
P. Tep-Areenan
Department of Physiology, Faculty of Medicine, Srinakharinwirot University, Bangkok 10110, Thailand
P. Sawasdee
Natural Products Research Unit, Department of Chemistry, Faculty of Science, Chulalongkorn University, Bangkok 10330, Thailand
International Journal of Pharmacology
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 119-124







ABSTRACT
The aim of the present research was to study vasorelaxant effects of dichloromethane extract of Anaxagorea luzonensis (CH2Cl2-AL) and its underlying mechanisms. CH2Cl2-AL (1-300 μg mL-1) induced concentration-dependent vasorelaxations which were reduced by endothelial denudation, 300 μM NG-nitro-L-arginine methyl ester (L-NAME) and a combination of 10 μM indomethacin and 300 μM L-NAME, but not indomethacin alone. Raising the extracellular KCl concentration to 60 mM inhibited vasorelaxant responses to CH2Cl2-AL in both endothelium-intact and -denuded rings. Moreover, the responses to CH2Cl2-AL were inhibited by 30 μM barium chloride, 2 μM clotrimazole, 10 μM glibenclamide and 10 μM 1-[(2-chlorophenyl) diphenylmethyl]-1H-pyrazole (TRAM-34), but not 1 mM 4-aminopyridine. Pre-incubation with CH2Cl2-AL (1-100 μg mL-1) inhibited contractions induced by CaCl2 in a Ca2+-free, high KCl buffer. The present findings demonstrate, in the rat isolated aorta, that vasorelaxant responses to CH2Cl2-AL are, in part, mediated via the endothelium and NO-dependent pathways. Moreover, activation of KIR, KCa, KATP channels seems to play a role in CH2Cl2-AL-induced responses. Interestingly, Inhibition of extracellular Ca2+ influx is largely involved in the action of CH2Cl2-AL. The present study provides scientific evidence to support the use of CH2Cl2-AL as a vasodilator agent.
PDF Abstract XML References Citation
Received: November 02, 2010;
Accepted: January 06, 2011;
Published: February 09, 2011
How to cite this article
P. Tep-Areenan and P. Sawasdee, 2011. The Vasorelaxant Effects of Anaxagorea luzonensis A. Grey in the Rat Aorta. International Journal of Pharmacology, 7: 119-124.
DOI: 10.3923/ijp.2011.119.124
URL: https://scialert.net/abstract/?doi=ijp.2011.119.124
DOI: 10.3923/ijp.2011.119.124
URL: https://scialert.net/abstract/?doi=ijp.2011.119.124
INTRODUCTION
Anaxagorea luzonensis A. Grey (AL) belongs to Annonaceae family, a pantropical angiosperm family (Scharaschkin and Doyle, 2005, 2006). Anaxagorea is the only genus of Annonaceae with distribution in the Southeast Asia and America (Scharaschkin and Doyle, 2005). In Thailand, AL has been known as Kam-lang-wua-thalueng. It is a shrub with light green, smooth, oblong leaves, 7-15 cm long and 3-5 cm wide. The flowers are solitary opposed to the leaves of about 2 cm in diameter with a short peduncle. Sepals are ovate and obtuse. Petals are ovate and oblong. Stamens are all similar and fertile. There are 2-4 oblong ovaries. Carpels are about 4 cm spatulate, obtuse, obliquely mucronate, glabrous and rather rugose. Heartwood is reddish-brown, hard, with a little of the whitish bark (Hooker, 1875; Na Songkhla, 1982).
The heartwood of AL has been widely used in Thai traditional medicine as a health promoting herb. The extract of AL has several pharmacological effects including blood tonic, antihistamine, antioxidant and antihypertensive agents (Mokkhasmit et al., 1971; Gonda et al., 2000; Takada and Yokoyama, 2000; Panriansaen et al., 2008).
Chemical investigations have demonstrated that the extract of the heartwoods of AL contains flavones, flavonones, flavonols and xanthones (Kitaoka et al., 1998; Gonda et al., 2000; Takada and Yokoyama, 2000). For example, 20 natural compounds are isolated from the bark of AL including one new flavonoid, 3, 5, 7, 4'-tetrahydroxy-2'-methoxyflavone and five new xanthones, 1, 3, 6-trihydroxy-5-methoxy-4-prenylxanthone, 1, 3, 5-trihydroxy-6-methoxy-2-prenylxanthone, 1, 3, 5-trihydroxy-4-(3-hydroxy-3-methylbutyl) xanthone, 1, 3, 6-trihydroxy-4-prenylxanthone, 3, 6-dihydroxy-1, 5-dimethoxyxanthone together with seven known xanthones, 1, 3, 5-trihydroxy-4-prenylxanthone, 1, 3, 5-trihydroxy-2-prenylxanthone, 1, 3, 6-trihydroxy-5-methoxyxanthone, genistein (1, 3, 7-trihydroxyxanthone), 1, 3, 5-trihydroxyxanthone, 3, 5-dihydroxy-1-methoxyxanthone and 1, 3, 5, 6-tetrahydroxy xanthone and seven known flavonoids, biochanin A, chrysin, 3'-methylorobol, orobol, taxifolin, kaempferol and quercetin. Almost all flavonoids and 1, 3, 5, 6-tetrahydroxy xanthone exhibit antioxidant activity toward α,α-diphenyl-β-picryhydrazyl radical (Gonda et al., 2000). In addition, 3, 5, 7, 4'-tetrahydroxy-2'-methoxyflavone isolated from AL has antioxidant effect on autoxidation of epinephrine hydrochloride (Takada and Yokoyama, 2000). A previous study has reported that 8-isopentenylnaringenin in the extract exerts estrogenic activity greater than genistein, a standard compound (Kitaoka et al., 1998).
In other plants, methoxyflavones and flavonoids induce vasorelaxation in isolated blood vessels (Ajay et al., 2003; Morello et al., 2006; Tep-Areenan et al., 2010; Tep-Areenan and Sawasdee, 2010). However, there is no pharmacological evidence about the vasorelaxant effects of AL which is rich in flavones and flavonoids. The present study aimed to investigate the vasorelaxant effect of the dichloromethane extract of AL on vascular tone and its underlying mechanisms in the isolated rat aorta.
MATERIALS AND METHODS
Extraction of Anaxagorea luzonensis A. Grey (AL): The dried and chopped heartwood (5 kg) of AL was extracted four times with methanol using soxhlet extractor. After filtration, the methanolic extract was concentrated under reduced pressure and was then partitioned with dichloromethane (CH2Cl2), yielding about 10.5 g of CH2Cl2-AL after evaporation.
Tissue preparation: In 2009, Experiments were performed using aorta obtained from male Wistar rats (300-350 g) purchased from National Laboratory Animal Center, Mahidol University, Thailand. Rats were housed in standard environmental condition (25°C) under 12 h light/dark cycles and fed with standard laboratory rat chow and tap water ad libitum. All experimental procedures were reviewed and approved by the Animal Research Ethics Committee of the Faculty of Medicine, Srinakharinwirot University.
Male Wistar rats were anaesthetized with Zolitil 50 mg kg-1 (tiletamine chloridrate and zolazepan chloridrate) into quadriceps muscle and killed by cervical dislocation. Following a thoracotomy, the thoracic aorta was carefully removed, cleaned of fat and connective tissue and cut into 5 mm ring segments. Each ring was transferred to a jacketed organ bath filled with 20 mL of modified Krebs-Henseleit solution, composed of (mM) NaCl 118, KCl 4.7, MgSO4 1.2, KH2PO4 1.2, NaHCO3 25, CaCl2 2, D-glucose 10, that was maintained at 37°C and bubbled continuously with 95 % O2 and 5% CO2 mixture. The solution in the organ bath was exchanged every 15 min for 1 h. The rings were mounted between two triangular stainless steel hooks that were passed through the lumen and stretched to an optimal passive tension of about 1 g and maintained at this tension for 1 h.The upper hook was connected to a isometric force transducer (MLT 0210, New South Wales, Australia) and changes in isometric force were recorded on a MacLab recording system (AD instruments, New South Wales, Australia).
Experimental protocol: Following a 1 h equilibration period, methoxamine (10-60 μM) was used to increase tone by approximately 1 g. Once a stable contraction was estabilshed, CH2Cl2-AL (1-300 μg mL-1) was added cumulatively. To explore the mechanisms involved in vasorelaxation induced by CH2Cl2-AL, aortic rings were incubated with various inhibitors added to the organ bath before methoxamine was added to increase tone. In vehicle-control experiments, dimethyl sulphoxide (DMSO) alone was added cumulatively in the same volumes as those used in the experiments with CH2Cl2-AL.
To examine the contribution of the endothelium in vasorelaxant responses to CH2Cl2-AL, the endothelium was mechanically removed by gently rubbing the luminal surface with a cocktail stick. Removal of the endothelium was demonstrated by vasorelaxation to 10 μM carbachol being less than 10% of the induced tone. To investigate the involvement of vasodilator prostanoids via the cyclooxygenase (COX) pathway and Nitric Oxide (NO) in vasorelaxation to CH2Cl2-AL, aortic rings were treated with indomethacin (10 μM), a COX inhibitor and NG-nitro-L-arginine methyl ester (L-NAME, 300 μM), an inhibitor of endothelial nitric oxide synthase, respectively.
To investigate the potential involvement of K+ channels in vasorelaxation to CH2Cl2-AL, aortic rings were pre-contracted with a high extracellular concentration of KCl (60 mM), which was prepared by replacing an equimolar concentration of NaCl with KCl (Tep-Areenan et al., 2003). These experiments were also performed in endothelium-denuded rings to determine any involvement of Endothelium-Derived Relaxing Factors (EDRFs) in the vasorelaxant effects of CH2Cl2-AL on activation of K+ channels. To characterized the types of K+ channels involved in vasorelaxation to CH2Cl2-AL, concentration-response curves to CH2Cl2-AL were constructed after incubation with 4-aminopyridine (4-AP, 1 mM), a voltage-gated K+ (KV) channel inhibitor, glibenclamide (10 μM), an ATP-sensitive (KATP) inhibitor, or barium chloride (BaCl2, 30 μM), an inward-rectifier (KIR) channel inhibitor, clotrimazole (2 μM), an intermediate-conductance calcium-activated potassium (IKCa) channel inhibitor and 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole (TRAM-34, 10 μM), a specific inhibitor of IKCa channels.
To examine the vascular effect of CH2Cl2-AL on extracellular Ca2+ influx, concentration-response curve to CaCl2 (10 μM-30 mM) were constructed in the presence and absence of CH2Cl2-AL (1, 10 and 100 μg mL-1) for 30 min. Aortic rings were first allowed to equilibrate at 1 g tension in a Ca2+-free Krebs solution and then the rings were bathed with Ca2+-free, high KCl (100 mM) Krebs solution. In vehicle-control experiments, DMSO was added in the same volume as that used in the experiments with CH2Cl2-AL.
Data and statistical analysis: The concentration of vasorelaxant giving half-maximal relaxation (EC50) and maximal responses (Rmax) were obtained from the concentration-response curve fitted to a sigmoidal logistic equation using the GraphPad Prism package described by Tep-Areenan et al. (2003). Rmax and pEC50 values (negative logarithm of the EC50) were compared by analysis of variance (ANOVA) with statistically significant differences between groups being determined by Bonferroni’s post-hoc test. These were expressed as Mean±SEM. The results were considered statistically significant when p value was less than 0.05. The number of animals in each group is represented by n.
Chemicals: All drugs and chemicals were purchased from Sigma Chemical Company (St. Louis, Missouri, USA), but zoletil was purchased from Virbac (Carros Cedex, France). Indomethacin was dissolved in ethanol. CH2Cl2-AL and glibenclamide were dissolved in DMSO. 4-AP, BaCl2 and TRAM-34 were dissolved in distilled water. The remaining drugs were dissolved in the Krebs solution. All drugs were made up on the day of the experiment.
RESULTS
The effects of endothelial denudation, indomethacin and L-NAME on vasorelaxation to CH2Cl2-AL in rat aortic rings: CH2Cl2-AL (1-300 μg mL-1) caused vasorelaxation in a concentration-dependent manner (pEC50 = 4.87±0.08 with Rmax = 118±4%, n = 6, Fig. 1). Removal of the endothelium significantly (p<0.05) reduced the effects of CH2Cl2-AL (Rmax: control = 118±4%, n = 6; denuded = 96.1±7.5%, n = 6, Fig. 1). Similarly, vasorelaxant responses to CH2Cl2-AL were significantly (p<0.05) reduced after pretreatment with a combination of indomethacin plus L-NAME, which were not different from L-NAME alone (Rmax: L-NAME alone = 99.3±5.5%, n = 6; indomethacin plus L-NAME = 95.3±8.6%, n = 6, Fig. 1).
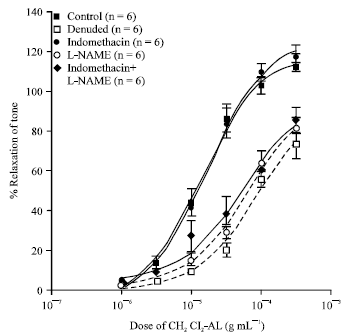 | |
| Fig. 1: | Effects of removal of the endothelium (denuded), indomethacin (10 μM), NG-nitro-L-arginine methyl ester (L-NAME, 300 μM) and a combination of indomethabin and L-NAME on vasorelaxation to CH2Cl2-AL in aortic rings precontracted with methoxamine. Data were shown as Mean±SEM |
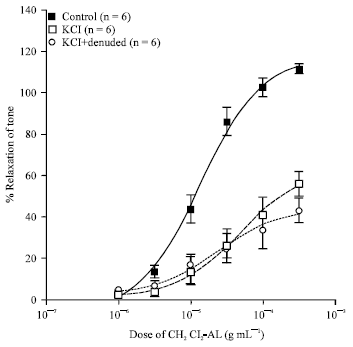 | |
| Fig. 2: | Effects of a high concentration of extracellular K+ (60 mM KCl) on vasorelaxation to CH2Cl2-AL in endothelium-intact and -denuded aortic rings precontracted with methoxamine. Data were shown as Mean±SEM |
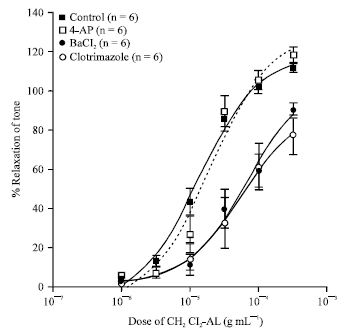 | |
| Fig. 3: | Effects of 4-aminopyridine (4-AP, 1 mM), barium chloride (BaCl2, 10 μM) and clotrimazole (2 μM) on vasorelaxation to CH2Cl2-AL in aortic rings precontracted with methoxamine. Data were shown as Mean±SEM |
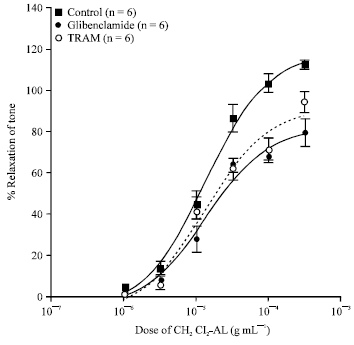 | |
| Fig. 4: | Effects of glibenclamide (10 μM) and 1-[(2-chlorophenyl)diphenylmethyl]-1H-pyrazole (TRAM-34, 10 μM) on vasorelaxation to CH2Cl2-AL in aortic rings precontracted with methoxamine. Data were shown as Mean±SEM |
The effects of high extracellular potassium and potassium channel inhibitors on vasorelaxation to CH2Cl2-AL: Raising extracellular K+ concentration (60 mM KCl) significantly (p<0.001) inhibited vasorelaxation induced by CH2Cl2-AL in endothelium-intact and-denuded rings (Rmax: 60 mM KCl = 63.1±9.1%, n = 6; 60 mM KCl with denuded = 44.4±6.1%, n = 6, Fig. 2). However, there was no significant difference between two groups.
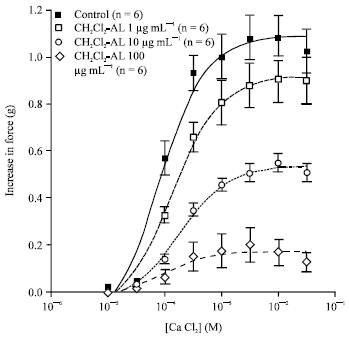 | |
| Fig. 5: | Effects of CH2Cl2-AL on CaCl2-induced contraction in aortic rings depolarized by 100 mM KCl. Data were shown as Mean±SEM |
In intact endothelium, vasorelaxant effects of CH2Cl2-AL were significantly (p<0.05) inhibited by either BaCl2, or clotrimazole, but not 4-AP (Rmax: BaCl2 = 101±5%, n = 6; clotrimazole = 91.7±10.8%, n = 6, Fig. 3). Moreover, pretreatment with glibenclamide or TRAM-34 significantly (p<0.001) inhibited vasorelaxation induced by CH2Cl2-AL (Rmax: glibenclamide = 82.5±4.35%, n = 6; TRAM-34 = 91.0±4.7%, n = 6, Fig. 4).
The effects of CH2Cl2-AL on CaCl2-induced contraction in rat aortic rings: CaCl2 (10 μM-30 mM) elicited concentration-dependent contraction of KCl (100 mM) depolarized rings in Ca2+-free medium. CH2Cl2-AL concentration-dependently reduced contractions induced by CaCl2 (Rmax: control = 1.09±0.04 g, n = 6; 1 μg mL-1 CH2Cl2-AL = 0.92±0.04 g, n = 6; 10 μg mL-1 CH2Cl2-AL = 0.54±0.01 g, n = 6; 100 μg mL-1 CH2Cl2-AL = 0.17±0.02 g, n = 6, Fig. 5).
DISCUSSION
The present study has demonstrated, for the first time, about the vasorelaxant effects of CH2Cl2-AL and mechanisms involved in its action. CH2Cl2-AL causes vasorelaxation in the isolated rat aorta, which is partly due to endothelium-derived No. Activation of K+ channels and inhibition of Ca2+ influx are largely involved in the vasorelaxant effects of CH2Cl2-AL.
Previous studies have reported that the extract of Anaxagorea luzonensis A.Gray contains several flavones and flavonoids (Kitaoka et al., 1998; Gonda et al., 2000). Therefore, vasorelaxant responses of rat aortic rings to CH2Cl2-AL may be caused by the effects of these compounds. The present findings are in agreement with several earlier investigations with plant extract showing that flavonoids exert a vasorelaxant property (Herrera et al., 1996; Ajay et al., 2003; Gilani et al., 2006; Morello et al., 2006). Recently, studies by our group have shown that the ethanolic extract of Kaempferia parviflora and its methoxyflavones cause vasorelaxation in the rat isolated aorta (Tep-Areenan et al., 2010; Tep-Areenan and Sawasdee, 2010).
The vascular endothelium plays an essential role in regulating vascular tone via synthesis and release Endothelium-Derived Relaxing Factors (EDRFs), such as NO and prostacyclin (Lokhande et al., 2006; Vanhoutte, 2009; Vanhoutte et al., 2009). The present experiments show that relaxations induced by CH2Cl2-AL were inhibited by removal of the endothelium. Moreover, the effects of CH2Cl2-AL were inhibited by L-NAME, a NOS inhibitor, but not inhibition of a COX pathway by indomethacin. These results indicate the participation of endothelium-derived NO in the effects of CH2Cl2-AL. The present study also found that pretreatment with L-NAME plus indomethacin showed no further inhibition of responses to CH2Cl2-AL than those observed with L-NAME alone. Therefore, these results indicate that the relaxant responses to CH2Cl2-AL involve NO, but not vasodilator prostanoids via the COX pathway. Although, previous studies have demonstrated that vasorelaxant effects of some flavones and flavonoids involve the COX pathway (Ajay et al., 2003; Uydes-Dogan et al., 2005), our CH2Cl2-AL constituents may be different. Further studies would be needed to identify the active ingredients in our extract.
Several types of K+ channels are located in vascular smooth muscle cells, including KATP, KCa, KV, and KIR channels. Opening of K+ channels in the cell membrane of vascular smooth muscle cells increases K+ efflux, causing hyperpolarization, which closure of voltage-gated Ca2+ channels and subsequently vasorelaxation (Jackson, 2000; Sobey, 2001). To investigate the involvement of K+ channels in vasorelaxant effects of CH2Cl2-AL, a high concentration of KCl was used to increase vascular tone. It was found that 60 mM KCl inhibited the effects of CH2Cl2-AL in both endothelium-intact and denuded aortic rings. However, there was no further inhibition of responses to CH2Cl2-AL in denuded rings, compared to intact rings. Taken together, it is suggested that opening of K+ channels on smooth muscle cells by CH2Cl2-AL is unlikely due to EDRFs.
In order to investigate the contribution of exact types of K+ channels involved in vasorelaxant effects of CH2Cl2-AL, we used different types of K+ channel inhibitors. It was found that vasorelaxation induced by CH2Cl2-AL were inhibited by barium chloride, a KIR channel inhibitor, glibenclamide, a KATP channel inhibitor, clotrimazole, an intermediate-conductance KCa channels, TRAM-34, a specific inhibitor of intermediate-conductance KCa (IKCa) channels. Conversely, 4-aminopyridine, a KV channel inhibitor, did not affect vascular responses to CH2Cl2-AL. These results suggest that vasorelaxant responses to CH2Cl2-AL are mediated by increasing K+ efflux, at least in part, through KIR, KATP, IKCa channels.
Another aspect examined in the present study was whether vasorelaxant responses to inhibition of extracellular Ca2+ influx involved. We found that contractile responses of rat aortic rings to CaCl2 in Ca2+ -free medium containing KCl were inhibited by CH2Cl2-AL. These findings support the notion that CH2Cl2-AL can block Ca2+ influx from the extracellular space. In addition, in the presence of CH2Cl2-AL, there were rightward shifts in concentration-response curves for CaCl2 with reduction in the Rmax values, thus suggesting that CH2Cl2-AL acts as a non-competitive calcium antagonist. Clinically, Ca2+ antagonist are potentially used to treat hypertension (Ishimitsu et al., 2009; Stokes, 2009; Adatia and Shekerdemian, 2010). Concerning the vasorelaxant effects of CH2Cl2-AL, it is likely to suggest that the extract of Anaxagorea luzonensis A. Grey may acts as antihypertensive agent.
CONCLUSION
From our data, it is possible to conclude that CH2Cl2-AL exerts its vasorelaxant effects by acting on multiple sites of actions. Vascular responses to CH2Cl2-AL are partly mediated by endothelium-dependent No. Additionally, inhibition of Ca2+ entry and activation of K+ channels are required for the vasorelaxant effects of CH2Cl2-AL. Further chemical and pharmacological experiments are required to isolate and identify the active constituents responsible for this vascular effect and to investigate the in vivo effect of GL extract.
ACKNOWLEDGMENTS
This study was funded by Srinakharinwirot University (Grant No. 375/2552). Financial support for P. Sawasdee through the National Research University Project of CHE and Ratchadaphiseksomphot Endowment Fund (FW001A) is very gratefully acknowledged. We would like to express our deepest gratitude to Dr. Alfredo Villarroel for improving the English. We also thank Mr. Phongphat Wetchasit for his technical support. The authors have no conflict of interest to report.
REFERENCES
- Adatia, I. and L. Shekerdemian, 2010. The role of calcium channel blockers, steroids, anticoagulation, antiplatelet drugs and endothelin receptor antagonists. Pediatr. Crit. Care Med., 11: S46-S52.
PubMedDirect Link - Ajay, M., A.H. Gilani and M.R. Mustafa, 2003. Effects of flavonoids on vascular smooth muscle of the isolated rat thoracic aorta. Life Sci., 74: 603-612.
CrossRefPubMedDirect Link - Gilani, A.H., M.N. Ghayur, P.J. Houghton, Q. Jabeen, S.F. Kazim, M.I. Jumani and S.A. Saeed, 2006. Studies on the hypotensive, cardio-suppressant, vasodilator and antiplatelet activities of betel nut crude extract and its constituents. Int. J. Pharmacol., 2: 33-41.
CrossRefDirect Link - Gonda, R., T. Takeda and T. Akiyama, 2000. Studies on the constituents of Anaxagorea luzonensis A. GRAY. Chem. Pharm. Bull. (Tokyo), 48: 1219-1222.
Direct Link - Herrera, M.D., A. Zarzuelo, J. Jimenez, E. Marhuenda and J. Duarte, 1996. Effects of flavonoids on rat aortic smooth muscle contractility: Structure-activity relationships. Gen. Pharmacol., 27: 273-277.
CrossRef - Ishimitsu, T., A. Numabe, T. Masuda, T. Akabane, A. Okamura, J. Minami and H. Matsuoka, 2009. Angiotensin-II receptor antagonist combined with calcium channel blocker or diuretic for essential hypertension. Hypertens. Res., 32: 962-968.
CrossRefDirect Link - Kitaoka, M., H. Kadokawa, M. Sugano, K. Ichikawa and M. Taki et al., 1998. Prenylflavonoids: A new class of non-steroidal phytoestrogen (Part 1). Isolation of 8-isopentenylnaringenin and an initial study on its structure-activity relationship. Planta Medica, 64: 511-515.
PubMed - Lokhande, P.D., B.S. Kuchekar, A.R. Chabukswar and S.C. Jagdale, 2006. Nitric oxide: Role in biological system. Asian J. Biochem., 1: 1-17.
CrossRefDirect Link - Morello, S., V. Vellecco, A. Alfieri, N. Mascolo and C. Cicala, 2006. Vasorelaxant effect of the flavonoid galangin on isolated rat thoracic aorta. Life. Sci., 78: 825-830.
CrossRef - Scharaschkin, T. and J.A. Doyle, 2005. Phylogeny and historical biogeography of Anaxagorea (Annonaceae) using morphology and non-coding chloroplast sequence data. Syst. Bot., 30: 712-735.
Direct Link - Scharaschkin, T. and J.A. Doyle, 2006. Character evolution in Anaxagorea (Annonaceae). Am. J. Bot., 93: 36-54.
Direct Link - Sobey, C.G., 2001. Potassium channel function in vascular disease. Arterioscler. Thromb. Vasc. Biol., 21: 28-38.
Direct Link - Stokes, G.S., 2009. Management of hypertension in the elderly patient. Clin. Interv. Aging, 4: 379-389.
CrossRefDirect Link - Takada, T. and S. Yokoyama, 2000. Antioxidant effects of new flavonoid isolated from Anaxagorea luzonensis A. Grey on autoxidation of epinephrine hydrochloride. Mater. Techonol., 18: 238-241.
Direct Link - Tep-Areenan, P. and P. Sawasdee, 2010. Vasorelaxant effects of 5,7,4`-trimethoxyflavone from Kaepmferia parviflora in the rat aorta. Int. J. Pharmacol., 6: 381-386.
CrossRefDirect Link - Uydes-Dogan, B.S., S. Takir, O. Ozdemir, U. Kolak, G. Topcu and A. Ulubelen, 2005. The comparison of the relaxant effects of two methoxylated flavones in rat aortic rings. Vascul. Pharmacol., 43: 220-226.
CrossRefPubMedDirect Link - Vanhoutte, P.M., 2009. Endothelial dysfunction: The first step toward coronary arteriosclerosis. Circ. J., 73: 595-601.
PubMedDirect Link - Vanhoutte, P.M., H. Shimokawa, E.H. Tang and M. Feletou, 2009. Endothelial dysfunction and vascular disease. Acta Physiol., 196: 193-222.
CrossRef