
M. Kadioglu
Department of Pharmacology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
E. Muci
Department of Pharmacology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
M. Kesim
Department of Pharmacology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
C. Ulku
Department of Pharmacology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
E. N. Duman
Department of Anesthesiology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
N. I. Kalyoncu
Department of Pharmacology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
E. Yaris
Department of Pharmacology, Faculty of Medicine, Karadeniz Technical University, Mine Kadioglu, TR-61187 Trabzon, Turkey
International Journal of Pharmacology
Year: 2011 | Volume: 7 | Issue: 2 | Page No.: 283-290







ABSTRACT
Selective serotonin re-uptake inhibitors are used in the treatment of many psychiatric diseases and also diabetic neuropathy. But treatment with these agents may directly interfere with blood glucose levels. Clinical and experimental trials have demonstrated that some of SSRIs have opposite effects on blood glucose levels. The aim of this study was to investigate the acute effect of paroxetine on glucose regulation. BALB/c mice of either sex weighting 20-40 g were used in the experiments. The animals with pre-treatment blood glucose between 60-160 mg dL-1 were divided into 2 (normoglycemic and glucose tolerance test) groups. The effect of three doses (5-10-20 mg kg-1) of paroxetine on blood glucose level and the mechanisms of this effect are studied. Paroxetine decreased blood glucose levels at almost all doses and times tested in normoglycemic mice and in the intraperitoneal glucose tolerance test. Similar to paroxetine, serotonin (20 mg kg-1) also decreased blood glucose levels significantly in both groups. Neither diazoxide (potassium channel opener, 25 mg kg-1) nor verapamil (calcium antagonist, 25 mg kg-1) could inhibit the hypoglycemic effect of paroxetine. But both methysergide (5-HT1/2B/2C antagonist, 1 mg kg-1) and ketanserin (5-HT2A antagonist, 1 mg kg-1) inhibited the decreasing effect of paroxetine and serotonin. In conclusion, paroxetine causes a decrease in blood glucose level probably by an increase in insulin secretion and this increase is provided possibly by intracellular mechanisms mediated by serotonergic receptors and postreceptor events, rather than an influx of calcium from the extracellular medium.
PDF Abstract XML References Citation
Received: December 04, 2010;
Accepted: March 30, 2011;
Published: May 05, 2011
How to cite this article
M. Kadioglu, E. Muci, M. Kesim, C. Ulku, E. N. Duman, N. I. Kalyoncu and E. Yaris, 2011. The Effect of Paroxetine, A Selective Serotonin Reuptake Inhibitor, on Blood Glucose Levels in Mice. International Journal of Pharmacology, 7: 283-290.
DOI: 10.3923/ijp.2011.283.290
URL: https://scialert.net/abstract/?doi=ijp.2011.283.290
DOI: 10.3923/ijp.2011.283.290
URL: https://scialert.net/abstract/?doi=ijp.2011.283.290
INTRODUCTION
Selective serotonin re-uptake inhibitors (SSRIs) are widely used in the treatment of many psychiatric diseases, such as depression and panic disorders and are also used in the treatment of peripheral neuropathy seen in diabetic patients. (Taliyan and Sharma, 2010). Hyperglycemia is an important factor in development and progress of diabetic peripheral neuropaty and also in controlling the pain in this disease (Taliyan and Sharma, 2010; Moghtaderi et al., 2006; Behnam-Rassouli et al., 2010, Mohamed et al., 2009). However, treatment with SSRIs may directly interfere with blood glucose levels or may interact with hypoglycemic agents and can present potential risks to diabetic patients. Clinical and experimental trials have demonstrated that some SSRIs have opposite effects on blood glucose levels (hypoglycemia or hyperglycemia) in different species. (Gomez et al., 2001a, b; Kesim et al., 2006; Pollak et al., 2001; Sansone and Sansone, 2003). The interaction of SSRIs with blood glucose levels may cause failure in the treatment of diabetes and have a negative effect on the life quality of these patients such as an increase in the frequency and severity of hypoglycemia and the absence of hypoglycemia symptoms. In particular, fluoxetine, sertraline and fluvoxamine, but not paroxetine, have been studied in this respect. (Gomez et al., 2001a; Pollak et al., 2001; Sansone and Sansone, 2003; Mahmood et al., 2010; Oswald et al., 2003).
The main factor in the regulation of blood glucose is insulin, which is secreted from the beta cells of pancreatic Langerhans islets (Velavan and Hazeena Begum, 2007). It is known that the coupling factor in insulin secretion from beta cells is calcium. Activation of glucoreceptors situated on the membranes of beta cells by glucose leads to calcium influx and membrane depolarization. This depolarization triggers a rapid exocytosis of granules in which insulin is stored and the amount of glucose entering into the cells increases. When glucose is metabolized, the amount of adenosine triphosphate (ATP) inside the cell also rises and interacts with the
ATP-dependent potassium channels. Closure of ATP-dependent potassium channels causes an increase in intracellular calcium via. the opening of voltage-dependent calcium channels situated in the membrane. This increase in calcium level is the main factor in the synthesis and secretion of insulin (Davis, 2006; Yang and Gills, 2004; Wasada, 2002; Straub and Sharp, 2002).
Serotonergic nerves in the pancreas have been demonstrated in several studies and serotonin (5-hydroxytryptamine, 5-HT) has been shown to participate in glucose regulation and insulin secretion (Adeghate et al., 1999; Sugimoto et al., 2003; Yamada et al., 1999). Peripheral 5-HT2A receptors are the main receptors responsible for the insulin secreting effect of 5-HT and are known to be coupled to phospholipase C present in the membrane of beta cells. The production of inositol triphosphate (IP3) by activation of phospholipase C causes a release of calcium from the intracellular stores. In addition activation of 5-HT2A receptors causes a slow depolarization followed by an increase in the amount of intracellular calcium (Adeghate et al., 1999; Sugimoto et al., 2003; Yamada et al., 1999).
The aim of this study was to investigate the acute effect of paroxetine, a SSRI which has not been previously tested in detail, on glucose regulation and the glucose tolerance test in normoglycemic.
MATERIALS AND METHODS
Animals: Study was done in the laboratories of Department of Pharmacology, Karadeniz Technical University, Faculty of Medicine. BALB/c mice (20-40 g) of either sex were used in the experiments. The animals were housed in cages with free access to food and water and were not starved before the experiments. The cages were placed in a quiet and temperature-humidity controlled
room (22±2°C and 60±5%, respectively) in which a 12:12 h light-dark cycle was maintained. Experiments were conducted between 9:00 and 17:00 h to minimize diurnal variation. The experimental protocol was approved by the Institutional Animal Ethics Committee of Karadeniz Technical University, Medical School (2004/10). Animals were allowed a 1-week acclimatization period before being used in experiments.
Drugs: Paroxetine hydrochloride (Novartis, Istanbul, Turkey), verapamil, serotonin, methysergide, ketanserin and glucose (Sigma) were dissolved in distilled water and diazoxide (Sigma) was dissolved in 0.1 M NaOH. All drugs were injected intraperitoneally in a dose volume of no more than 10 mL kg-1 of body weight.
Experimental protocols: Blood glucose levels were measured before the drug application procedure in all groups. Blood was collected by puncture of the distal end of the mouse tail and blood drops were placed on the test zone of the strip for immediate measurement of glycemia using a glucometer device (MediSense-Optium, Abbott, UK). Animals with a pre-treatment blood glucose level between 60-160 mg dL-1 (normoglycemic) were divided into 2 main groups. While in first group the effect of paroxetine on the blood glucose levels in 3 different doses (5, 10, 20 mg kg-1) and the possible mechanism were investigated, in the second group, an intraperitoneal glucose tolerance test (IGTT) was applied and the effect of paroxetine (5, 10, 20 mg kg-1) and serotonin (20 mg kg-1) on this test was investigated.
The experimental groups and the time intervals of blood glucose level measurements were shown in Table 1. The doses of the drugs were decided with literature review.
| Table 1: | Experimental groups: group 1 (normoglycemic mice), group 2 (intraperitoneal glucose tolerance test in normoglycemic mice) (In each subgroup number of animals were 8) |
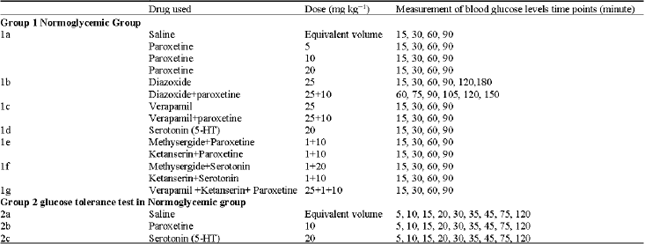 | |
Intraperitoneal glucose tolerance test (IGTT): Normoglycemic mice (group 2) were fasted overnight (12 h). A bolus of glucose (1 g kg-1 b.wt.) was administered by i.p. injection (Gletsu et al., 2005). Blood samples were taken before i.p. glucose injection and 5, 10, 15, 20, 30, 35, 45, 75 and 120 min after the injection as shown in Fig. 1. Thereafter, the mice were allowed free access to food and water.
Statistical analysis: The data were expressed as the Mean±SEM. Statistical analyses were carried out using the one way ANOVA test followed by the post hoc Bonferroni test when needed (Mohajeri et al., 2009; Chaudhari et al., 2010).
 | |
| Fig. 1: | Intraperitoneal glucose tolerance test (IGTT) |
The level of significance was set at p<0.05.
RESULTS
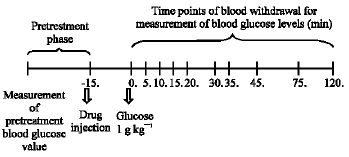
Paroxetine decreased blood glucose levels at almost all doses used and at all times tested in the first group of animals when compared with the pre-treatment blood glucose values and also when compared each corresponding value with saline group (Fig. 2a, p<0.006) and in the intraperitoneal glucose tolerance test (Fig. 2b, p<0.006).
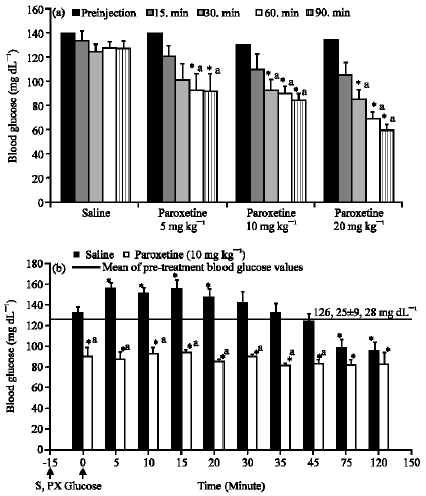
While diazoxide (an ATP-dependent potassium channel opener, 25 mg kg-1) increased blood glucose levels significantly at 60, 90 and 120. min when given alone (Fig. 3a), paroxetine at the 10 mg kg-1 dose given after diazoxide, reversed this increase and caused a decrease in blood glucose levels (Fig. 3b).
The hypoglycemic effect of paroxetine at the 10 mg kg-1 dose was not inhibited by application of the voltage-sensitive calcium channel antagonist verapamil (25 mg kg-1) (Fig. 4).
 | |
| Fig. 2: | The effect of paroxetine on blood glucose level in normoglycemic mice and in intraperitoneal glucose tolerance test (B). Results are expressed as Mean±SEM. *p<0.05, compared with the mean of pre-treatment blood glucose values, a: p<0.05, compared with saline group. S: Saline, PX: Paroxetine |
 | |
| Fig. 3: | The effect of diazoxide (a) on blood glucose level and the effect of paroxetine (PX) on the increase in blood glucose level induced by diazoxide when applied after diazoxide (b). Results are expressed as Means±SEM. *p<0.05, compared with the mean of pre- treatment blood glucose values |
Similar to paroxetine, serotonin 20 mg kg-1 also decreased blood glucose levels significantly in normoglycemic mice (Fig. 5a, p<0.005). In the intraperitoneal glucose tolerance test applied to animals in the second group, the increase in blood glucose levels due to glucose administration was completely prevented both by paroxetine at the 10 mg kg-1 dose and serotonin 20 mg kg-1 dose. Paroxetine and serotonin also decreased blood glucose levels to below pre-treatment blood glucose values when compared to both pre-treatment blood glucose levels and blood glucose levels in the saline group (Fig. 5b).
Both methysergide (a 5-HT1/2B/2C receptor antagonist, 1 mg kg-1) and ketanserin (a 5-HT2A receptor antagonist, 1 mg kg-1) inhibited the decreasing effect of paroxetine on blood glucose levels (Fig. 6a). Similar to paroxetine, the hypoglycemic effect of serotonin was also inhibited by these two antagonists (Fig. 6b).
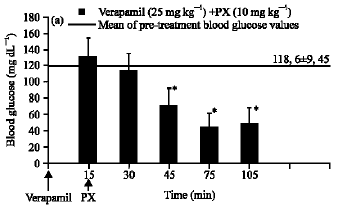 | |
| Fig. 4: | The effect of paroxetine (PX) on blood glucose level in the presence of verapamil. Results are expressed as Means±SEM. *p<0.05, compared with the mean of pre- treatment blood glucose values |
While inhibition of voltage-sensitive calcium channels via verapamil did not prevent hypoglycemia induced by paroxetine (Fig. 4), ketanserin used in combination with verapamil reversed this hypoglycemia (Fig. 7).
DISCUSSION
In our study, paroxetine caused a decrease in blood glucose levels in normoglycemic mice. There have been no previous investigations evaluating the direct effect of paroxetine on blood glucose in humans or animals. Some indirect results have been obtained from previous studies which suggested increased insulin sensitivity in humans (Weber-Hamann et al., 2006), hyperglycemia observed in a woman with panhypopituitarism (Petty, 1996) and increased glucose metabolism in the brain in depressed patients (Kennedy et al., 2001) and hypoglycemia in mice (Kesim et al., 2006).
The main mechanism involved in decreasing blood glucose levels is the secretion of insulin from pancreatic beta cells. Glucose is the main stimulant triggering this secretion (Davis, 2006; Yang and Gills, 2004). A method of testing the secretion of insulin induced by glucose in clinical practice is the glucose tolerance test. While paroxetine caused a decrease in blood glucose levels, it also altered the pattern of the glucose tolerance test by preventing the expected increase due to glucose. In additional, paroxetine decreased blood glucose levels to below pre-treatment values. The decrease in glucose level in spite of the overload of glucose may be related to a possible increase in insulin secretion caused by paroxetine.
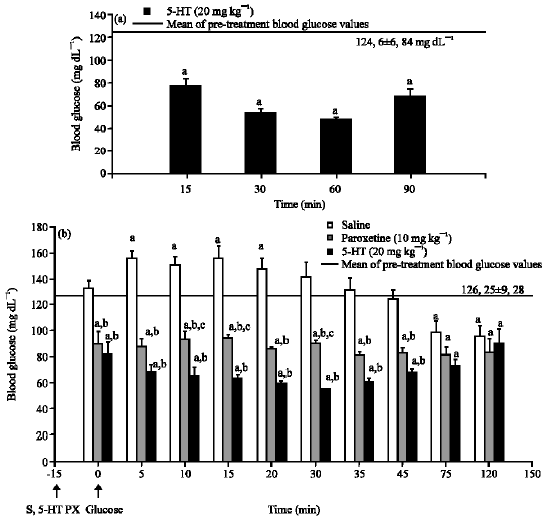 | |
| Fig. 5: | The effect of serotonin (5-HT) on blood glucose level in normoglycemic mice and the effect of 5-HT and paroxetine (PX) in intraperitoneal glucose tolerance test (B). Results are expressed as Mean±SEM. a: p<0.05, compared with the mean of pre- treatment blood glucose values; b: p<0.05, compared with saline group; c: p<0.05, compared with 5-HT group. S: Saline, PX: Paroxetine, 5-HT: Serotonin |
The coupling factor for the secretion of insulin from beta cells is known to be intracellular calcium. An increase in intracellular calcium is provided both by calcium influx via. voltage-sensitive calcium channels and release from intracellular stores (Davis, 2006; Yang and Gills, 2004; Wasada, 2002; Straub and Sharp, 2002). The mediators of this release from intracellular stores are inositol triphosphate (IP3) and cAMP. The contribution of ATP which is the product of glucose metabolism has also been noted. Closure of ATP-dependent potassium channels causes a depolarization which triggers the opening of voltage-sensitive calcium channels.
While the ATP-dependent potassium channel opener, diazoxide, caused an increase in blood glucose level, possibly by preventing insulin secretion due to inhibition of membrane depolarization, paroxetine alone at the 10 mg kg-1 dose given after diazoxide reversed this increase and caused a decrease in blood glucose to a level lower than pre-treatment measurements. Each drug showed its own but opposite effect on blood glucose. In the combination group of paroxetine with diazoxide, diazoxide did not prevent the hypoglycemic effect of paroxetine. If the hypoglycemic effect of paroxetine is related to the potassium channels, this can be explained by the reclosure of ATP-dependent potassium channels by paroxetine which were previously opened by diazoxide. In the literature, there are no studies showing the direct effect of paroxetine on ATP-dependent potassium channels in the pancreas. However, there are two studies reporting the inhibitory effect of paroxetine on TREK channels (a type of potassium channel expressed throughout the central nervous system) and G-protein activated potassium channels (Kobayashi et al., 2006; Thummler et al., 2007). Hence if the effect of paroxetine does not depend on any interaction with ATP-dependent potassium channels, then this effect may be due to another mechanism activated by paroxetine.
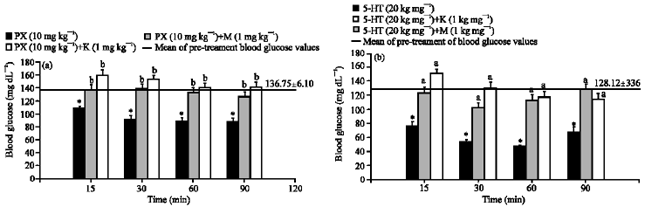 | |
| Fig. 6: | The effect of serotonin (5-HT) (A) and paroxetine (PX) (B) on blood glucose level in the presence of methysergide (M) and ketanserin (K). Results are expressed as Mean±SEM. *p<0.05, compared with the mean of pre-treatment blood glucose values, a: p<0.05, compared with 5-HT group; b: p<0.05, compared with paroxetine group |
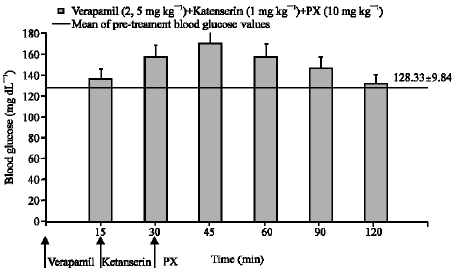 | |
| Fig. 7: | The effect of paroxetine (PX) on blood glucose level in the presence of verapamil plus ketanserin in normoglycemic mice. Results are expressed as Mean±SEM |
The opening of voltage-sensitive calcium channels situated in the membrane also caused an increase in intracellular calcium level. The calcium channel blocker, verapamil, has been previously reported to cause hyperglycemia following a decrease in insulin secretion in vitro (Semple et al., 1983). Paroxetine was shown to be a calcium channel blocking agent in some smooth muscles (Yaris et al., 2003). The hypoglycemic effect of paroxetine in our study is thought to be attributed to its calcium channel blocking effect. However, verapamil, a specific blocker of these channels did not show any effect on blood glucose level in our study and paroxetine decreased blood glucose level in the presence of verapamil. We can conclude that paroxetine shows its hypoglycemic effect via. different mechanisms rather than calcium channels. Another finding supporting this suggestion was the ineffectiveness of verapamil on the hypoglycemia induced by paroxetine administered after verapamil.
Paroxetine, as an inhibitor of 5-HT re-uptake may be expected to strengthen serotonergic action in the central nervous system (Bourin et al., 2001). The results of other in vitro studies suggest that this action may be valid in some peripheral systems. Thus, the strengthening of serotonergic action may be observed in the periphery and possibly in the pancreas. The similar effects of 5-HT and paroxetine seen in our study, such as decreasing blood glucose levels, support this idea. In addition, the inhibitory actions of the 5-HT antagonists were similar both for 5-HT and paroxetine. It was previously shown that 5-HT is present in the nerves innervating the pancreas in many mammalian species (Adeghate et al., 1999). The identification of serotoninergic nerves in the periaciner regions of the pancreas indicates that serotonin may play a role in the regulation of acinar secretion and stimulates the endocrine secretagogue cells. It was also shown that serotonin elicited a concentration-dependent increase in insulin secretion and thereafter, a decrease in blood glucose levels (Adeghate et al., 1999; Sugimoto et al., 2003; Yamada et al., 1999; Sugimoto et al., 1990). In our study, correlated with literature, the decreasing effect of serotonin on blood glucose level was also thought to be related to the increase in insulin secretion stimulated by serotonin. 5-HT1A and 5-HT2A receptors are thought to be involved in glucose regulation (Sugimoto et al., 2005). The decreasing effect of 5-HT on blood glucose levels in our study is possibly related to this secretory action. 5-HT2A receptors which are thought to involved in the secretory action of 5-HT, are coupled to the enzyme phospholipase C situated in the membrane. The activation of this enzyme is known to cause the production of IP3 and release of calcium from intracellular stores. Although it was not proved that paroxetine caused a decrease in blood glucose levels which was directly related to an increase in insulin secretion, it can be speculated that the effect of paroxetine on blood glucose level might have been secondary to an increase in insulin secretion via serotonergic mechanisms and/or receptors. Further studies such as determining insulin level, or incubating pancreatic beta cells with paroxetine will help to clarify the decreasing effect of paroxetine on blood glucose level.
In conclusion, paroxetine causes a decrease in blood glucose level probably by an increase in insulin secretion and this increase is provided possibly by intracellular mechanisms mediated by serotonergic receptors and postreceptor events, rather than an influx of calcium from the extracellular medium.
ACKNOWLEDGMENT
This study was supported by Karadeniz Technical University Research Fund (Grant No. 2004.114.003.11). The authors wish to thank Founder of Karadeniz Technical University for their special contribution. Part of this study is presented as poster presentation in 4th Panhellenic Congress of Pharmacology, 19-20 May 2006, Greece and published in the issue of Review of Clinical Pharmacology and Pharmacokinetics devoted to this congress and also presented an oral presentation and poster presentation in 19th National Congress of Pharmacology, Turkish Pharmacological Society. 24-27 September 2007, Turkey. The manuscript is edited professionally by International Science Editing service for English language.
REFERENCES
- Adeghate, E., A.S. Ponery, D. Pallot, S.H. Parvez and J. Singh, 1999. Distribution of serotonin and its effect on insulin and glucagon secretion in normal and diabetic pancreatic tissues in rat. Neuro Endocrinol. Lett., 20: 315-322.
PubMed - Behnam-Rassouli, M., M.B. Ghayour and N. Ghayour, 2010. Microvascular complications of diabetes. J. Biol. Sci., 10: 411-423.
CrossRefDirect Link - Chaudhari, U.P., A. Raje, N.D. Trivedi and A.N. Bhandari, 2010. Antidepressant like effect of N(G)-nitro-l-arginine methyl ester. Int. J. Pharmacol., 6: 183-191.
CrossRefDirect Link - Gletsu, N., T.N. Doan, J. Cole, R.L. Sutliff and K.E. Bernstein, 2005. Angiotensin II-induced hypertension in mice caused an incerase in insulin secretion. Vascul. Pharmacol., 42: 83-92.
PubMed - Gomez, R., J. Huber, F. Lhullier and H.M.T. Barros, 2001. Plasma insulin levels are increased by sertraline in rats under oral glucose overload. Braz. J. Med. Biol. Res., 34: 1569-1572.
Direct Link - Gomez, R., J. Huber, G. Tombini and H.M. Barros, 2001. Acute effect of different antidepressants on glycemia in diabetic and non-diabetic rats. Braz. J. Med. Biol. Res., 34: 57-64.
PubMed - Kennedy, S.H., K.R. Evans, S. Kruger, H.S. Mayberg and J.H. Meyer et al., 2001. Changes in regional brain glucose metabolism measured with positron emission tomography after paroxetine treatment of major depression. Am. J. Psychiatry, 158: 899-905.
Direct Link - Kesim, M., E.N. Duman, M. Kadioglu, C. Ulku, E. Muci, N.I. Kalyoncu and E. Yaris, 2006. Antinociceptive effects of fluoxetine and paroxetine with their related actions on glycemia in mice. Neuro Endocrinol. Lett., 27: 281-287.
PubMed - Kobayashi, T., K. Washiyama and K. Ikeda, 2006. Inhibition of G protein-activated inwardly rectifying K+ channels by the antidepressant paroxetine. J. Pharmacol. Sci., 102: 278-287.
CrossRef - Mahmood, D., M. Akhtar, D. Vohora and R. Khanam, 2010. Comparison of antinociceptive and antidiabetic effects of sertraline and amitriptyline on streptozotocin-induced diabetic rats. Hum. Exp. Toxicol., 29: 881-886.
PubMed - Moghtaderi, A., R. Alavi-Naini, M. Mohammadi and H. Rashidi, 2006. Diabetic neuropathy: Determining the sensitivity of peripheral nerves during a short course of glycemic control. J. Medical Sci., 6: 818-822.
CrossRefDirect Link - Mohajeri, D., G. Mousavi and Y. Doustar, 2009. Antihyperglycemic and pancreas-protective effects of Crocus sativus L. (Saffron) stigma ethanolic extract on rats with alloxan-induced diabetes. J. Biol. Sci., 9: 302-310.
CrossRefDirect Link - Mohamed, A.M., F.Z. El-Sharkawy, S.A.A. Ahmed, W.M. Aziz and O.A. Badary, 2009. Glycemic control and therapeutic effect of Nigella sativa and Curcuma longa on rats with streptozotocin-induced diabetic hepatopathy. J. Pharmacol. Toxicol., 4: 45-57.
CrossRefDirect Link - Oswald, P., D. Souery and J. Mendlewicz, 2003. Fluvoxamine-induced hyperglycaemia in a diabetic patient with comorbid depression. Int. J. Neuropsychopharmacol., 6: 85-87.
PubMed - Pollak, P.T., S.D. Mukherjee and A.D. Fraser, 2001. Sertraline-induced hypoglycemia. Ann. Pharmacother., 35: 1371-1374.
CrossRefDirect Link - Sansone, R.A. and L.A. Sansone, 2003. Sertraline-induced hyperglycemia: Case report. Int. J. Psychiatry Med., 33: 103-105.
PubMed - Semple, C.G., J.A. Thomson, G.H. Beastall and A.R. Lorimer, 1983. Oral verapamil does not affect glucose tolerance in non-diabetics. Br. J. Clin. Pharmacol., 15: 570-571.
PubMed - Straub, S.G. and G.W.G. Sharp, 2002. Glucose-stimulated signaling pathways in biphasic insulin secretion. Diabetes/Metab. Res. Rev., 18: 451-463.
CrossRefDirect Link - Sugimoto, Y., K. Inoue and J. Yamada, 2003. The tricyclic antidepressant clomipramine increases plasma glucose levels of mice. J. Pharmacol. Sci., 93: 74-79.
CrossRefDirect Link - Sugimoto, Y., N. Takashima, T. Noma and J. Yamada, 2005. Inhibitory effects of the 5-HT(1A) receptor agonist buspirone on stress-induced hyperglycemia in mice: Involvement of insulin and a buspirone metabolite, 1-(2- pyrimidinyl)piperazine (1-PP). Biol. Pharm. Bull., 28: 733-735.
PubMed - Taliyan, R. and P.L. Sharma, 2010. Diabetic neuropathic pain: An update and novel pharmacological strategies for relief of pain. J. Med. Sci., 10: 93-109.
CrossRefDirect Link - Thummler, S., F. Duprat and M. Lazdunski, 2007. Antipsychotics inhibit TREK but not TRAAK channels. Biochem. Biophys. Res. Commun., 354: 284-289.
CrossRef - Velavan, S. and V.M.H. Begum, 2007. Modulatory role of Asparagus racemosus on glucose homeostasis in aged rats. Int. J. Pharmacol., 3: 149-154.
CrossRefDirect Link - Wasada, T., 2002. Adenosine triphosphate-sensitive potassium (K(ATP)) channel activity is coupled with insulin resistance in obesity and type 2 diabetes mellitus. Intern. Med., 41: 84-90.
Direct Link - Weber-Hamann, B., M. Gilles, F. Lederbogen, I. Heuser and M. Deuschle, 2006. Improved insulin sensitivity in 80 nondiabetic patients with MDD after clinical remission in a double-blind, randomized trial of amitriptyline and paroxetine. J. Clin. Psychiatry, 67: 1856-1861.
PubMed - Yang, Y. and K.D. Gills, 2004. A highly Ca2+-sensitive pool of granules is regulated by glucose and protein kinases in insulin-secreting INS-1 cells. J. Gen. Physiol., 124: 641-654.
CrossRefDirect Link - Yaris, E., M. Kesim, M. Kadioglu, N.I. Kalyoncu, C. Ulku and R. Ozyavuz, 2003. The effects of paroxetine on rat isolated vas deferens. Pharmacol. Res., 48: 335-345.
PubMed