
E. A. Hussein
Department of Biology, Faculty of Science, Ibb University, Ibb, Yemen
A. M. Taj-Eldeen
Center of Genetic Resources, Faculty of Agriculture, Sana`a University, Sana a, Yemen
A. S. Al-Zubairi
Department of Biochemistry and Molecular Biology, Faculty of Medicine and Health Sciences, University of Sana`a, Yemen
A. S. Elhakimi
Center of Genetic Resources, Faculty of Agriculture, Sana`a University, Sana a, Yemen
A. R. Al-Dubaie
Department of Biology, Faculty of Science, Sana a University, Sana`a, Yemen
International Journal of Pharmacology
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 464-471







ABSTRACT
In the present study, callus was successfully obtained from hypocotyl explants of Brassica nigra (L.) in vitro under both dark and light incubation conditions separately using Murashige and Skoog’s basal salts enriched with sucrose, benzyladenine (BA), Naphthalene Acetic Acid (NAA) and 2,4-Dichlorophenoxyacetic acid (2,4 D). Callus growth curve was plotted on the basis of single callus dry weight in grams versus time in weeks. The obtained calli were subjected to phytochemical screening for some secondary plant metabolites using thin layer chromatography, determination of total phenolics using Folin Ciocalteu reagent, estimation of the antioxidant activity by the scavenging activity of DPPH free radicals and screening calli extracts for antibacterial activity against 4 pathogenic bacterial strains. Except for phytochemical screening which was carried out at the end of the growth period only, other analyses were carried out when calli were 2, 4 and 6 weeks old. Results of the present study have revealed that calli were rich in secondary metabolites as they gave positive results for testes of volatile oils, anthraquinones, flavonoids and tannins. Although there were some exceptions, it was generally observed that total phenolic, antioxidant activity and antibacterial activity were higher in calli obtained under light incubation conditions than calli obtained under dark incubation conditions or the mother plant parts from which calli were induced. It is also observed that older calli accumulated more amounts of total phenolics, exhibited higher antioxidant activity and stronger antibacterial activity.
PDF Abstract XML References Citation
Received: March 04, 2010;
Accepted: May 06, 2010;
Published: September 02, 2010
How to cite this article
E. A. Hussein, A. M. Taj-Eldeen, A. S. Al-Zubairi, A. S. Elhakimi and A. R. Al-Dubaie, 2010. Phytochemical Screening, Total Phenolics and Antioxidant and Antibacterial Activities of Callus from Brassica nigra L. Hypocotyl Explants. International Journal of Pharmacology, 6: 464-471.
DOI: 10.3923/ijp.2010.464.471
URL: https://scialert.net/abstract/?doi=ijp.2010.464.471
DOI: 10.3923/ijp.2010.464.471
URL: https://scialert.net/abstract/?doi=ijp.2010.464.471
INTRODUCTION
Because of the many scientific, economic and ecological advantages of plant tissue culture, it is nowadays considered an important strategy for in vitro production of bioactive compounds for drug and food industries (Bougard et al., 2001; Mulabagal et al., 2004; Srivastava and Srivastava, 2007). Phenolics, antioxidants and antibacterial compounds which are dealt with in this study, are needed for both types of industries. Plant phenolics are modulators of indol acetic acid catabolism and play vital role in plant life. Phenolics range from simple low molecular weight single ringed compounds to complex tannins and derived polyphenols. Many plant phenolics exhibit antibacterial properties (Kefeli et al., 2003) and a relatively fair correlation between phenolic content of plant extracts and their antioxidant activities has been also reported (Kaur and Kapoor, 2002; Ivanova et al., 2005; Farrukh et al., 2006).
An antioxidant is a molecule stable enough to donate electron to a rampaging free radical or Reactive Oxygen Species (ROS), neutralize it and reduce its capacity to damage living cells and this is why antioxidants are used to prevent deleterious consequences of oxidative stress on human (Halliwell and Gutteridge, 1998). The majority of herbal antioxidant active compounds are phenolics, flavonoids, flavones, isoflavones, anthocyanins, caumarins, lignans, catechins and isocatechins (Larson, 1988; Maharik et al., 2009; Kaur and Kapoor, 2002; Prior, 2003; Cai et al., 2004; Chye and Sim, 2009). On the other hand antibacterial resistance is a world-wide growing problem (WHO, 2001; Cohen, 2000). Bonjar (2004) reported that one of the measures to combat the increasing rate of resistance on the long run is to have continuous investigation for new, safe and effective antibacterial compounds.
Callus and cell suspension cultures of B. nigra were started in previous studies (Vibha et al., 1990; Gerdemann-Knorck et al., 1994) with the purpose of plantlet regeneration. The objectives of this study was to induce callus formation from hypocotyl explants of Brassica nigra L. (black mustard) under both dark and light incubation conditions in vitro and to detect the potential of calli to produce active secondary metabolites and exhibit biological activities. The calli extracts were subjected to TLC phytochemical screening, analysis of total phenolics, assay of free radical scavenging and antibacterial activities.
MATERIALS AND METHODS
Explant preparation: Seeds of Brassica nigra (L.), black mustard were bought from Sana’a City, Yemen and the work was done during the years of 2008-2009. The seeds were surface sterilized according to Chawla (2003) by submerging the explants into a solution of 70% ethanol with continuous and gentile stirring for 1 min. The seeds were transferred to 100 mL conical flask containing 20% solution of sodium hypochlorite (1% active chlorine) with continuous gentle stirring for 7 min. Then the seeds were washed with 3 successive rinses of sterile distilled water under aseptic conditions and dried between two layers of sterile filter papers in a Petri-dish. Using sterile scalpel the seeds were transferred to Petri-dishes containing hormone free culture media (Murashige and Skoog, 1962) and were allowed to germinate for 1 week at room temperature. Hypocotyls were then cut under aseptic conditions into 1 cm length explants for callus initiation.
Callus initiation: For induction of callus from hypocotyls explants, Murashige and Skoog’s culture media (Murashige and Skoog, 1962) containing 3% sucrose supplemented with various combinations of Benzyl Adenine (BA) and a mixture of Naphthalene Acetic Acid (NAA) and 2,4-Dichlorophenoxyacetic acid (2,4 D) (1:1). The concentrations used of each growth substance were 0.0, 0.1, 0.5, 1.0, 1.5, 2.0 and 2.5 mg L-1, accordingly a total of 49 different hormonal combinations were prepared. The pH value of the culture media was adjusted to 5.7 with 0.8% (w/v) agar-agar (Glaxo, bacteriological grade) prior to autoclaving. Culture media were dispensed into sterile petri dishes. Each treatment was represented by 6 replicates and each replicate contained 3 hypocotyl explants. The cultures were divided into two groups, the first of which was incubated at room temperature in light while the second was incubated in dark. Morphogenic responses were recorded after 6 weeks and the optimum hormonal combination for callus induction was determined.
For production of callus, explants were cultured on the same culture media supplemented with 2.5 mg L-1 BA+0.5 mg L-1 (NAA/ 2,4 D mixture). Plates were divided into two groups, the first of which was incubated in dark while the other was incubated under continuous light illumination. Calli were harvested after 2, 4 and 6 weeks of growth to study the total phenolics content, the antioxidant and antibacterial activities.
Growth curve under light and dark incubation conditions: To measure the callus growth, hypocotyl explants were cultured on MS culture medium supplemented with BA (2.5 mg L-1) and NAA/2,4 D mixture (0.5 mg L-1). Calli were harvested weekly until 8 weeks of incubation ( in dark and/or light conditions, 2000 lux) and a growth curve was plotted (dry weight/ single callus in mg) versus time in weeks.
Phytochemical analysis using TLC: Thin layer chromatographic screening of some plant secondary metabolites (volatile oils, alkaloids, anthraquinone glycosides, saponins and flavonoids) was carried out on a mixture of powder of calli collected from first, second and third stages of growth and it was carried out as described by Macek (1972) using precoated Silica gel G 60 F254 plates (Merk). Detection was carried out using visible light, short and long UV lights before and after spraying with specific reagents. Colors and Rf values of detectable spots were recorded. The mobile phases and spray reagents used are shown in Table 1.
Determination of total phenolics: The amount of total phenolics in extract of the control tissue (hypocotyl segments) and calli produced under dark and light incubation conditions were determined according to the method of Spanos and Wrolstad (1990) using Folin-Ciocalteu reagent after 2, 4 and 6 weeks of callus growth. A known weight of extract was dissolved in 10 mL of methanol. To 5 mL of each sample, 0.25 mL of 10% diluted Folin-Ciocalteu reagent was added and the resulting mixture was incubated at 25°C for 15 min. Absorbance of the resulted color was measured spectrophotometrically at 765 nm. A standard curve was made using different concentrations of gallic acid and the concentrations in the tested extracts were expressed as milligram gallic acid equivalent per gram dry weight of plant material powder (mg GAE g-1 dw).
| Table 1: | The mobile phases and their spray reagents used to detect the phytochemical classes |
 | |
Free radical scavenging assay: Diphenylpiacrylhydrazyl (DPPH) is a molecule containing a stable free radical. In the presence of an antioxidant which can donate an electron to DPPH, the purple color which is typical to free DPPH radical decays and the change in the absorbency at 517 nm is followed spectrophotometrically. Quantitative estimation was carried out according to the method of (Brand-Williams et al., 1995). The reaction mixture contained 500 μL of test extract and 125 μL of DPPH (DPPH reagent is prepared by dissolving 5 mg of DPPH in 10 mL ethanol) and 375 μL solvent (methanol). Different concentrations of test samples (10, 50, 100, 500 and 1000 μg mL-1) were prepared. The reaction mixtures were taken in eppendorf tubes and incubated at 37°C for 30 min, the absorbance was measured at 517 nm. Pure methanol was used instead of plant extract to prepare blank and ascorbic acid was used as a positive control. Antioxidant activity as measured by the percentage of free radical scavenging activity was calculated from the following Eq. 1:
| (1) |
Antibacterial activity test
Preparation of callus extracts: Calli (2, 4 and 6 weeks old) were harvested and their fresh weights were recorded. Calli were then dried in hot air oven at 50°C. After constant weights were attained, calli were grinded into a fine powder. One gram of dry fine calli powder was soaked in 70% ethanol (150 mL) at room temperature for 1 week with stirring from time to time. After that, filtration was carried out and the residue was washed with 3 successive rinses (100 mL) of ethanol 80%. The filtrate and washings were combined and evaporated to dryness, weighed and then redissolved in 1 mL of 80% ethanol.
Test microorganisms and assay of antibacterial activity: The bacterial strains used in this study were Escherichia coli (ATTCC 11229), Pseudomonas auruginosa (CS.25), Klebsiella pneumonia (ATCC 13883) and Staphylococcus aureus (NCTC7447). Assay of antibacterial activity was carried out by the disk-diffusion method described by Salie et al. (1996). Five millimeter (diameter) filter paper disks were allowed to imbibe different aliquots of ethanolic extract in order to have an exact amount of dry callus extract equal to 25 mg disc-1. After the organic solvent was completely evaporated, discs were put on the surface of nutrient agar seeded with test bacteria in 9 cm diameter Petri-dishes. All the plates were incubated at 37°C for 24 h. The experiment was repeated 3 times under strict aseptic conditions. Antibacterial activity was determined by measuring the diameter of the inhibition zone and the mean values were calculated.
Statistics: Statistical analysis was performed using SPSS statistical package software Version 15.0 (USA). The results were presented as Mean±SEM and all parameters were analyzed by using student’s t-test. The data was considered as significant, if the p-value is p<0.05.
RESULTS
Results of the present study have revealed that after 6 weeks of incubation (Photoperiod 16/8) in vitro, 33 of the 49 used hormonal combinations were able to induce callus formation from B. nigra hypocotyl explants in vitro. The weights of calli obtained differed from one treatment to another. The lowest callus biomass was obtained with the hormonal combination (BA 1.5 mg L-1+NAA/2,4 D mix. 1.5 mg L-1) and it was about 117±20 mg/explant while the heaviest callus was obtained using the hormonal treatment (BA 2.5 mg L-1+NAA/2,4 D mix. 0.5 mg L-1) where the dry callus biomass reached up to 1040±18 mg/explants. One of the four weeks old calli is shown in Fig. 1, it is yellowish whitish, soft and nodular surfaced. The calli is (as seen under 40-X light microscope) composed of almost spherical cells showing numerous discoid green chloroplasts (Fig. 2).
Growth of calli, as measured by fresh and dry weight per single callus under both dark and light incubation conditions over a period of 8 weeks is shown in Fig. 3. Growth of calli in both incubation conditions was slow in the beginning, then gradually and exponentially increased and then gradually decreased. By the end of the growth period (8 weeks), calli which were grown under dark incubation conditions could accumulate dry biomass of about 0.73 g/single callus while under light incubation conditions this amount reached up to 1 g per callus.
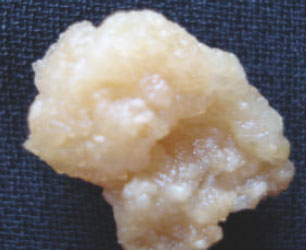 | |
| Fig. 1: | Callus obtained from B.nigra hypocotyl explants after 4 weeks under photoperiod (16/8) incubation conditions |
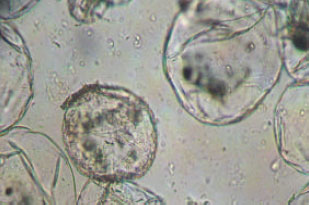 | |
| Fig. 2: | Cells of callus of B. nigra. Almost spherical with numerous discoid chloroplasts |
Phytochemical analysis have shown, in general, that control tissues (hypocotyls explants) contained secondary metabolites belonging to various phytochemical classes. Calli, whether produced under light or dark incubation conditions were able to synthesize various secondary metabolites too, however, differences and similarities between control and calli tissues existed. There were also some differences when thin layer chromatographic plates were inspected under normal day light, short and long UV lights. The largest number of spots which could be detected was obtained under long UV light. Twenty spots were detected in control tissue (2 volatile oils, 3 alkaloids, 5 anthraquinone glycosides, 3 saponins, 4 tannins and 3 flavonoids), 11 spots were detected in dark induced calli (3 volatile oils, 2 alkaloids, 3 anthraquinone glycosides, 2 tannins and 1 flavonoid, while saponins were absent) and 7 spots could be detected in light induced calli (1 alkaloid, 2 anthraquinone glycosides, 3 tannins and 1 flavonoid, while volatile oils and saponins were absent).
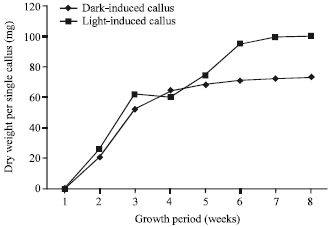 | |
| Fig. 3: | Growth curve of callus of B. nigra |
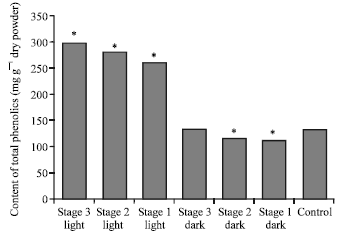 | |
| Fig. 4: | Total phenolics (mg g-1 dry powder) of various B. nigra plant materials. *Significant value |
Results of total phenolics in dark obtained calli (Fig. 4) have shown that (stage I and II) were significantly lower (p<0.05) than control tissue (131.67±2.73), while it could gradually accumulate phenolics in stage 3 almost equal to the control tissues (about 132.33±1.45 mg g-1). Calli which were produced under continuous illumination accumulated significantly higher amounts of total phenolics (p<0.05) compared to control tissue as well as corresponding calli which were obtained under dark incubation. Amount of total phenolics in stage III analysis reached up to 297.00±1.53 mg g-1, i.e., more than two folds higher (p<0.05) than the content in control tissue.
Antioxidant activity as expressed by the percentage of DPPH free radical scavenging activity (Table 2) in both types of calli increased with the increase in age of culture. In addition, the light induced calli were more active in the synthesis of antioxidant compounds as they were significantly higher (p<0.05) than both the control tissue and the corresponding calli produced in dark in almost all the concentrations used. Calli which were obtained under dark conditions could synthesize (especially in the analysis of stage III) significantly higher amounts of antioxidant components than the control tissue.
| Table 2: | Free radical scavenging activities (%) of methanolic extracts of various B. nigra plant materials |
 | |
| *p<0.05. Each value is a mean of 3 determinations ±SE | |
| Table 3: | Antibacterial activity of extracts of various plant materials of B. nigra as detected by the disk plate diffusion assay |
 | |
| Diameter of inhibition zones are means of three determinations measured in mm | |
All the tested tissues whether control or calli were found to have antibacterial activity against all the pathogenic bacteria tested with various degrees (Table 3), the lowest effect (inhibition halo = 6 mm) was obtained with extract of stage I dark induced callus while maximum inhibition zone (15 mm) was obtained with extract of stage III light induced callus against E. coli. Extracts of dark induced calli exhibited lower effect on the pathogenic bacteria tested than control while inhibition halos resulted from extracts of calli produced under light were wider than control.
DISCUSSION
Hormonal combination of 2.5 mg L-1 BA+0.5 mg L-1 (mix of NAA and 2,4 D) was used to study growth of calli because this treatment was found to produce maximum callus dry biomass (1040 mg/explant). Light might have enhanced photosynthetic pigment formation and this may be the reason that biomass of calli which were grown under light incubation conditions accumulated higher amounts of biomass than calli were grown under dark incubation conditions. Light was required for Lemna gibba callus induction and growth (Moon and Stomp, 1997).
In this study, volatile oils were detected in both control tissue (hypocotyls segments) and dark induced calli. It has been reported that volatile oils were difficult to be formed in undifferentiated callus tissues because it needs specialized botanical structures like glands (Sevo�n and Oksmann-Caldentey, 2002). However, glands are places for storage of volatile oils and biosynthetic potential depends on genetic information retained in the cells, whether differentiated or undifferentiated. Volatile oils however were detected in callus cultures of Ruta graveolens, Perilla species (Ohnishi et al., 1983) and Rosmarinus officinalis (Youssef and Rady, 2000). In this study, calli obtained under dark incubation conditions contained volatile oils while light induced calli did not, although these calli showed better total phenolics content, higher antioxidant activity and stronger antibacterial effects. It has been previously mentioned that five weeks old green calli of Rosmarinus officinalis, for example, were observed to synthesize carnosic acid (one of the active principles of rosemary) while corresponding non-green calli could not (Caruso et al., 2000). Very recently, it has been reported by Shinde et al. (2010) that cell cultures of Psoralea coraylifolia grown under continuous illumination conditions produced several more folds isoflavones daidzein and genistein than that of field grown plants. However, present results were found to be on the same line supporting the previously published findings reported that growth under light illumination produced more secondary metabolites.
Earlier phytochemical investigations have proved that, in many cases, plant cells or tissues cultured in vitro are able to synthesize compounds belonging to a diversity of classes of plant secondary metabolites like alkaloids, flavonoids, caumarins, glycosides, terpenoids, phenolics and many others (Khafagi, 2007; Priya and Patric, 2008). It has been demonstrated that optimization of the cultural conditions can increase the content of a specific metabolite in the cultured cells many folds than the concentration in the parent intact whole plant (Verpoorte et al., 1994) and even induce the cultured cells to synthesize metabolites which could not be detected in the wild parent plant. Accordingly our results showed that maximum calli growth was observed with hormonal concentrations of BA 2.5 mg L-1+NAA/2,4 D mix. 0.5 mg L-1. Biosynthesis of phenolics in plant cell cultures has been detected and reported in previous studies (Angelova et al., 2001; Bahorun et al., 2005). It has been observed that calli which were obtained in vitro accumulated significant amounts of total phenolics in comparison to the parent plant parts phenolics (Cvikrova et al., 1990; Daayef et al., 2003).
Significant antioxidant activities were detected in this study as well as in extracts of calli obtained in vitro from explants of many other plants like Ipomea aquatica Forsk. (Prasad et al., 2006), Stevia rebaudiana (Tadhani et al., 2007), Crataegus sinaica Boiss. (Maharik et al., 2009) and Psoralea corylifolia (Shinde et al., 2010). In comparison to corresponding plant parts from which explants were taken for induction of callus formation in vitro, the obtained calli were observed to exhibit higher antioxidant activity in some cases (Maharik et al., 2009; Shinde et al., 2010). An antioxidant may belong to any of the classes of secondary metabolites which were detected in this study as it has been mentioned that the majority of herbal antioxidant active compounds are phenolics, flavonoids, flavones, isoflavones, anthocyanins, caumarins, lignans, catechins and isocatechins (Larson, 1988; Maharik et al., 2009; Kaur and Kapoor, 2002; Prior, 2003; Cai et al., 2004; Lila, 2004). Jacob and Malpathak (2004) reported that the plant cell cultures under illumination conditions may enhance photosynthesis which can generate intermediate compounds not included in the culture media but needed for improvement of primary and secondary metabolism as well. This may explain the higher antioxidant activity which was observed in this study in extracts of calli induced under continuous illumination conditions in which the total phenolics content were significantly higher under light illumination than that incubated in the dark.
A direct correlation between total phenolics, percentage of free radical scavenging activity and antibacterial activity may exist. It is reported that many of the phenolic contents in plant cells exhibit antibacterial activity (Prior et al., 1998; Kefeli et al., 2003). Significantly higher antibacterial activity has been observed in calli extracts than extracts of intact plant parts (Priya and Patric, 2008; Premjit and Premjit, 2009). The antibacterial activity observed in this study may come back also to the potential of antioxidant activity of callus extracts because many of the phenolic antioxidants have broad spectrum antibacterial properties. Santos et al. (2003) declared that some plant cell cultures can synthesize phenolic antioxidants like phenolic acids, flavonoids and phenolic diterpenes. This may explain the fair correlation observed in this study between total phenolics, antioxidant activity and antibacterial activity.
CONCLUSION
The present study through some light on the possibility of obtaining callus from hypocotyls explants of B. nigra in vitro and indicate that callus tissue can synthesize secondary metabolites belonging to various phytochemical classes and that callus extracts exhibited antioxidant and antibacterial activities. Although, there were some exceptions, this study also showed that obtaining callus under light incubation condition enhances callus growth and production of phenolics to a level exceeding the dark induced calli and mother plant parts. A fair correlation between the content of total phenolics, free radical scavengers and antibacterial activity was also observed.
REFERENCES
- Angelova, Y., S. Petkova and E. Zozikova, 2001. Effect of kinetin and 4PU-30 on the growth and the contents of polyphenol in tobacoo cullus cultures. Bulgarian J. Plant Physiol., 27: 36-42.
Direct Link - Aqil, F., I. Ahmad and Z. Mehmood, 2006. Antioxidant and free radical scavenging properties of twelve traditionally used Indian medicinal plants. Turk. J. Biol., 30: 177-183.
Direct Link - Bahorun, T., V.S. Neergheen and O.I. Aruoma, 2005. Phytochemical constituents of Cassia fistula. Afr. J. Biotechnol., 4: 1530-1540.
CrossRefDirect Link - Bougard, F., A. Gravot, S. Milesi and E. Gontier, 2001. Production of plant secondary metabolites: A historical perspective. Plant Sci., 161: 839-851.
CrossRef - Brand-Williams, W., M.E. Cuvelier and C. Berset, 1995. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol., 28: 25-30.
CrossRefDirect Link - Cai, Y., Q. Luo, M. Sun and H. Corke, 2004. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci., 74: 2157-2184.
CrossRefPubMedDirect Link - Caruso, J.L., J. Callahan, C. de Chant, K. Jayasimhulu and G.D. Winget, 2000. Carnosic acid in green callus and regenerated shoots of Rosmarinus officinalis. Plant Cell Rep., 19: 500-503.
CrossRef - Chye, F.Y. and K.Y. Sim, 2009. Antioxidative and antibacterial activities of Pangium edule seed extracts. Int. J. Pharmacol., 5: 285-297.
CrossRefDirect Link - Cohen, M.L., 2000. Changing patterns of infectious disease. Nature, 406: 762-767.
CrossRefDirect Link - Cvikrova, M., L. Meravy, M. Hurbcova and J. Eder, 1990. Dark induced hanges in the content of phenolic acids in callus culture of alfalfa ( Medicago sativa L.). Biol. Plant., 32: 161-170.
CrossRef - Gerdemann-Knorck, M., M. Sacristan, C. Braatz and O. Schieder, 1994. Utilizatipon of asymmetric somatic hyberdizatioin for the transfere of disease resistance from Brassica nigra to Brassica napus. Plant Breed., 113: 106-113.
CrossRef - Ivanova, D., D. Gerova, T. Chervenkov and T. Yankova, 2005. Polyphenols and antioxidant capacity of Bulgarian medicinal plants. J. Ethnopharmacol., 96: 145-150.
CrossRefPubMedDirect Link - Jacob, A. and N. Malpathak, 2004. Green hairy root cultures of Solanum khasianum Clark: A new route to in vitro solasodine production. Curr. Sci., 87: 1442-1447.
Direct Link - Kaur, C. and H.C. Kapoor, 2002. Anti-oxidant activity and total phenolic content of some Asian vegetables. Int. J. Food Sci. Technol., 37: 153-161.
CrossRefDirect Link - Kefeli, V.I., M.V. Kalevitch and B. Borsari, 2003. Phenolic cycle in plants and environment. J. Cell Mol. Biol., 2: 13-18.
Direct Link - Khafagi, I.K., 2007. Variation of callus induction and active metabolite accumulation in callus cultures of two varieties of Ricinus communis L. Biotechnology, 6: 193-201.
CrossRefDirect Link - Lila, M.A., 2004. Anthocyanins and human health: An in vitro investigative approach. J. Biomed. Biotechnol., 5: 306-313.
CrossRefDirect Link - Maharik, N., S. Elgengaihi and H. Taha, 2009. Anthocyanin production in callus cultures of Crataegus sinaica Boiss. Inter. J. Acad. Res., 1: 30-34.
Direct Link - Moon, H.K. and A.M. Stomp, 1997. Effect of medium components and light on callus induction, growth and frond regeneration in Lemna gibba (Duckweed). In vitro Cellu. Develop. Biol., 33: 20-25.
CrossRef - Vanisree, M., C.Y. Lee, S.F. Lo, S.M. Nalawade, C.Y. Lin and H.S. Tsay, 2004. Studies on the production of some important secondary metabolites from medicinal plants by plant tissue cultures. Bot. Bull. Acad. Sin., 45: 1-22.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Ohnishi, N.K., T. Hirose and H. Sugisawa, 1983. Monoterpene biosynthesis by callus tissues and suspension cells of Pirella species. Phytochemistry, 22: 423-425.
CrossRef - Prasad, N.K., M. Shiva Prasad, S.M. Aradhya and G.R. Shivamurthy, 2006. Callus induction from Ipomoea aquatica Forsk. leaf and its antioxidant activity. Ind. J. Biotechnol., 5: 107-111.
Direct Link - Prior, R.L., G. Cao, A. Martin, E. Sofic and J. McEwen et al., 1998. Antioxidant capacity as influenced by total phenolic and anthocyanin content, maturity and variety of Vaccinium species. J. Agric. Food Chem., 46: 2686-2693.
CrossRefDirect Link - Prior, R.L., 2003. Fruits and vegetables in the prevention of cellular oxidative damage. Am. J. Clin. Nut., 78: 570S-578S.
Direct Link - Priya, J. and R. Patric, 2008. Antibacterial activity studies on Jasminum grandiflorum and Jasminum sambac. Ethnobotani. Leaflets, 12: 481-483.
Direct Link - Salie, F., P.F.K. Eagles and H.M.J. Leng, 1996. Preliminary antimicrobial screening of four South African Asteraceae Species. J. Ethanopharmacol., 52: 27-33.
CrossRef - Sevón, N. and K.M. Oksman-Caldentey, 2002. Agrobacterium rhizogenes-mediated transformation: Root cultures as a source of alkaloids. Planta Med., 68: 859-868.
CrossRefPubMedDirect Link - Bonjar, G.H.S., 2004. Screening for antibacterial properties of some iranian plants against two strains of Escherichia coli. Asian J. Plant Sci., 3: 310-314.
CrossRefDirect Link - Shinde, A.N., N. Malpatak and D.P. Fulzele, 2010. Determination of isoflavone content and antioxidant activity in Psoralea corylifolia L. callus cultures. Food Chem., 118: 128-132.
CrossRef - Srivastava, S. and A. Srivastava, 2007. Hairy root cultures for mass production of high value secondary metabolites. Crit. Rev. Biotechnol., 27: 29-43.
PubMedDirect Link - Tadhani, M.B., V.H. Patel and R. Subhash, 2007. In vitro antioxidant activities of Stevia rebaudiana leaves and callus. J. Food Comp. Anal., 20: 323-329.
CrossRef - Verpoorte, R., R. van der Heijden, H. Hoge and H. Hoopen, 1994. Plant cell biotechnology for the production of secondary metabolites. Pure Applied Chem., 66: 2307-2310.
Direct Link - Vibha, G., A. Abha, V. Jagannathan, V. Gupta and A. Agnihotri, 1990. Plantlet regeneration from callus and protoplasts of Brassica nigra (IC 257) through somatic embryogenesis. Plant Cell Rep., 9: 427-430.
Direct Link - Spanos, G.A. and R.E. Wrolstad, 1990. Influence of processing and storage on the phenolic composition of Thompson Seedless grape juice. J. Agric. Food Chem., 38: 1565-1571.
CrossRefDirect Link - Daayf, F., M. El-Belhaj, M. El-Hassni, F. Jaiti and I. Hadrami, 2003. Elicitation of soluble phenolics in date palm (Phoenix dactylifera L.) callus by Fusarium oxysporum f. sp. albedinis culture medium. Environ. Exp. Bot., 49: 41-47.
CrossRefDirect Link