this article
J. Boateng and M. Verghese, 2012. Protective Effects of the Phenolic Extracts of Fruits against Oxidative Stress in Human Lung Cells. International Journal of Pharmacology, 8: 152-160.
DOI: 10.3923/ijp.2012.152.160
URL: https://scialert.net/abstract/?doi=ijp.2012.152.160
DOI: 10.3923/ijp.2012.152.160
URL: https://scialert.net/abstract/?doi=ijp.2012.152.160
INTRODUCTION
The lungs participate in several critical functions including respiration as well as serving as a physical barrier for the heart (Piao et al., 2008). Because they are typically exposed to higher oxygen tension than other organs, this makes them highly susceptible to oxidative damage due to the overproduction of ROS (Kinnula and Crapo, 2003; Bargagli et al., 2009; Ciencewicki et al., 2008). Many external factors including; cigarette smoke, drugs, radiation and nanoparticles have been reported to increase the production of ROS and in many instances are implicated in several lung pathologies (Ramesh et al., 2007; Ravichandran and Ramanibai, 2008). For example, Idiopathic Pulmonary Fibrosis (IPF), Emphysema/chronic obstructive pulmonary disease (COPD), cystic fibrosis and asthma (Kinnula et al., 2005; Bargagli et al., 2009; Ciencewicki et al., 2008). Furthermore, studies have revealed that these diseases including smoking are a prelude to lung cancer (Mayne et al., 1999). This is quite significant since more people die from lung cancer than from the combined deaths caused by breast, colorectal, prostate and pancreatic cancers (Venugopal et al., 2007; CDC and NCI, 2010).
In a review Kinnula and Crapo (2003), stressed that the lung possess a highly specialized and compartmentalized antioxidant defense system. These include low molecular weight thiol compounds such glutathione, superoxide dismutase (SOD), catalase (CAT) and detoxification enzyme systems such as glutathione-S-transferases. However, problem arises due to an imbalance between pro-oxidants and antioxidants resulting in oxidative stress (Prasanna and Purnima, 2011). Hence, the strategy to prevent this imbalance is to utilize products with antioxidant capabilities that act as free radical scavengers (Oboh, 2008). And more importantly, are capable of replenishing these endogenous enzymes.
It well known that fruits and vegetables provide protection against several diseases. Evidence supporting this has been extensively documented in both epidemiological and laboratory research findings (Potter, 1997; McCarty, 2004). The property thought to provide the protection afforded by fruits is through the actions of flavonoids and their ability to act as antioxidants (Eastwood, 1999; Cedenas and Packer, 1996; Liu, 2003). Flavonoids are widely distributed in dietary plants including fruits, vegetables, teas and wine (Gupta et al., 2010). They exhibit a wide range of biological activities including the scavenging of free radicals, chelating metal ions and inhibiting lipoprotein oxidation, among others (Liu, 2003; Raihan et al., 2009; Gupta et al., 2010; Renukadevi and Suhani Sultana, 2011). In light of these beneficial effects, adequate supply of antioxidants from the diet might help prevent or delay the occurrence of pathological changes that are associated with oxidative stress.
In this study, the organic hydroperoxide tert-butyl hydroperoxide (tBHP) was utilized. It has served as a useful model to investigate the mechanism of cell injury through its ability as an initiator of oxidative stress (Latour et al., 1995; Kennedy et al., 1992; Aherne and O’Brien, 2000; Hix et al., 2000; Lazze et al., 2003). It is metabolized to free radical intermediates through reactions by cytochrome P450 enzymes to yield tert-butoxy and methyl radicals (Latour et al., 1995; Kennedy et al., 1992; Lazze et al., 2003). Consequently, these radicals have been reported to deplete glutathione and protein thiols (Aherne and O’Brien, 2000; Hix et al., 2000) thus leading to oxidative stress.
It has been established that the lung is susceptible to the generation of reactive oxygen species. It is possible that the antioxidant action of polyphenolic compounds from fruits may attenuate these actions and decrease the susceptibility of the lung to oxidative challenges. Thus, the aim of this study was to assess the effects of phenolic extracts from fruits (grapes, apples, cranberries which are commonly consumed in the United States) in enhancing endogenous antioxidant enzyme activities in human lung cells. The cytoprotective properties of the fruit extracts after exposure to tert-butyl hydroperoxide was also evaluated.
MATERIALS AND METHODS
This study was carried out at Alabama A and M University from 2008-2010. Fresh fruits were purchased at commercial maturity from a local farmers’ market. Phytochemicals standards were purchased from Indofine (Hillsborough, NJ). (Lactate dehydrogenase) kit was purchased from Roche Applied Science (Indianapolis, IN). Eagle’s Minimum Essential Medium (EMEM), non-essential amino acids, sodium pyruvate, Fetal Bovine Serum (FBS) and antibiotics were all obtained from American type tissue culture (ATTC, Mannasas, VA). All other reagents and chemicals were of analytical grade and purchased from Fisher Scientific Company (Suwannee, GA).
Preparation of phenolic extraction from fruits: Phenolics from fresh fruits were extracted with 80% aqueous methanol (methanol: water, 80:20 v/v) (1:10) in a homogenizer. Phenolic compounds were extracted under restricted light and the mixture was centrifuged @ 1500 RPM/20 min/4°C. The residue was further extracted under the same conditions. The two filtrates were combined and transferred to an evaporating flask. The solvent was removed using a rotary evaporator at 40°C under restricted light and the phenolic concentrate was diluted to an appropriate volume. The phenolic extracts were dried and flushed with nitrogen gas and stored at -80°C until use.
Determination of total phenolics: Total phenolics were analyzed by Folin-Ciocalteu colorimetric method (Singleton et al., 1999) as previously described by (Kim et al., 2003). The absorbance was read at 750 nm and results from 3 separate experiments were expressed as mg gallic acid equivalents per 100 grams fresh fruit (mg gallic acid 100 g-1 fresh).
Determination of total flavonoids: Total flavonoids were analyzed by a colorimetric method previously reported by Sun and Liu (2006). The absorbance was read at 510 nm with a blank consisting of distilled water. Absorbance values were compared against catechin standard. Results from 3 separate experiments were expressed as mg catechin equivalents per 100 grams fresh fruit (mg catechin 100 g-1 fresh).
Cell culture: Human lung fibroblast cells (CCD-25LU) were obtained from American Type Tissue Culture (ATTC, Mannasas, VA) and cultured in Eagle’s Minimum Essential Medium (EMEM) supplemented with 10% fetal bovine serum (FBS), 0.1 mM non-essential amino acids and 1.0 mM sodium pyruvate with 100 U mL-1 penicillin and 100 μg mL-1 streptomycin. Cells were incubated at 37°C, 5% CO2 until confluent (80-90%).
Oxidative treatments: For experiments, cells were seeded in 12 well tissue culture plates at densities of 1.28x105 cells and incubated as previously specified until a monolayer developed. After the development of a monolayer, cells were washed twice with PBS and pretreated with 1 mL of fresh media (serum and pyruvate free) containing different concentrations (50-200 μg mL-1) fruit extracts. The cells were incubated for 24 and 48 h, the media alone (without serum) served as a negative control and for the positive control media containing tBHP was used. The experiment was repeated three times and samples were analyzed in triplicates. After the incubation period the medium was removed and the monolayer rinsed twice with cold PBS. To induce oxidative stress, serum free medium containing tBHP (300 μM) was applied to the cells and incubated for 3 h at 37°C. The medium was collected for lactate dehydrogenase (LDH) determination and the cells were harvested and used for the determination of antioxidative enzyme activities.
Determination of LDH: LDH release after incubation with tBHP was assayed after 3 min of incubation at room temperature at A-490 with reference wavelength at A-630 nm using a Biotek (Synergy) microplate reader (Winooski, Vermont).
LDH release was determined based on the following Eq:
Sample (treated cells) - Low control (untreated cells) High control (cells lysed with triton-x solution)-Low control x100 |
Preparation of cell lysate for enzyme analysis: Cells were lysed with lysis buffer (10 mM tris-HCL, pH 7.4, 1 mM EDTA, 0.15 M NaCl, 5 μg mL-1 aprotinin, 5 μg mL-1 leupeptin, 0.5 mM phenyl methyl sulfoxide (PMSF) 2 mM sodium orthovadate, 0.5% triton x-100) for 10 min at room temperature. The lysate was centrifuged at 14,000 xg for 20 min at 4°C. The supernatant which contained the cytosolic fraction was collected and used for enzyme analysis. Protein was measured using BCA protein assay kit from Pierce (Rockford, IL).
Enzymatic assays: CAT activity was detected at 240 nm by monitoring the decomposition of H2O2 as described by Johansson and Borg (1988). Glutathione peroxidase (GPx) activity was determined spectrophotometrically following the methods of Jaskot et al. (1983). Glutathione (GSH) was determined using the method of Griffith (1982). For Glutathione-S-tranferase (GST) activity, the method of Habig et al. (1974) was utilized.
Statistical analysis: Data were analyzed using SAS (2007) version 9.1 software. The data were expressed as Means±Standard error of mean (SEM) from means calculated from the means of triplicate replicates obtained in three separate or independent experiments. Wherever appropriate, the data were subjected to statistical analysis by one-way analysis of variance (ANOVA). For pairwise comparisons of means, Tukey test was applied, considering p<.05 statistically significant.
RESULTS
Total phenolics, flavonoids and total monomeric anthocyanins: Total phenolics, flavonoids and anthocyanins determined in the selected fruits (Apples, red cranberry (RC), red plums (RP) and red grapes (RG)) are shown in Table 1.
| Table 1: | Total Phenolics, total Flavonoid and total monomeric Anthocyanins contents in selected fruits |
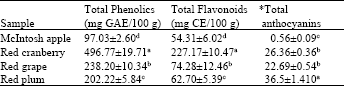 | |
| Data were expressed as Means±Standard Error of Mean (SEM) from means calculated from six replicates obtained in three separate or independent experiments, abcd Means within column with different letters are significantly different (p<0.05) using Tukey’s studentized range test, *(Cyanidin 3-glucoside equivalents mg 100 g-1), GAE: Gallic acid equivalents and CE: Catechin equivalents | |
Total phenolics (mg GAE 100 g-1 fresh weight) was significantly (p<0.05) higher in CB (496.77) compared to the other fruits, RG (238), RG (202.2) and APP (97.03). Total phenolics contents reported in RC was approximately two times higher than in red RP and RG and nearly five times higher than in APP. A similar trend was observed with total flavonoids content (mg CE 100 g-1 fresh weight) where RC showed highest flavonoids content (227.17) and the lowest flavonoids content established in APP (54.31). Sun and Liu (2006) reported total phenolic contents in CB at approximately 507 mg GAE 100 g-1, this value is similar to the phenolic content of CB (496.11 GAE 100 g-1) observed in the current study. Elsewhere, the levels of total phenolics and flavonoids in apples were reported as 118 mg GAE 100 g-1 and 62.0 mg CE 100 g-1, respectively (Kim et al., 2003). Despite having the highest total flavonoids and phenolics contents, total monomeric anthocyanidin content (Cyanidin 3-glucoside equivalents mg 100 g-1) was highest in RP when compared to CB, APP and RG.
Lactate dehydrogenase (LDH) activity in CCD-25LU cells: LDH release (%) in cells pretreated with fruit extracts was significantly (p<0.05) decreased compared to cells exposed to tBHP alone (Fig. 1). LDH release after 24 h pretreatment ranged from 18% (cells pretreated with RG and APP at 400 μg mL-1) to 50% (cells pretreated with 50 μg mL-1 CB). After 48 h pretreatment it ranged from 28% (cells pretreated with 400 μg mL-1 RG) to 61% (cells pretreated with 50 μg mL-1 CB). Comparing the incubation periods, LDH release was significantly (p<0.05) lowered in cells pretreated with fruit extracts for 24 h compared to 48 h. We noted approximately 12% to 38% increase in LDH release with prolonged incubation. Cells pretreated with APP and RG had a lower LDH release while those pretreated with CB and RP had a higher LDH release. Although these were significantly (p<0.05) lower compared to cells exposed to tBHP alone (79.56%).
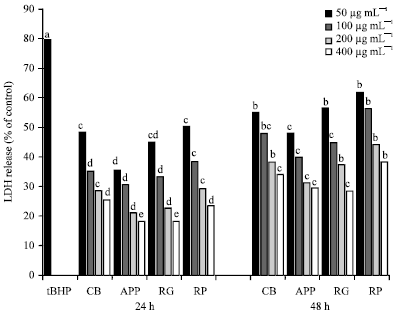 | |
| Fig. 1: | LDH release in CCD-25 LU cells pretreated with whole fruits extracts after exposure to tert-butyl hydroperoxide. Data were expressed as Means±Standard Error of Means (SEM) from means calculated from three replicates obtained in three separate or independent experiments. α-eMeans on a bar with different letters differ (p<0.05) using Tukey’s studentized range test, CV: Cranberries, APP: Apples (McIntosh apples), RG: Red grapes, RP: Red plums and tBHP: Tert butyl hydroperoxide |
| Table 2: | Glutathione-S-transferase activity (nmol mg-1 protein) in CCD-25LU cells pretreated with fruits extracts |
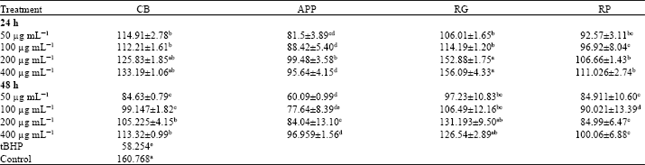 | |
| Data were expressed as Means±Standard Error of Mean (SEM) from three independent experiments. a-eMeans in a row with different letters differ (p<0.05) using Tukey’s studentized range test, CB: Cranberries, APP: Apples (McIntosh apples), RG: Red grapes, RP: Red plums and tBHP: tert butyl hydroperoxide | |
Also, it appeared that LDH release was decreased with increasing concentrations, thus indicating fruits extracts used in this study conferred protection against cytotoxicity in CCD25-LU cells.
Glutathione-S-transferase (GST) activity in CCD-25LU cells: Glutathione-S-transferase (GST) activity (nmol mg-1 protein) in cells pretreated with whole fruit extracts are shown in Table 2. We noted that GST activity was significantly (p<0.05) lower in cells pretreated with tBHP alone (58.25) compared to cells pretreated with fruit extracts after 24 and 48 h. GST was induced by approximately 31 to 62% in cells pretreated with fruit extracts for 24 and 4 to 51% after 48 h pretreatment. After 24 h it was observed that GST activity was significantly (p<0.05) increased compared to prolonged pretreatment (48 h). In CCD25-LU cells pretreated with CB and RG; GST activity was significantly (p<0.05) increased at all tested concentrations (50 and 400 μg mL-1), compared to cells pretreated with APP and RP. At 200 and 400 μg mL-1 GST activity in cells pretreated with RG for both incubation periods (24 and 48 h) was found to be statistically similar to control untreated cells. A similar trend was also observed with CB but only after 24 h pretreatment. It was also observed that the induction of GST activity was increased with increasing concentration regardless of incubation times.
| Table 3: | Glutathione peroxidase (GPx) activity (nmol/mg protein) in CCD-25LU cells pretreated with fruits extracts |
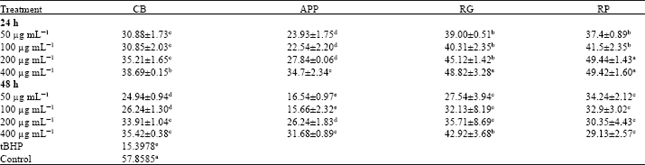 | |
| Data were expressed as Means±standard Error of Mean (SEM) from three replicates obtained in three separate or independent experiments, a-eMeans in a row with different letters differ (p<0.05) using Tukey’s studentized range test, CB: Cranberries, APP: Apples (McIntosh apples), RG: Red grapes, RP: Red plums and tBHP: Tert butyl hydroperoxide | |
| Table 4: | Glutathione (GSH) levels in (nmol/mg protein) CCD-25LU cells pretreated with pretreated with fruits extracts |
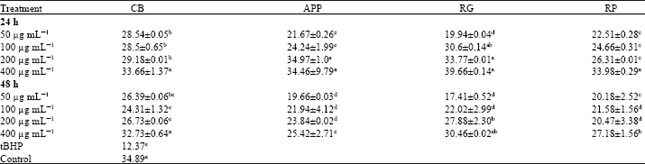 | |
| Data were expressed as Means±Standard Error of Mean (SEM) from three replicates obtained in three separate or independent experiments, a-eMeans in a row with different letters differ (p<0.05) using Tukey’s studentized range test, CB: Cranberries, APP: Apples (McIntosh apples), RG: Red grapes, RP: Red plums and tBHP: Tert butyl hydroperoxide | |
Glutathione peroxidase (GPx) activity in CCD-25LU cells: Table 3 shows glutathione peroxidase (GPx) activities (nmol mg-1 protein) in CCD-25LU cells, following pretreatment with fruit extracts. Pretreatment of cells with fruit extract for 24 and 48 h showed significant (p<0.05) increase in GPx activity compared to cells exposed to tBHP alone (15.39). Enzyme activity was increased by approximately 1-3 folds and 1-2 folds after 24 and 48 h pretreatment, respectively; except in cells pretreated with 50 and 100 μg mL-1 APP for 48 h. Here, data showed no significant differences compared to cells exposed to tBHP alone. In cells pretreated with RG (400 μg mL-1) and RP (200 and 400 μg mL-1) extracts, GPx activity was comparable to the control untreated cells, indicating a distinct protection by these fruit extracts. As previously observed with GST activities, GPx increased after 24 h pretreatment as compared to 48 h, GPx activity was increased by at least 4 to 31%. It was also observed that GPx activities increased with increasing concentrations after 24 and 48 h incubation.
Glutathione (GSH) levels in CCD-25LU cells: Glutathione GSH (nmol mg-1 protein) levels in CCD-25LU cells exposed to tBHP alone was severely depleted (12.37) and significantly (p<0.05) lower compared to cells pretreated with fruit extracts (Table 4). After 24 h pretreatment with fruit extracts, GSH levels was approximately 2-3 fold higher than in cells exposed to tBHP alone. GSH levels were comparable to or higher than in the control untreated cells in cells pretreated with APP and RG (both at 200 and 400 μg mL-1) and CB and RP(both at 400 μg mL-1). GSH levels slightly decreased after 48 h pretreatment with fruit extracts at all concentrations tested. Compared to 24 h, GSH level decreased by 7% in CB treated cells (50 μg mL-1) to 31% in cells treated with APP extracts (200 μg mL-1).
Catalase (CAT) activity in CCD-25LU cells: CAT (μmol mg-1 protein) activity (Table 5) in cells pretreated with whole fruit extracts was significantly (p<0.05) increased compared to cells exposed to tBHP alone. However, it was comparable to or higher than the control untreated cells. After 24 h, cells pretreated with RG extract had the highest CAT activity at all concentration tested, followed by cells pretreated with CB, RP and APP extracts. Compared to the cells exposed to tBHP alone, CAT activity increased by approximately 56 to 60% in cells pretreated with RG extracts. In cells pretreated with CB, APP and RP extracts the increase was 46 to 53%, 25 to 50 and 40 to 47%, respectively.
| Table 5: | Catalase (CAT) activity (μmol mg-1 protein) in CCD-25LU cells pretreated with whole fruit extracts |
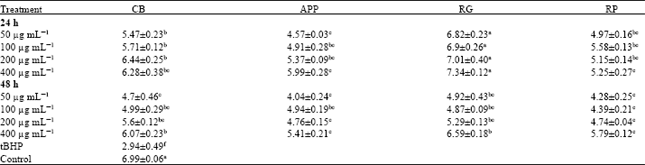 | |
| Data were expressed as Means±Standard Error of Mean (SEM) from three replicates obtained in three separate or independent experiments, a-eMeans in a row with different letters differ (p<0.05) using Tukey’s studentized range test, CB: Cranberries, APP: Apples (McIntosh apples), RG: Red grapes, RP: Red plums and tBHP: Tert butyl hydroperoxides | |
After 48 h pretreatment with fruit extracts, CAT activity slightly decreased by nearly 3 to 29% compared to 24 h pretreatment. However, compared to cells exposed to tBHP alone, it increased by 37 to 51% in cells pretreated with CB extract. There was a 27 to 45% increase in cells pretreated with APP extracts. In the other treatment groups CAT was increased by 39 to 55% and 31 to 49% in cells pretreated with RG and RP extracts, respectively.
DISCUSSION
This study demonstrated that the phenolic extracts from apples, cranberries and grapes have the potential of protecting human lung cells (CCD-25LU) against tBHP-induced oxidative stress, by modulating endogenous enzymatic and non enzymatic antioxidants. LDH is a crucial biomarker for assessing cell viability of proliferation (Gupta et al., 2011). Using LDH as a marker for cell viability and membrane integrity, this study revealed that pretreatment of CCD-25LU with fruit extracts resulted in a dramatic decrease in LDH. These are in line with studies where similar observations were noted (Garcia-Alonso et al., 2007; Lima et al., 2006; Liu et al., 2010). The decrease in the loss of cell viability by the fruit extracts may indicate a reduction in the production of ROS. For example, increased cytosolic Ca+ levels can lead to the production of ROS which is likely to cause increase in peroxidation of membrane phospholipids. The peroxidation of membrane phospholipids results in increase of membrane fluidity which eventually leads to disruption of membrane structure and function (Sohn et al., 2005; Alia et al., 2005). In this case, we propose that the flavonoids compounds in the fruit extracts may have afforded protection to the cells by acting as antioxidants.
GSH is a major intracellular thiol of redox system in the body that protects the cells against oxidative damage (Nishigaki et al., 2007). Although GSH levels were significantly decreased in CCD-25LU cells exposed to tBHP alone, pretreatment with fruit extracts prevented its depletion. In their study (Alia et al., 2005; Martin et al., 2008) reported similar effects with dietary antioxidants in HepG2 cells. One of the primary consequences ascribed to GSH depletion is the induction of mitochondrial permeability transition (MPT). This leads to loss of Mitochondrial Membrane Potential (MMP) which is crucial in maintaining intracellular GSH homeostasis (Tripathi et al., 2009). The inability to effectively maintain intracellular homeostasis leads to loss of cell viability which was detected when CCD-25LU was exposed to tBHP alone. In terms of maintaining GSH levels, it can be suggested that fruits extracts, in preventing the collapse of MMP, adds to their cytoprotective role. To expatiate further, persons with lung fibrosis have been found to have diminished levels of GSH (Liu and Pravia, 2010) which for all practical purposes, results in damage to macromolecules. And as mentioned earlier, oxidative induced diseases of the lung can eventually lead to cancer. Therefore, the protection afforded by the fruit extracts, indicate its ability to potentially prevent oxidative related lung diseases.
CAT and GPx are key players in protecting cells from oxidative stress (Vandana et al., 2006). CAT is known for catalyzing the dismutation of hydrogen peroxide (H2O2) into water (H2O) and molecular oxygen (O2) and unlike GPx requires no cofactors in its catalytic mode. Under our experimental conditions pretreatment with fruit extracts enhanced GPx and catalase (CAT) activities in CCD-25LU cells. Others have reported similar results using Chinese hamster lung fibroblasts (V79-4) cells (Kang et al., 2005, 2006; Piao et al., 2008). In the current study CAT activity was higher than GPx activity. Verkerk and Jongkind (1992) noted this to be the case during acute oxidative stress. GPx requires GSH as a cofactor to exert its function. Therefore, the depletion of GSH can reduce the activity of GPx and consequently limit the activity of the enzyme. This was observed in CCD-25LU cells exposed to tBHP. Once more, the increased activities of CAT and GPx could most likely be due to the ability of whole fruits extracts to maintain GSH levels during peroxide-induced toxicity. In any case, these results suggest that fruit extracts developed an endogenous protection to CCD-25LU.
The principle of Phase II detoxification enzyme GST catalyzes the conjugation of GSH to yield hydrophobic molecules. Hence, it participates in reactions that destroy hydrogen peroxide and organic peroxides (Garcia-Alonso et al., 2007). For that reason, increased GST activity in CCD-25LU cells after pretreatment with fruit extracts add to the antioxidative properties of these extracts. Indeed, total phenolics and flavonoids contents in the fruits utilized may have contributed significantly to this property; i.e., ability to scavenge free radicals. Present findings are in accordance with Alia et al. (2005); (Garcia-Alonso et al., 2007) who reported enhanced GST activities in HepG2 after pretreatments with phenolic juice extracts. In CCD-25LU cells exposed to tBHP, we noted a decline in the enzyme activity. During their study Shanmugarajan and Devaki (2008) also reported reduced GST activity in liver of mice administered Azathioprine (AZA). In concurrence, it was suggested that the decrease in GSH levels might have contributed to decreased enzyme activity. On the other hand, the elevated cellular GSH content may have contributed to an increase GST activity observed in cells pretreated with fruit extracts. It has been shown that phytochemicals which are inducers of phase II enzymes, may also have cancer preventing properties (Kensler et al., 1993; Boateng et al., 2007).
In the current study we noted decreased enzymatic activities with prolonged pretreatment of CCD-25LU with fruit extracts. However, the fact that fruits extract had a stronger capability to ameliorate tBHP-induced oxidative stress during short preincubation could be due to the following; 1: As previously noted, the optimum time and dose to provide protection in our cell system seem to be 24 h prior to exposure to tBHP, 2: Short exposures to antioxidants induced a longer cytoprotective response through the induction of antioxidant enzyme than at higher concentrations and for longer incubation periods (Rubiolo et al., 2008). Even so, GSH, GPx, CAT and GST activities were significantly enhanced compared to cells exposed to tBHP alone.
CONCLUSION
Our goal in this study was to highlight the protective effects of phenolic rich fruits extract on the modulation of endogenous antioxidants and cell toxicity in human lung fibroblast (CCD-25LU) against tBHP-induced. To our knowledge, these results are the first to demonstrate an antioxidative and cytoprotective role of fruits (apples, cranberries and grapes) against normal human lung cells. While results are preliminary, fruits utilized in this study could be considered as chemo preventive agents against lung cancer. Since lung cancer is the leading cause of cancer death in men and women, these results could provide data to further investigate an in depth study on the protective properties of fruits against ROS related lung diseases.
ACKNOWLEDGMENT
Funding for this research was made available by U.S. Department of Defense (NATICK).
REFERENCES
- Aherne, S.A. and N.M. O'Brien, 2000. Mechanism of protection by the flavonoids, quercetin and rutin, against tert-butylhydroperoxide- and menadione-induced DNA single strand breaks in Caco-2 cells. Free Radical Biol. Med., 29: 507-514.
CrossRefPubMedDirect Link - Alia, M., S. Ramos, R. Mateos, L. Bravo and L. Goya, 2005. Response of the antioxidant defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2). J. Biochem. Mol. Toxicol., 19: 119-128.
CrossRefDirect Link - Boateng, J., M. Verghese, L. Shackelford, L.T. Walker and J. Khatiwada et al., 2007. Selected fruits reduce azoxymethane (AOM)-induced aberrant crypt foci (ACF) in fisher 344 male rats. Food Chem. Toxicol., 45: 725-732.
CrossRefPubMedDirect Link - Eastwood, M.A., 1999. Interaction of dietary antioxidants in vivo: How fruit and vegetables prevent disease? Q. J. Med., 92: 527-530.
PubMedDirect Link - Garcia-Alonso, F.J., A. Guidarelli and M.J. Periago, 2007. Phenolic-rich juice prevents DNA single-strand breakage and cytotoxicity caused by tert-butylhydroperoxide in U937 cells: The role of iron chelation. J. Nutr. Biochem., 18: 457-466.
CrossRefDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Hix, S., M.B. Kadiiska, R.P. Mason and O. Augusto, 2000. In vivo metabolism of tert-butyl hydroperoxide to methyl radicals. EPR spin-trapping and DNA methylation studies. Chem. Res. Toxicol., 13: 1056-1064.
CrossRefDirect Link - Johansson, L.H. and L.A. Borg, 1988. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem., 174: 331-336.
CrossRefPubMedDirect Link - Kang, K.A., K.H. Lee, S. Chae, R. Zhang and M.S. Jung et al., 2006. Cytoprotective effect of phloroglucinol on oxidative stress induced cell damage via catalase activation. J. Cell. Biochem., 97: 609-620.
CrossRefPubMedDirect Link - Kang, K.A., K.H. Lee, S. Chae, R. Zhang and M.S. Jung et al., 2005. Cytoprotective effect of tectorigenin, a metabolite formed by transformation of tectoridin by intestinal microflora, on oxidative stress induced by hydrogen peroxide. Eur. J. Pharmacol., 519: 16-23.
CrossRefDirect Link - Kennedy, C.H., D.F. Church, G.W. Winston and W.A. Pryor, 1992. tert-Butyl hydroperoxide-induced radical production in rat liver mitochondria. Free Radical Biol. Med., 12: 381-387.
PubMedDirect Link - Kensler, T.W., N.E. Davidson, J.D. Groopman, B.D. Roebuck, H.J. Prochaska and P. Talalay, 1993. Chemoprotection by inducers of electrophile detoxication enzymes. Basic Life Sci., 61: 127-136.
PubMed - Kinnula, V.L. and J.D. Crapo, 2003. Superoxide dismutases in the lung and human lung diseases. Am. J. Respir. Crit. Care Med., 167: 1600-1619.
PubMedDirect Link - Latour, I., J.B. Demoulin and P. Buc-Calderon, 1995. Oxidative DNA damage by t-butyl hydroperoxide causes DNA single strand breaks which is not linked to cell lysis. A mechanistic study in freshly isolated rat hepatocytes. FEBS Lett., 373: 299-302.
CrossRefPubMedDirect Link - Lazze, M.C., R. Pizzala, M. Savio, L.A. Stivala, E. Prosperi and L. Bianchi, 2003. Anthocyanins protect against DNA damage induced by tert-butyl-hydroperoxide in rat smooth muscle and hepatoma cells. Mutat. Res./Genet. Toxicol. Environ. Mutagen., 535: 103-115.
CrossRefDirect Link - Lima, C.F., M. Fernandes-Ferreira and C. Pereira-Wilson, 2006. Phenolic compounds protect HepG2 cells from oxidative damage: Relevance of glutathione levels. Life Sci., 79: 2056-2068.
CrossRefPubMedDirect Link - Liu, R.H., 2003. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr., 78: 517S-520S.
CrossRefPubMedDirect Link - Liu, S., W. Hou, P. Yao, B. Zhang, S. Sun, A.K. Nussler and L. Liu, 2010. Quercetin protects against ethanol-induced oxidative damage in rat primary hepatocytes. Toxicol. In vitro, 24: 516-522.
CrossRefPubMedDirect Link - Liu, R.M. and K.A.G. Pravia, 2010. Oxidative stress and glutathione in TGF-beta-mediated fibrogenesis. Free Radic Biol. Med., 48: 1-15.
PubMed - Martin, M.A., S. Ramos, R. Mateos, A.B.G. Serrano, M. Izquierdo-Pulido, L. Bravo and L. Goya, 2008. Protection of human HepG2 cells against oxidative stress by cocoa phenolic extract. J. Agric. Food Chem., 56: 7765-7772.
CrossRefPubMedDirect Link - Mayne, S.T., J. Buenconsejo and D.T. Janerich, 1999. Previous lung disease and risk of lung cancer among men and women nonsmokers. Am. J. Epidemiol., 149: 13-20.
PubMed - McCarty, M.F., 2004. Proposal for a dietary phytochemical index. Med. Hypotheses, 63: 813-817.
CrossRefDirect Link - Piao, M.J., K.A. Kang, R. Zhang, D.O. Ko and Z.H. Wang et al., 2008. Hyperoside prevents oxidative damage induced by hydrogen peroxide in lung fibroblast cells via an antioxidant effect. Biochim. Biophys. Acta, 1780: 1448-1457.
CrossRef - Potter, J.D., 1997. Cancer prevention: Epidemiology and experiment. Cancer Lett., 114: 7-9.
CrossRefDirect Link - Raihan, S.Z., A.K. Chowdhury, G.H. Rabbani, F. Marni, M.S. Ali, L. Nahar and S.D. Sarker, 2009. Effect of aqueous extracts of black and green teas in arsenic-induced toxicity in rabbits. Phytother. Res., 23: 1603-1608.
CrossRefPubMedDirect Link - Rubiolo, J.A., G. Mithieux and F.V. Vega, 2008. Resveratrol protects primary rat hepatocytes against oxidative stress damage: Activation of the Nrf2 transcription factor and augmented activities of antioxidant enzymes. Eur. J. Pharmacol., 591: 66-72.
CrossRef - Singleton, V.L., R. Orthofer and R.M. Lamuela-Raventos, 1999. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In: Methods in Enzymology, Burslem, G.L. (Ed.), Academic Press, Cambridge, Massachusetts, ISBN: 9780121822002, pp: 152-178.
CrossRefDirect Link - Sohn, J.H., K.L. Han, S.H. Lee and J.K. Hwang, 2005. Protective effects of panduratin A against oxidative damage of tert-butylhydrop-eroxide in human HepG2 cells. Biol. Pharm. Bull., 28: 1083-1086.
PubMedDirect Link - Tripathi, M., B.K. Singh and P. Kakkar, 2009. Glycyrrhizic acid modulates t-BHP induced apoptosis in primary rat hepatocytes. Food Chem. Toxicol., 47: 339-347.
CrossRefDirect Link - Vandana, S., S. Ram, M. Ilavazhagan, G.D. Kumar and P.K. Banerjee, 2006. Comparative cytoprotective activity of vitamin C, E and beta-carotene against chromium induced oxidative stres in murine macrophages. Biomed. Pharmacother., 60: 71-76.
PubMedDirect Link - Bargagli, E., C. Olivieri, D. Bennett, A. Prasse, J. Muller-Quernheim and P. Rottoli, 2009. Oxidative stress in the pathogenesis of diffuse lung diseases: A review. Respir. Med., 103: 1245-1256.
CrossRefPubMedDirect Link - Kinnula, V.L., C.L. Fattman, R.J. Tan and T. Oury, 2005. Oxidative stress in pulmonary fibrosis: A possible role for redox modulatory therapy. Am. J. Respir. Crit. Care Med., 172: 417-422.
CrossRefDirect Link - Venugopal, R., P. Senthilnathan, V. Magesh, B. Rajkapoor and D. Sakthisekaran, 2007. Chemotherapeutic efficacy of Solanum Trilobatum along with paclitaxel in lung cancer bearing swiss albino mice. Int. J. Cancer Res., 3: 119-126.
Direct Link - Ramesh, T., R. Mahesh and V. Hazeena Begum, 2007. Effect of Sesbania grandiflora on lung antioxidant defense system in cigarette smoke exposed rats. Int. J. Biol. Chem., 1: 141-148.
CrossRef - Prasanna, G.S. and A. Purnima, 2011. Protective effect of leaf extract of Trichilia connaroides on hypercholesterolemia induced oxidative stress. Int. J. Pharmacol., 7: 106-112.
CrossRefDirect Link - Renukadevi, K.P. and S.S. Sultana, 2011. Determination of antibacterial, antioxidant and cytotoxicity effect of Indigofera tinctoria on lung cancer cell line NCI-h69. Int. J. Pharmacol., 7: 356-362.
CrossRefDirect Link - Ravichandran, K. and R. Ramanibai, 2008. Protective effect of flavanoidal fraction of Indigofera tinctoria on benzo (α) pyrene induced lung carcinogenicity in swiss albino mice. Int. J. Cancer Res., 4: 71-80.
CrossRefDirect Link - Kim, D.O., O.K. Chun, Y.J. Kim, H.Y. Moon and C.Y. Lee, 2003. Quantification of polyphenolics and their antioxidant capacity in fresh plums. J. Agric. Food Chem., 51: 6509-6515.
CrossRefDirect Link - Sun, J. and R.H. Liu, 2006. Cranberry phytochemical extracts induce cell cycle arrest and apoptosis in human MCF-7 breast cancer cells. Cancer Lett., 241: 124-134.
PubMedDirect Link - Nishigaki, I., B. Rajkapoor, R. Venugopal, P. Senthilnathan and D. Sakthisekaran, 2007. Protective effect of fresh Apple extract on human umbilical endothelial vein cells against glycated protein-iron chelate induced toxicity. Asian J. Cell Biol., 2: 34-42.
CrossRefDirect Link - Gupta, V.K., R. Kumria, M. Garg and M. Gupta, 2010. Recent updates on free radicals scavenging flavonoids: An overview. Asian J. Plant Sci., 9: 108-117.
CrossRefDirect Link - Gupta, A., S. Singh, F. Jamal, S. Nath, S. Mehrotra and B. Sharma, 2011. Synergistic effects of glutathione and vitamin E on ROS mediated ethanol toxicity in isolated rat hepatocytes. Asian J. Biochem., 6: 347-356.
CrossRefDirect Link - Shanmugarajan, T.S. and T. Devaki, 2008. Ficus hispida Linn. leaf extract possesses antioxidant potential and abrogates azathioprine induced prooxidant and antioxidant imbalance in rat liver. Int. J. Pharmacol., 4: 376-381.
CrossRefDirect Link - Verkerk, A. and J.F. Jongkind, 1992. Vascular cells under peroxide induced oxidative stress: A balance study on in vitro peroxide handling by vascular endothelial and smooth muscle cells. Free Radical Res. Commun., 17: 121-132.
PubMedDirect Link - Jaskot, R.H., E.G. Charlet, E.C. Grose, M.A. Grady and J.H. Roycroft, 1983. An automated analysis of glutathione peroxidase, S-transferase, and reductase activity in animal tissue. J. Anal. Toxicol., 7: 86-88.
PubMedDirect Link - Griffith, O.W., 1982. Mechanism of action, metabolism and toxicity of buthionine sulfoximine and its higher homologs, potent inhibitors of glutathione synthesis. J. Biol. Chem., 257: 13704-13712.
PubMedDirect Link - Oboh, G., 2008. Antioxidative potential of Ocimum gratissimum and Ocimum canum leaf polyphenols and protective effects on some pro-oxidants induced lipid peroxidation in rat brain: An in vitro study. Am. J. Food Technol., 3: 325-334.
CrossRefDirect Link