
T. Cherian
Department of Health Sciences, Higher Colleges of Technology, Dubai, United Arab Emirates
S. Singh
Department of Physiology, Faculty of Medicine, United Arab Emirates University, Al Ain, United Arab Emirates
T. Thomas
Department of Medicine, Government Medical College, Trivandrum, Kerala, India
International Journal of Pharmacology
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 472-479







ABSTRACT
The aim of the present study is to evaluate the role autonomic receptors in initiating neurogenic contractions and the role of post-synaptic alpha adrenergic receptors in controlling the contractility of the ovine ureter. Isolated sheep ureter strips were studied to clarify the autonomic receptor functions by investigating the actions of selective agonists and antagonists. Noradrenaline shows a well pronounced excitatory effect on uretero-pelvic and vesicular regions which can be blocked by phenoxybenzamine, a non-selective alpha adrenergic receptor blocker. Selective blockade of alpha-1 receptors with prazosin did not inhibit the action noradrenaline but yohimbine, a selective alpha-2 receptor blocker, abolished the effect of noradrenaline in pelvic, proximal and vesicular regions of the ureter. Selective blockade of alpha-1 receptors with prazosin inhibit the action of noradrenaline all regions of the ureter except pelvic, proximal and vesicular regions of the ureter. Clonidine, a selective alpha-2 agonist, produced a significant contractile effect on the ureter in the pelvic, proximal and vesicular regions but not significant effect on the rest of the regions of the ureter. Phenylephrine, a selective alpha-1 receptor agonist show remarkable contractile effect on all parts of the ureter except the pelvic, proximal and vesicular regions. High intensity, high frequency, short duration stimuli applied on spontaneously contracting ureteral segments show enhancement of contraction which can be blocked by pretreating the preparation with phenoxybenzamine and tetrodotoxin (TTX). This study suggests that post-synaptic receptors responsible for the mediation of the stimulatory action of noradrenaline are of the alpha-2 subtype in the pelvic, proximal and vesicular regions of the ureter and alpha-1 in the rest of the regions.
PDF Abstract XML References Citation
Received: April 04, 2010;
Accepted: May 21, 2010;
Published: September 02, 2010
How to cite this article
T. Cherian, S. Singh and T. Thomas, 2010. A Qualitative Study of the Autonomic Receptors Modulating the Contractile Activity of Isolated Ovine Ureter. International Journal of Pharmacology, 6: 472-479.
DOI: 10.3923/ijp.2010.472.479
URL: https://scialert.net/abstract/?doi=ijp.2010.472.479
DOI: 10.3923/ijp.2010.472.479
URL: https://scialert.net/abstract/?doi=ijp.2010.472.479
INTRODUCTION
The role of sympathetic innervation in promoting ureteral contraction is a subject of much controversy as both myogenic and neurogenic theories are always debated to know the exact functioning of the ureter. However the sympathetic innnervation to the kidney and ureter appears to play a modulating role in ureteral contractions (Schulman, 1985). Histochemical studies have shown the presence of autonomic innervation in human ureter with the presence of cholinergic and adrenergic fibers throughout the length (Prieto et al., 1993; Hannappel and Golenhofen, 1974; Morita et al., 1994; Ohki et al., 2010). Ureteral peristalsis is believed to be triggered spontaneously by the pace maker cells located at the pelvic-calyceal junctions by external stimuli which may be electrical, mechanical, chemical or conduction from an adjacent excited cell. It has been demonstrated that both electrical and mechanical stimulation of the ureter produce contractions that are propagated both proximally and distally from the site of excitation (Thulesius et al., 1986). This leads to the myogenic theory of ureteral contraction which proposes the spontaneous activity of the ureter to be purely a myogenic process without nervous influence. Demonstrations that denervated ureter and excised ureteral sections retain their normal rhythmic activity, have added support to this theory (Angelo-Khattar et al., 1985; Morita, 1986). Furthermore, it has shown that in excised ureteral strips prostaglandins also responsible for the propagation and maintenance of spontaneous ureteral activity (Thulesius et al., 1987; Angelo-Khattar et al., 1989). Furthermore, adrenergic and cholinergic agonists and antagonists have been demonstrated to have an effect on the ureter (Weiss et al., 1978; Hernandez et al., 1992) which coupled to the demonstration of the presence of nerve terminals (Prieto et al., 1994) in the proximal ureteral region provide evidence to support the influence of the autonomic nervous system on ureteral function. The role of both alpha-l and beta adrenoceptors in modulating the phasic contractility and the basal tone of the pig intravesical ureter has been well documented (Hernandez et al., 1992).
It has also been shown that electrical stimulation of excised ureteral segments with high intensity, high frequency and short duration stimuli, which were subthreshold for the direct excitation of ureteral smooth muscles, potentiated the spontaneously driven contractions through the release of endogenous alpha adrenergic agonist (Weiss et al., 1978).
In the clinical management of renal colic the stone passage is hindered by the overactive ureter with uncoordinated contractions of the ureter resulting in severe colic pain. Clinical studies indicate that in renal colic, drugs which allow continued peristalsis along with preventing uncontrolled irregular contractions would be more advantages for the expulsion of ureteral stone. Clinical success in the use of alpha-1 adrenergic receptor blockers to enhance the passage of distal ureteral stone by relaxing the ureter supports the influence of circulating epinephrine on the ureteral motility at the receptor level and the availability of specific receptors on different regions of the renal system (Davenport et al., 2007; Nakada et al., 2007; Hollingsworth et al., 2009). Furthermore when alpha adrenergic receptors are blocked epinephrine activates beta receptors to relax the ureter to a greater extent (Wheeler et al., 1990; Weiss et al., 1978). The role of alpha adrenergic subtypes involved in the control of ureteral peristalsis has not been fully evaluated even though norepinephrine causes a remarkable increase in the frequency of contractions (Tindall, 1972; Mayo and Halbert, 1981). It has been suggested in canine in vivo ureteral studies that the ureteral urine transport is controlled by the activation of both alpha-1 and alpha-2 adrenoceptors through regulation of peristaltic frequency and bolus volume (Morita et al., 1987).
This study has facilitated the characterization of the autonomic receptor function of the isolated sheep ureter especially the role of both alpha-1 and alpha-2 adrenergic receptors by investigating the actions of selective agonists and antagonists and demonstration of endogenous neurohumoral influence on modulating the ureteral contractions. Sheep has been selected as an animal choice is due to the easy availability of the specimens from the local abattoir and its histological and functional similarity to human ureter.
MATERIALS AND METHODS
The specimens used in this study were obtained from freshly slaughtered male Australian Merino sheep weighing approximately 30 kg. The ureters were excised with the kidneys en bloc and transported from the local abbatoire to the laboratory in thermos flasks containing chilled Krebs-Henseleit solution. The ureters were dissected free of connective tissue and fat and 4 mm long rings were cut from the ureter proximal to uretero-pelvic region of the kidney. The preparations were suspended vertically in 10 mL organ baths filled with Krebs-Henseleit solution, maintained at 37°C and bubbled with 95% oxygen and 5% carbon dioxide. The ureteral rings were attached to the bottom of the organ bath and the upper end connected to a Bioscience UFI force transducer. A preload of 2 g was applied to the tissue by adjustment of micrometer screw and isometric tension continuously recorded on a Gilson 5/6 thermal chart recorder system. The spontaneous onset of contractions was observed within 20 min and the preparation was allowed to equilibrate for 1 h. before the addition of cumulative dose of drugs to the organ bath. The pattern of rhythmic contractions is shown in Fig. 1. The amplitude of the contractions at 60 min is taken as the baseline value and any change in amplitude following the addition of drugs is calculated as a percentage increase or decrease of the original value. Cumulative dose response curves to noradrenaline (10-8-10-4 M) were obtained before and after pretreatment with the following adrenergic blocking agents for 30 min phenoxybenzamine 5x10-6 M, prazosin 10-6 M and yohimbine 10-6M. The actions of the adrenergic agonists adrenaline (10-8 -10-4), before and after pretreatment with the beta blocker propranolol 10-6 M was studied. The effects of cumulative doses of the adrenergic agonists, clonidine (10-8-10-4) and phenylephrine (10-8-10-4 M) on the ureteral strips were also investigated, both before and after blockade with prazosin and yohimbine, respectively. The actions of isoprenaline (10-8_10-4 M) was also examined. Cumulative dose-response curves to acetylcholine (10-8-10-4 M) alone and in the presence of atropine (5x10-6 M) were also obtained.
In another set of experiments proximal ureteral ring preparations obtained with the above procedure were mounted on a platinum needle electrode in specially designed stimulating 2 mL micro-organ bath in Krebs-Henseleit solution aerated with 95% oxygen and 5% carbon dioxide. The preparations were stretched by the needle terminals of the electrode and a preload of 2 g was applied. One hour after the onset of spontaneous contractions, the preparation was stimulated with high intensity, high frequency short duration pulses (10 V, 20-30 Hz, 1 m sec) delivered at a frequency of 3 min-1 using a Grass S-4 stimulator.
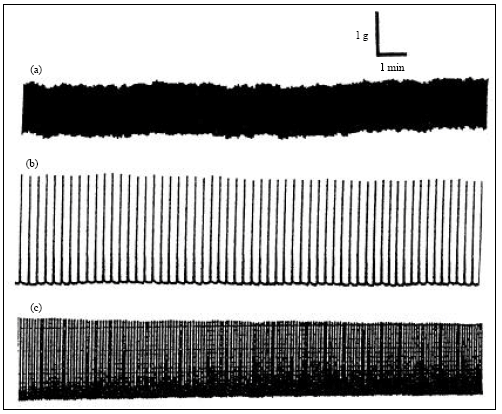 | |
| Fig. 1: | The panels a, b and c shows typical contractile pattern of normal sheep ureteral strips obtained from uretero-pelvic, middle and uretero-vesical regions, respectively |
The tissue was pre-treated with the following drugs for 30 min, phenoxybenzamine (10-6 M), propranolol (10-6 M) and repeated stimulation with high intensity, high frequency and short duration stimuli. The preparation was stimulated with the same parameters in presence of tetrodotoxin (TTX) (10-8 M).
Drugs and chemicals: The bathing medium Krebs-Henseleit solution contained (mM): NaCl 115.3, KCI 4.6, CaCl 2.3, MgSO4 1.2, NaHCO3 22.1, KH2PO4 1.1 and glucose 7.8 and was maintained at pH 7.4. The following drugs were used: adrenaline, noradrenaline, phenylephrine, yohimbine (Sigma Chemical Co., St.Louis, USA); prazosin (Pfizer, Brussels, Belgium); phenoxybenzamine (dibenyline, Smith Kline and French Lab Ltd., Hertz, UK); clonidine hydrochloride (Boehringer Ingelheim, West Germany); isoprenaline hydrochloride (Sigma Chemical Co., St.Louis, USA); acetylcholine chloride (BDH Chemicals Ltd., Pool, England); atropine sulphate, propranolol hydrochloride, tetrodotoxin (Sigma Chemical Co., St.Louis, USA).
Calculations: For each concentration of the adrenergic agonists and antagonists 10 experiments were performed and the amplitude is calculated as the percentage of maximum contractile response where 100% denotes the maximum response of the tissue before the addition of any drug. Competitive antagonism of the drugs are determined by using Schild plot (Arunlakshana and Schild, 1959). Standard statistical methods were used to calculate mean values, standard deviations and standard errors. The students independant t-test was used to determine statistical differences between two sets of data. The SPSS statistical program and GraphPad Prism version 5.03 were used for all statistical analyses and drawings.
RESULTS
Figure 1 shows typical contractile patterns of ring preparations obtained from the uretero-pelvic, middle and uretero-vesicular regions. The uretero-pelvic section is seen to have a higher frequency and lower amplitude of contractions than the rest of the ureter. The middle part shows distinct contractions with higher amplitudes and lower frequency and the uretero-vesical region shows higher frequency and lower amplitude compared to the middle region. The onset of contractions are slow initially for all regions but stabilizes after 1 h and trace shown in this Fig. 1 is 1 h after onset of contractions in the organ bath. Figure 2 shows the effect of noradrenaline, which resulted in a dose dependent rise in the amplitude with a minor increase in the frequency of ureteral contractions.
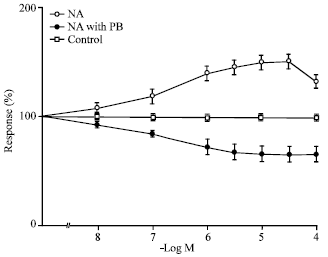 | |
| Fig. 2: | In vitro effect of nor-adrenaline on sheep uretero-pelvic region before and after pretreatment with phenoxybenzamine represented by open and closed circles respectively (n = 10). Open squares represent control values for each dose (n = 10) and 100 % denotes no change in control parameters. For treated and untreated preparations p<0.001 for 10-6 to 10-4 M, p<0.01 for 10-7 M and p is not significant for 10-8 M compared to controls. Mean value±SE are given; n: No. of experiments |
Regional differences were seen with respect to the actions of noradrenaline. The uretero pelvic and uretero-vesicular regions were found to be the most responsive to noradrenaline whereas the middle section of the ureter was not equally responsive to the agonist compared to the rest of the regions. The stimulant effect of noradrenaline was completely abolished following pretreatment of the tissue with the non-competitive alpha receptor blocker phenoxybenzamine in all regions of the ureter. Figure 3 shows that selective blockade of alpha-l receptors with prazosin 10-6 M did not inhibit the action of noradrenaline in the uretero-pelvic and proximal regions whereas pretreatment with yohimbine 10-6 M, a selective alpha-2 receptor blocker, abolished the effect of noradrenaline and resulted in the relaxation of the ureter. Figure 4 shows that alpha-2 agonist clonidine caused a statistically significant rise in amplitude of contraction of the uretero-pelvic and proximal part of the ureter strips after blocking the alpha-1 receptors with prazosin 10-6 M while phenylephrine, a selective alpha-1 agonist, did not produce any significant enhancement in ureteral contractions of the ureteral strips pretreated with yohimbine 10-6 M compared to controls. Figure 5 shows the effect of clonidine on the ureterovesicular regions of the ureter after blocking the alpha-1 receptors with prazosin 10-6 M.
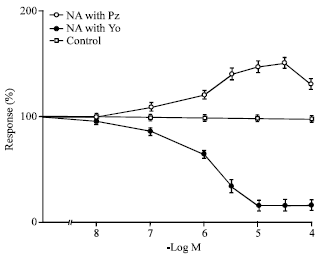 | |
| Fig. 3: | Dose-response curves to nor-adrenaline on uretero-pelvic and proximal strips after blocking alpha-1 receptors with prazosin and alpha-2 receptors with yohimbine represented by open and closed circles respectively (n = 10). Open squares represent the corresponding control values and 100 % denotes no change in control situation (n = 10). All concentrations of noradrenaline from 10-6 to 10-4 M are highly significant p<0.001, for 10-7 M, p<0.05, 10-8 M, p is insignificant compared to controls. Mean value±SE are given; n: No. of experiments |
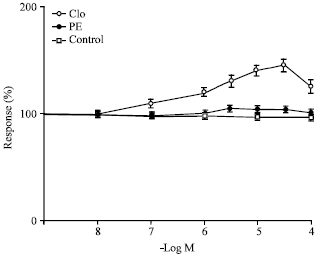 | |
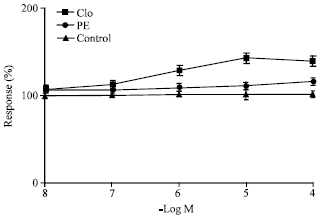
| Fig. 4: | Effect of clonidine and phenylephrine on uretero-pelvic and proximal strips represented by open and closed circles respectively (n = 10). Open squares represent the corresponding control values (n = 10) and 100 % denotes no change in control parameters. All concentrations of clonidine are highly significant p<0.001 except 10-7 M which is also significantly different p<0.01 compared to controls while all contractions of phenylephrine are not significantly different from controls |
 | |
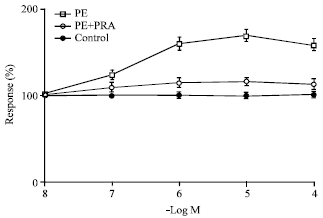
| Fig. 5: | The effect of phenylephrine and clonidine on the ureterovesicular regions of the kidney represented by closed squares and closed circles respectively. Closed triangles represent the corresponding control values (n = 10) and 100% denotes no change in control values. Concentrations of Clonidine from 10¯6-10-4 M are significant p<0.01 compared to controls while concentrations of phenylephrine are not significantly different from controls. Mean value±SE are given; n: No. of experiments |
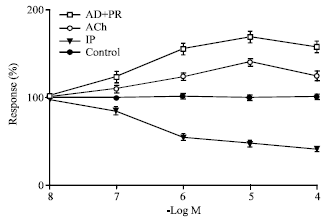
Clonidine shows a statistically significant rise in amplitude of contractions while phenylephrine did not show any significantly different rise in amplitude after pretreating the preparation with yohimbine 10-6 M compared to controls. In Fig. 6 selective alpha-1 agonist phenylephrine show statistically significant contractile response in ureteral regions between pelvic and vesicle regions compared to controls. Pretreatment of alpha-1 receptor blocker prazosin 10-6 M inhibited the phenylephrine induced contractions in these regions compared to controls. Figure 7 shows the effect of adrenaline following pretreatment of the tissue with beta blocker propranolol 10-6 M. Isoprenaline at concentrations of 10-8-10-4 M caused a significant relaxation of ureteral smooth muscle indicating the beta-influence. Acetylcholine showed stimulatory effect on all parts of the ureter which is statistically significant in the pelvic regions, which could be blocked by atropine.
Transmural stimulation of the preparation with high intensity, high frequency, long duration stimuli, subthreshold for the direct stimulation of smooth muscle, increased the force of contraction comparable to the stimulatory effect of noradrenaline. Pretreating the preparation with non-specific alpha receptor blocker phenoxybenzamine (5x10-6 M) decreased slightly the amplitude of contraction on stimulating with high intensity, high frequency and long duration stimuli.
 | |
| Fig. 6: | The effect of phenylephrine and phenylephrine after pretreating the preparation with prazosin on mid-ureteral regions of the kidney represented by open squares and closed circles respectively. Closed circle represents the corresponding control values (n = 10) and 100% denotes no change in control values. Concentrations of Phenylphrine from 10-6-10-4 M are significant p<0.001 compared to controls while concentrations of phenylephrine after pretreatment with prazosin are not significantly different from controls. Mean value±SE are given; n: No. of experiments |
 | |
| Fig. 7: | The effect of adrenaline after pretreatment with propranolol, acetylcholine and isoprenaline on proximal ureter are shown with open squares, open circles and closed triangle respectively. Closed circle represents the corresponding control values (n = 10) and 100% denotes no change in control values. Concentrations of adrenaline from 10-6-10-4 M are highly significant p<0.001 compared to controls. Concentrations of acetylcholine from 10-6-10-4 M are significant p<0.01 and isoprenaline concentrations from 10-7-10-4 M are highly significant p<0.001 compared to controls. Mean value±SE are given; n: No. of experiments |
Stimulating the preparation after blocking the beta receptors with propranolol 10-6 M restored the same intensity of contraction. Stimulating the preparation after blocking cholinergic receptors with atropine (5x10-6) has generated a force of contraction similar to the transmural stimulation of the preparation. Stimulating the tissue in presence of TTX (10-8 M) has completely abolished the force of contraction generated.
DISCUSSION
The in vitro model of the spontaneously contracting sheep ureter is a useful means for the demonstration of drug effects on ureteral smooth muscle. The present study has shown that noradrenaline, increases the force of contractions of the ureter at concentrations of (10-8 to 10-4 M). The reversal of this action by phenoxybenzamine suggests that the excitatory effect of noradrenaline is attributed to alpha receptor stimulation. Adrenaline on the other hand only mimicked noradrenaline in its actions following blockade of beta receptors with propranolol. This observation coupled with the finding that the beta agonist, isoprenaline has a strong relaxant effect of the ureter, has lead to the suggestion that beta receptor stimulation has an inhibitory effect on ureteral smooth muscle. These data suggest the presence of alpha-stimulatory and beta-inhibitory adrenergic receptors in the sheep ureter, which is in accordance to the observation of Weiss et al. (1978) in electrically stimulated preparations of isolated dog ureter and the findings of McLeod et al. (1973) and Rose and Gillenwater (1974) on in vivo ureters.
Electrical stimulation of isolated ureteral segment with high intensity, high frequency, short duration stimuli, which is subthreshold values for direct stimulation of ureteral smooth muscle, releases endogenous neurohumors which enhances the contractions of the ureter. A decrease in the force of contraction observed after blocking the alpha receptors with phenoxybenzamine is attributed to beta effect of noradrenaline which can be blocked by propranolol. Cholinergic receptor blocking with atropine has not affected the force of contraction developed after transmural stimulation. Blocking the neuronal release of neurotransmitters with TTX clearly indicates the presence of innervation in the ureter in consistent with the findings of Weiss et al. (1978).
This study has made possible the further characterisation of the alpha receptor subtype responsible for mediating the stimulant action of noradrenaline in the ureter. The actions of selective alpha-l and alpha-2 receptor agonists and antagonists have shown that only clonidine can mimic the actions of noradrenaline in the pelvic, proximal and vesicular regions and furthermore, in the above regions of the ureter, the actions of noradrenaline can be blocked by the alpha-2 receptor blocker yohimbine and not by the alpha-l receptor blocker prazosin. These observations lead to the conclusion that the alpha-2 receptor subtype is responsible for mediating the stimulatory action of noradrenaline in the pelvic, proximal and vesicle regions of the ureter while alpha-1 receptor population in the rest of the ureter. It is interesting to note that the middle part of the ureter showed only a mild relaxation when alpha-1 receptor was blocked with prazosin to the action of noradrenaline. Furthermore, Phenylephrine, alpha-1 receptor agonist, shows more pronounced stimulatory action on the ureter compared to clonidine in the middle and distal regions of the ureter. Yohimbine, identified as a competitive alpha-2 receptor antagonist in many vascular smooth muscle preparations (De Mey and Vanhoutte, 1981) was shown to non-competitively block alpha receptors in the pelvic and proximal part of the ureter. Earlier studies of the action of yohimbine on rat bladder (Ruffolo et al., 1981) have also shown that blockade of alpha receptors in this tissue is non-competitive. Results of this study suggest that the postsynaptic receptors responsible for the mediation of the stimulatory action of noradrenaline are of the alpha-2 subtype in the pelvic, proximal and vesicular regions of the ureter while alpha-1 in the rest of the ureter.
Earlier studies showing the histological distribution of sympathetic nerves to be essentially in the uretero-pelvic and uretero-vesical region (Duarte-Escalante et al., 1969; Edyvane et al., 1995) conforms with this finding which show greater response to the adrenergic agonists and antagonists predominantly alpha-2 adrenergic subtypes. This study also confirms the presence of mainly alpha-1 receptors in the distal and mid regions of ureter. It has been shown in human urteral studies that the density of alpha-1 adrenoceptor population is greatest in the distal ureter as compared with the proximal regions (Sigala et al., 2005). Alpha-1 adrenoceptor antagonists have been clinically used to treat patients with urolithiasis to promote stone passage and it has been shown highly successful in promoting the passage of distal ureteral calculi (Hollingsworth et al., 2006).
Conventionally, postsynaptic adrenergic receptors are of alpha-l subtype and presynaptic receptors are alpha-2 subtype (Berthelsen and Pettinger, 1977). Investigations in the last few decades have demonstrated several exceptions to this classification and many subtypes of both alpha-1 and alpha-2 receptors were identified and charterised by radioligand binding methods (Bylund and Toews, 1993). The presence of alpha-2 postsynaptic receptors has been described in other tissues such as urinary bladder and urethra (Andersson et al., 1984) feline middle cerebral and lingual arteries (Skarby et al., 1983), canine saphenous vein (De Mey and Vanhoutte, 1981) and in the internal carotid, mesentric, splenic, renal and femoral vascular beds of the dog (Polonia et al., 1986).
In our earlier studies prostaglandin synthesis has been shown to be a contributing factor for the propagation and transmission of ureteral contractions in vitro (Thulesius et al., 1986) and treatment of isolated ureteral rings with indomethacin has been shown to result in the complete cessation of ureteral activity (Thulesius and Angelo-Khattar, 1985; Angelo-Khattar et al., 1985). However, prostaglandin synthesis inhibitors on the intact sheep ureter in vivo, only partially suppressed contractile activity (Angelo-Khattar and Thulesius, 1985). Therefore, we speculate that both neurogenic and myogenic components play a vital role in the normal functioning of the ureter. The contractile activity is dependent to some extent on the nervous stimulation of, in particular, the pelvic region and the propagation and the rhythmicity of contraction is influenced by the local synthesis of prostaglandins and neurohumoral agents acting on specific postsynaptic alpha-1 and alpha-2 receptors.
The summary of the finding suggest that neurogenic contractions of the ureter is initiated by the release of noradrenaline affecting predominantly on the alpha-2 adrenergic receptors in the pelvic and vesicle part of the kidney and Alpha-1 adrenergic receptors in the rest of the regions. From the functional point of view control of ureteral peristalsis is an important factor in the management urolithiasis to the benefit of the patients in managing pain and stone removal. The new findings in ovine ureter suggest that similar in vitro studies with human ureter is essential to determine the efficacy of the alpha-1 adrenergic receptor blockers clinically used to relax the ureter for the ureteral stone removal.
ACKNOWLEDGMENTS
The authors gratefully acknowledges the excellent co-operation and support received from the colleagues at the Faculty of Medicine, Kuwait University, Kuwait; Faculty of Medicine, UAE University, Al Ain, United Arab Emirates; Department of Health Sciences, Higher Colleges of Technology, Dubai and extend special thanks to Prof. Olav Thulesius, Dr. Maria Angelo-Khattar, Dr. Leon Joesph and Dr. Peter Alomaka for all the related earlier studies.
REFERENCES
- Andersson, K.E., B. Larsson and C. Sjogren, 1984. Characterisation of the alpha-adrenoceptors in the female rabbit urethera. Br. J. Pharmacol., 81: 293-300.
Direct Link - Angelo-Khattar, M., Thulesius, O. and T. Cherian, 1989. The effect of glucocorticosteroids on in vitro motility of the ureter of the sheeep. Br. J. Pharmacol., 96: 527-530.
Direct Link - Angelo-Khattar, M., O. Thulesius, T. Nilsson, T. Cherian and L. Joseph, 1985. Motility of the human ureter with special reference to the effect of Indomethacin. Scand. J. Urol. Nephrol., 19: 261-265.
Direct Link - Arunlakshana, O. and H.O. Schild, 1959. Some quantitative uses of drug antagonists. Br. J. Pharmacol. Chemother., 14: 48-58.
PubMedDirect Link - Berthelsen, S. and W.A. Pettinger, 1977. A functional basis for classification of alpha adrenergic receptors. Life Sci., 21: 595-606.
PubMed - Bylund, D.B. and M.L. Toews, 1993. Radioligand binding methods: Practical guide and tips. Am. J. Physiol., 9: L421-L429.
Direct Link - Davenport, K., A.G. Timoney and F.X. Keeley Jr., 2007. The role of ureteral relaxation in the promotion of stone passage. Proc. 5th AIP Conf., 900: 243-252.
CrossRefDirect Link - Duarte-Escalante, O., P. Labay and S. Boyarsky, 1969. The neurohistochemistry of mammalian ureter: A new combination of histochemical procedures to demonstrate adrenergic, cholinergic and chromaffm structures in ureter. J. Urol., 101: 803-811.
PubMed - Hannappel, J. and K. Golenhofen, 1974. The effect of catecholamines on ureteral peristalsisin different species (dog, guinea-pig and rat). Pflugers Arch. Eur. J. Physiol., 350: 55-68.
CrossRefDirect Link - Hernandez, M., D. Prieto, U. Simonsen, L. Rivera, M.V. Barahona and A. Garcia-Sacristan, 1992. Noradrenaline modulates smooth muscle activity of the isolated intravesical ureter of the pig through different types of adrenoceptors. Br. J. Pharmacol., 107: 924-931.
PubMed - Hollingsworth, J.M., M.M. Davis, B.T. West, J.S. Wolf Jr. and B.K. Hollenbeck, 2009. Trends in medical expulsive therapy use for urinary stone disease in U.S. emergency departments. J. Urol., 74: 1206-1209.
PubMed - Hollingsworth, J.M., M.A.M. Rogers, S.R. Kaufman, T.J. Bradford, S. Saint, J.T. Wei and B.K. Hollenbeck, 2006. Medical therapy to facilitate urinary stone passage: A Meta-analysis. Lancet, 368: 1171-1179.
PubMed - Edyvane, K.A., P.J. Smet, J. Jonavicius and V.R. Marshall, 1995. Regional differences in the innervation of the Human ureterovesical junction by the tyrosine hydroxylase, vasoactive intestinal peptide and neuropeptide y-like immunoreactive nerves. J. Urol., 154: 262-268.
PubMed - Ohki, K., O. Yosihiro and K. Suzuki, 2010. The investigation of ureteral sympathetic innervations, using semi-serial sections: Why does the α1-adrenergic receptor antagonist work well for ureteral stones? Int. Urol. Nephrol., 42: 113-117.
CrossRefDirect Link - Mayo, M.E. and S.A. Halbert, 1981. The effect of autonomic drugs on ureteric peristalsis: A canine in vivo study. Urol. Res., 9: 209-216.
CrossRefDirect Link - McLeod, D.G., D.G. Reynolds and K.G. Swan, 1973. Adrenergic mechanisms in the canine ureter. Am. J. Physiol., 224: 1054-1058.
PubMed - Morita, T., 1986. Characteristics of spontaneous contraction and effects of isoproterenol on contractility in isolated rabbit renal pelvic smooth muscle strips. J. Urol., 135: 604-607.
PubMed - Morita, T., M. Ando, K. Kihara and H. Oshima, 1994. Function and distribution of autonomic receptors in canine ureteral smooth muscle. Neurourol. Urodyn., 13: 315-321.
Direct Link - Morita, T., I. Wada, T. Suzuki and S. Tsuchida, 1987. Characterisation of α-adrenoceptor subtypes involved in regulation of ureteral fluid transport. Tohoku J. Exp. Med., 152: 111-118.
PubMed - Nakada, S.Y., T.L.C. Coyle, M.K. Ankem, T.D. Moon and T.D. Jerde, 2007. Doxazosin relaxes ureteral smooth muscle and inhibits epinephrine-induced ureteral contractility in vitro. J. Urol., 70: 817-821.
Direct Link - Prieto, D., M. Hernandez, L. Rivera, E. Ordaz and A. Garcia-Sacristan, 1993. Catecholaminergic innervations of the equine ureter. Res. Vet. Sci., 54: 312-318.
PubMed - Prieto, D., U. Simonsen, J. Martin, M. Hernandez and L. Rivera et al., 1994. Histochemical and functional evidence for cholinergic innervations of the equine ureter. J Auton. Nervous Syst., 47: 159-170.
CrossRef - Rose, J.G. and J.Y. Gillenwater, 1974. The effect of adrenergic and cholinergic agents and their blockers upon ureteral activity. Invest. Urol., 11: 439-451.
PubMed - Ruffolo, Jr., R.R., J.E. Waddell and E.L. Yaden, 1981. Postsynaptic alpha adrenergic receptor subtypes differentiated by Yohimbine in tissues from the rat: Existence of alpha-2 adrenergic receptors in rat aorta. J. Pharmacol. Exp. Ther., 217: 235-240.
Direct Link - Sigala, S., M. Dellabella, G. Milanese, S. Fornari and S. Faccoli et al., 2005. Evidence for the presence of alpha1 adrenoceptor subtypes in the human ureter. Neurourol. Urodyn., 24: 142-148.
CrossRefDirect Link - Thulesius, O. and M. Angelo-Khattar, 1985. The effect of indomethacin on the motility of isolated sheep ureter. Acta Pharmacol. Toxicol., 56: 298-301.
CrossRefDirect Link - Thulesius, O., L. Ugaily-Thulesius and M. Angelo-Khattar, 1986. Generation and transmission of ovine ureteral contractions with special reference to prostaglandins. Acta Physiol. Scand., 127: 485-490.
CrossRefDirect Link - Tindall, A.R., 1972. Preliminary observations on the mechanical and electricalactivity of the rat ureter. J. Physiol. (Load.)., 223: 633-647.
Direct Link - Weiss, R.M., A.L. Bassett and B.F. Hoffinan, 1978. Adrenergic innervation of the ureter. Invest. Urol., 16: 123-127.
PubMed - Wheeler, M.A., Y.H. Cho, K.W. Hong and R.M. Weiss, 1990. Age-dependent alterations in beta-adrenergic receptor function in guinea pig ureter. Prog. Clin. Biol. Res., 327: 711-715.
PubMed