
Sarah Mousavi
Faculty of Pharmacy and Pharmaceutical Sciences Research Center, Tehran University of Medical Sciences, Tehran 1417614411, Iran
Mojtaba Mojtahedzadeh
Faculty of Pharmacy and Pharmaceutical Sciences Research Center, Tehran University of Medical Sciences, Tehran, Iran
Mohammad Abdollahi
Faculty of Pharmacy and Pharmaceutical Sciences Research Center, Tehran University of Medical Sciences, Tehran, Iran
International Journal of Pharmacology
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 326-337







ABSTRACT
Supraphysiological concentrations of oxygen (hyperoxia) is necessary for treatment or prevention of hypoxemic condition, but hyperoxia through activation of oxidative stress pathways and generation of reactive oxygen species directly or indirectly result in lung injury and dysfunction. Many mediators are involved in the pathways to hyperoxic cell death seeming that transition metal ions especially iron promote the generation of the very reactive free radicals which are damaging to cells. Thus concerning the role of iron, a systematic review was conducted by reviewing all papers found from searching keywords of iron, lung injury, oxidative stress or hyperoxia in bibliography databases. Due to different pathways involved in hyperoxia-induced lung injury, several classes of drugs have been tested for example antioxidants, dexamethasone, pentoxifylline, erythropoietin, lisofylline, sildenafile, N-acetyl cysteine, or prostaglandine but some of them were not only successful but caused significant adverse effects. Findings indicate that metal ions especially iron through catalyzing of most reactive free radicals has a key role in oxidative stress process and hyperoxic condition and thus limitation of iron for prevention of hyperoxia-induced injury seems reasonable. Iron chelation has been recently used for conditions without iron overload such as neurodegenerative, infectious, reperfusion injury, cardioprotection. There are some evidences about positive effects of desferrioxamine alone or in combination when tested in animal models of lung injury or in human but they are not convincing. Further studies are necessary to clarify the importance of intervention with desferrioxamine or new long acting oral agent deferasirox and their risk/benefit in hyperoxia-induced lung injury.
PDF Abstract XML References Citation
How to cite this article
Sarah Mousavi, Mojtaba Mojtahedzadeh and Mohammad Abdollahi, 2010. Place of Iron Chelators Like Desferrioxamine and Deferasirox in Management of Hyperoxia-induced Lung Injury; A Systematic Review. International Journal of Pharmacology, 6: 326-337.
DOI: 10.3923/ijp.2010.326.337
URL: https://scialert.net/abstract/?doi=ijp.2010.326.337
DOI: 10.3923/ijp.2010.326.337
URL: https://scialert.net/abstract/?doi=ijp.2010.326.337
INTRODUCTION
High concentration of oxygen is essential for patients with hypoxic respiratory failure in intensive care unit, but prolonged exposure to oxygen cause tissue damage and lead to acute and chronic lung injury. Hyperoxia through direct oxygen toxicity and accumulation of inflammatory mediators within the lungs result in a process that initiate by damaging to pulmonary epithelium and endothelium and finally lead to pulmonary interstitial fibrosis (Ansari et al., 2008; Vazin et al., 2005a, b). Proliferation of type Π alveolar epithelial cells, destruction of type I alveolar epithelial cell and pulmonary vascular remodeling are examples of changes in hyperoxic condition. No level of oxygen is safe indefinitely; studies of normal individuals exposed experimentally to 100% oxygen at normal pressure have shown evidence of tracheobronchitis and changes in vital capacity, diffusing capacity and lung permeability (Beckett and Wang, 1988; Davis et al., 1983). Inadvertent exposure of patients with normal lungs to prolonged hyperoxia resulted in clinical findings compatible with oxygen toxicity mainly through oxidative stress. Addition of hyperoxia to mechanical ventilation especially with high tidal volume augmented lung injury (Quinn et al., 2002). Different underlying mechanisms are involved in this process whereas oxidative stress is the major one that damage major macromolecules within cells resulting in cell apoptosis or death (Abdollahi et al., 2004). Concerning the importance of oxygen therapy, different drug classes have been tested for treatment or prevention of hyperoxia-induced lung injury specially by authors' team (Kajbaf et al., 2007; Mahmoodpoor et al., 2010; Mojtahedzadeh et al., 2008; Najafi et al., 2009; Salari et al., 2005a, b; Soltan-Sharifi et al., 2007; Vazin et al., 2005a, b).
Neutrophils and macrophages are two major sources of reactive oxygen and nitrogen species (ROS and RNS) in the lung. Two major enzyme systems NADPH oxidase (Babior et al., 2002; Chung-Wai et al., 2003) and nitric oxide synthases (Bogdan, 2001) are responsible for production of ROS or RNS especially superoxide anion, hydrogen peroxide and hydroxyl radical (Abdollahi et al., 2000a, b). Iron as a metal has especial role in production of hydroxyl radical, which can be explained in terms of following reactions (Lloyd et al., 1997):
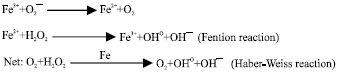 |
The overall reaction is called Haber-weiss reaction (Lloyd et al., 1997). Hydroxyl radical react at diffusion-limited rates with various biomolecules including lipids, proteins, DNA (Nelson and McCord, 1998) and lead to cell injury (Rezaie at al., 2007). So chelation of iron could be a critical event in preventing iron-mediated oxidative stress.
MATERIALS AND METHODS
In order to provide enough data about the subject, we did systematic search utilizing Pubmed, Scopus, Google scholar and Embase databases. The initial search terms were iron, lung injury, oxidative stress or hyperoxia to provide data about the role of iron in hyperoxia-induced lung injury. Then search was continued with hyperoxia, lung injury and treatment as keywords without narrowing or limiting search elements to find the most relevant literatures about the subject. References from each article were also evaluated for relevancy of inclusion in the study. All papers were reviewed to omit irrelevant or duplicate papers. Then their data were extracted into tables and summarized.
RESULTS
As summarized in Table 1, different drug classes have been used for treatment of hyperoxia-induced lung injury. Almost all of the studies were done in animal models, because of the limitation to simulate same clinical condition of high concentrations of oxygen (>95%) for long duration and problems for sampling and analyzing of lung tissue or even bronchoalveolar lavage fluid in human (Cross et al., 1994; Van der Vliet et al., 1999). Furthermore, due to different pathways involved in hyperoxia-induced lung injury, each drug targeted a part of this system (Ahmad et al., 2006; Ciencewicki et al., 2008; Lee and Choi, 2003; Romashko et al., 2003; Sprong et al., 1991).
| Table 1: | Animal studies considering hyperoxia-induced lung injury treatment |
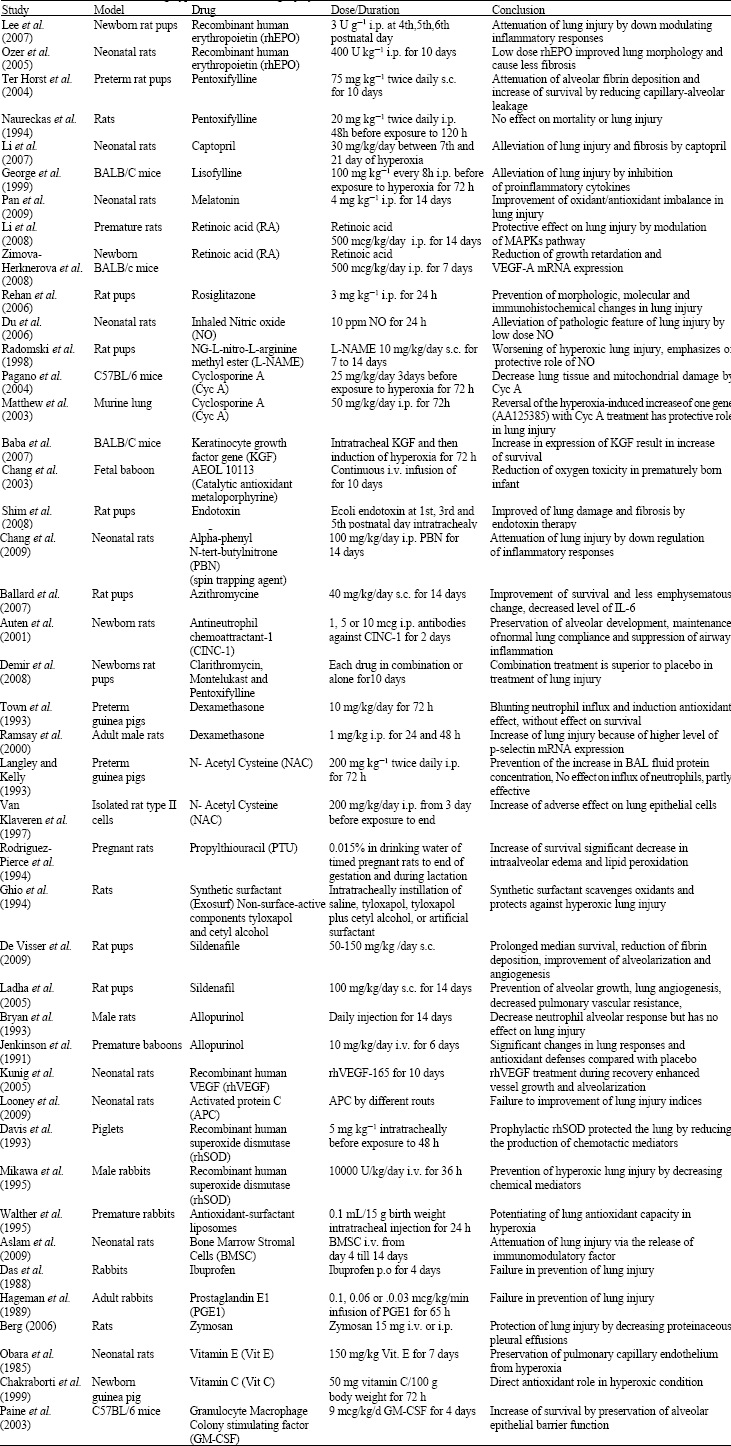 | |
| MPO: i.p.: Intraperitonealy, s.c.: Subcutaneously, i.v.: Intravenous, p.o: Per oral, U:unit, mcg: Microgeram, ppm: Part per million, kg: Kilogeram, h: Hour, d: Day; SOD: Superoxide dismutase, CAT: Catalase, MAPK: Mitogen-activated protein kinasaes, VEGF: Vascular endothelial growth factor | |
Prevention or declining of inflammatory responses and proinflammatory cytokine release such as interleukine 6 or tumor necrosis factor alpha and improvement in antioxidant defenses by increasing activity of antioxidant enzymes such as superoxide dismutase and catalase and also blockade of neutrophil influx are the major mechanisms found for prevention or treatment of oxidative-induced lung injury (Auten et al., 2001; Ballard et al., 2007; Chakraborti et al., 1999; Chang et al., 2003; Davis et al., 1993; George et al., 1999; Mikawa et al., 1995; Obara et al.,1985; Ozer et al., 2005; Rehan et al., 2006; Walther et al., 1995).
Other drugs result in improvement of pulmonary angiogenesis, vessel growth, alveolarization and decreasing pathological lung injury (Kunig et al., 2005; Ladha et al., 2005; Lee et al., 2007; Ozer et al., 2005). Pentoxifylline (Ter Horst et al., 2004), erythropoietin (Ozer et al., 2005), lisofylline (George et al., 1999) and sildenafile (De Visser et al., 2009) are examples of drug that attenuate alveolar fibrin deposition and result in increased survival of animals.
On the other hand some of the drugs especially some commonly used such as N-acetyl cysteine (Langley and Kelly, 1993; Van Klaveren et al., 1997), dexamethasone (Ramsay et al., 2000; Town et al., 1993) or prostaglandine (Hageman et al., 1989) have not been successful in treatment of hyperoxia-induced lung injury but even lead to some adverse effects on epithelial lung cells.
Alteration of pathways that lead to cell apoptosis such as mitogen-activated protein kinases (MAPKs) is another way that prevent lung damage (Hageman et al., 1989; Matthew et al., 2003; Paine et al., 2003; Reynolds et al., 2010; Veness-Meehan et al., 2000; Zimova-Herknerova et al., 2008)
DISCUSSION
Iron as a new target: Iron is uptaken into epithelial cells for sequestration within ferritin. Alveolar macrophages elevate ferritin production after exposure to iron. They posses cell surface receptors specific for both haptoglobin and hemopexin to facilitate their removal (Birgens et al., 1998; Van Snick et al., 1977). Divalent metal transporter 1 (DMT1), Natural resistance-associated macrophage (Nramp) 1,2, transferrin and lactoferrin pathways are involved in iron sequestration in macrophages (Cellier et al., 1995) but continuous exposure overwhelms the capacity of ferritin to sequester the metal and finally releases it (Fig. 1). Ferritin is composed of 24 subunits of heavy (21 KD) and light (19 KD) chains and can reserve up to 4500 atom of Fe3+ ion. H subunit has ferrioxidase activity converting ferric to ferrous forms to fight against oxidative stress. The ratio between H and L subunits differs in different tissue. Increased H-subunit is found in proliferating tissues (Emerit et al., 2001; Harrison and Arosio, 1996). Tumor Necrosis Factor-α and Interleukin-1 transcriptionally induce the H ferritin (Torti and Torti, 2002). Ferritin synthesis is regulated post transcriptionally via the interaction of an iron-regulatory protein (IRP 1 and 2) with Iron Responsive Element (IRE) (Eisenstein, 2000). IRP-1 is an iron-sulfur cluster protein that exists in two forms when iron is abundant. It exists as acytosolic aconitase and IRP-1 that losses its affinity for IRE. The mRNA of ferritin is translated and ferritin synthesis is increased. In the absence of iron, apo IRP-1 put on IRE and the mRNAs of TRF are stabilized and the translation of mRNA of ferritin is suppressed (Papanikolaou and Pantopoulos, 2005; Thomson et al., 1999). IRP-2 protein is abundant in iron insufficiency, but it degrades rapidly in iron excess through targeting of unique 73 amino acid sequence (Lavaute et al., 2001). Nitric oxide and H2O2 activate the IRP-1 through signaling mechanisms and thus mobilize iron from the 4 Fe-4S cubane cluster (Pantopoulos and Hentz, 1998). Down regulation of IRP activity could be a common response to increased formation of H2O2 and O2¯ (Pantarulo, 2005). Iron is imported into cells by transferrin and lactoferrin receptors. Transferrin level as a percentage of total protein in BAL fluid is very high (4-5.6%) compared with values for plasma (Mateos et al., 1998). Transferrin is produced and secreted at high levels in the lung seeming a major extracellular antioxidant in the lung. However, some of the iron carried by transferrin is not sequestered within ferritin, but rather, released into a catalytically active low molecular weight pool which allows the metal to catalyze ROS (Ghio, 2009).
Lactoferrin is structurally and functionally related to transferrin that comes mainly from the airway rather than alveolar region (Mateos et al., 1998), however its localization as sites at which an organism interacts with its environment e.g., secretary epithelium, suggests it plays a role in metal detoxification (Lyer and Lonnerdal, 1993).
Non Transferrin Bound Iron (NTBI) can be transported into cells via transferrin independent pathways and result in the activation of iron responsive proteins including ferritin and lactoferrin.
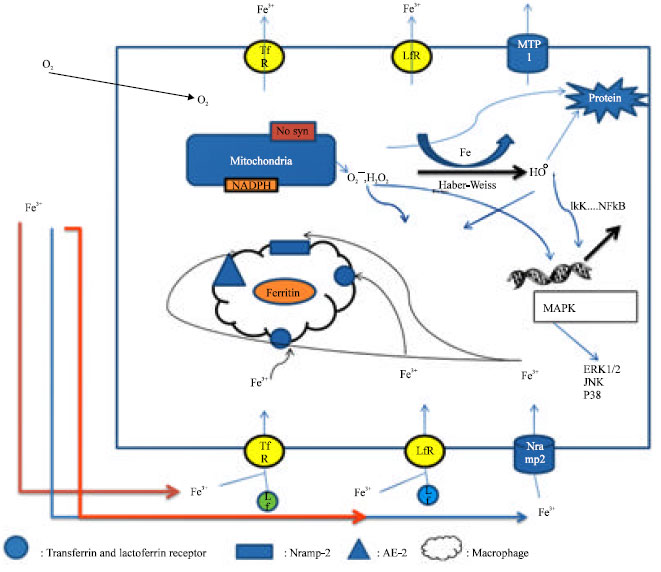 | |
| Fig. 1: | Pathways of iron transport and iron-mediated damage within the lung epithelial cell; Tf: Transferrin, Lf: Lactoferrin, TfR: Transferrin Receptor, LfR: Lactoferrin Receptor, OH0: Hydroxyl radical, H2O2: Hydrogen peroxide, O2¯: Superoxide anion, kappa: IKK: IκB kinase, NFκB : Nuclear Factor κ B, Fe: iron, MAPK: Mitogen-activated Protein Kinase, ERK1/2: Extra-cellular signal-regulated Kinase, JNK: Jun-NH2-terminal Kinase, MTP: Metal Transport Protein, Nramp2: Natural resistance-associated macrophage protein, ROS: Reactive Oxygen Species, AE2: Anion Exchange protein 2, NTBI: Non-Transferrin Bound Iron. Iron has critical role in many cellular cell functions, but it can also generate reactive oxygen species that can damage major macromolecules in cells and result in cell death. So cells have different mechanisms to detoxify iron, the major one, is storage in ferritin. Iron mainly imported to cells by transferrin and lactoferrin receptors, Nramp-2 are responsible of NTBI uptake after ferrireduction of iron. To prevent excessive cellular accumulation, iron exported from cells by means of transferrin and lactoferrin receptors and also MTP-1. Alveolar macrophage through same receptors uptake iron for subsequent storage in ferritin and then releases ferritin in airway to diminish iron stress within cells. Overwhelming of these mechanisms result in excess level of free iron which is through Habber-Weiss reaction produce hydroxyl radical from H2O2 and superoxide anion. These ROS damage directly and indirectly to major macromolecules within cells and through over expression of some genes activate different pathways and protein such as MAPK and IKK pathways that finally lead to cell necrosis and apoptosis |
NTBI uptake is frequently accomplished using Nramp-2 (DMT-1 or DCT-1) (Vidal et al., 1995). mRNA for DMT1increases after lung epithelial cell exposure to iron transcriptional control of IRE isoform, provides a mechanism of regulating Nramp 2 expression to diminish oxidative stress effectively (Lee et al., 1998). Nramp2 usually colocalize with transferrin seeming a candidate for both transferrin and non-transferrin-dependent uptake of iron. Ferrireductase activity is required prior to NTBI uptake and expression of cytochrome b also appears to increase with iron exposure (Ghio, 2009). Superoxide can participate in ferrireduction in lung epithelial cells trough ascrobat-mediated cyt b ferrireduction (Ghio et al., 2003). Anion-exchange protein is a novel system in the lung for transferring-independent iron transport, which superoxide dismutase significantly decrease iron import (Ghio et al., 2003; Turi et al., 2004). Export of intracellular metals can be mediated by either ferritin or transferrin release (Ghio, 2009). Release of ferritin from alveolar macrophages in BAL provides a mechanism to diminish iron stress (Wesselius et al., 1994). Metal transporter protein 1 (MTP 1) or ferroportin 1 which present mainly in the apical membrane of airway epithelium, release iron to either the airway or the alveoli rather than to the blood for systemic distribution (Yang et al., 2005), however, ferrireduction is required before transport which is done mainly by O2¯. Anion Exchange 2 (AE2) may also play a part in this release of metal (Yang et al., 2002).
Epithelial lining fluid contain several enzymatic and non-enzymatic antioxidant systems such as extracellular forms of superoxide dismutase (Su et al., 1997), glutathione peroxidase (Avissar et al., 1996), catalase (Cantin et al., 1999) as well as several metal-binding proteins (transferrin, ceruloplasmin and lactoferrin) that minimize involvement of transition metal ions in oxidant reaction (Cantin et al., 1999). Gland cell take up plasma and interstitial uric acid and secret it along with lactoferrin into the ELF (Pedan et al., 1993) where uric acid scavenges inhaled oxidants, reacts with singlet oxygen and also prevents oxidation of GSH and ascorbic acid by chelating transition metal ion (Davies et al., 1986).
In pathological condition, all these defenses overwhelmed and in presence of high oxygen concentration, iron result in production of powerful free radicals especially hydroxyl radical which can damage major macromolecules (lipid, DNA, protein) and lead to cell death.
Some studies demonstrate role of iron in lung oxidative injury and hyperoxia. Yang et al. (1999) proved that there was no increase in the levels of intracellular antioxidants, inflammatory cytokines and hemeoxygenase-1 in the hypotransferrinemic mouse lung exposed to hyperoxia (95% O2) compared with those in wild-type mice, however there were elevated expressions of ferritin and lactoferrin in the lung of hypotransferrinemic mice, especially in the alveolar macrophages. The degree of iron sequestration in transferrin was greatly increased in epithelial lining fluid (from 24 to 80%) but not in serum (remained 24%) of animals with respiratory failure, suggesting that the iron originated in the lung compartment (Hallman et al., 1994). Instillation of iron containing particles into airways resulted in an increased production of ferritin and lactoferrin and declined transferrin concentrations (Ghio et al., 1998). Hemeoxygenase is responsible for the metabolism of heme imported from the extracellulur space; thereby liberate carbon monoxide, bilirubin and ferrous iron. Bilirubin which has been shown to be a potent antioxidant, with albumin-bilirubin complexes being very efficient scavengers of free radicals (Voget et al., 1995). Intracellular free iron is sequestered by ferritin and the induction of ferritin by oxidants appear to be coupled to induction of HO-1, there are two isozymes of HO; HO-1 is the inducible enzyme that is induced by oxidants, whereas HO-2 is considered constitutive (Maines, 1988), recently a third isoenzymes, HO-3 which is similar to HO-2 has been described (McCoubrey et al., 1997). HO-1 expression in the rat lungs was shown to be increased following 24 to 72 h of a hyperoxic insult (Fogg et al., 1999).
Demnery and coworkers in a study exposed HO-2 null mutant mice to >95% O2 and compared with wild type controls. Similar basal levels were observed, except that the knockout had a twofold increase in total glutathione content and also were sensitized to hyperoxia-induced oxidative injury and mortality (Dennery et al., 1998). Wesselius et al. (1996) showed that there was dose dependent accumulation of iron and ferritin synthesis in alveolar macrophages exposed to iron-supplemented media. Exposure to hyperoxia (60 and 95% O2) also decreased iron uptake and to a great extent ferritin synthesis by AM in iron-supplemented media. These data suggest that iron uptake promote hyperoxic injury to AM and the hyperoxia impairs the capacity of AM to sequester iron in ferritin. Therefore iron was increased in lung where it catalyzes oxidative stress contributing to lung injury. Thus lung must have a delicate method for detoxifying iron to prevent its deleterious effects.
Role of iron chelator: Iron chelation has been recently used for conditions without iron overload such as neurodegenerative, infectious, reperfusion injury, cardioprotection and others suggesting the role of iron as an oxidative-induced injury (Hershko, 1994; Kontoghioghes et al., 2010). Desferrioxamine (DFO) has a very high affinity for Fe3+ and is very efficient in preventing its reduction to Fe2+ and the participation of iron in the Haber-Weiss reaction. DFO is a Hexidenate chelator. The maximal coordination of iron is six, so DFO binds to all sites and completely deactivate free iron (Hershko, 1992). DFO has been used in some diseases for prevention or reduction of oxidative stress induced injury (Deboer and Clark, 1992; Drossos et al., 1995; Menasche et al., 1987; Omar et al., 1989; Paraskevaidis et al., 2005; Prass et al., 2002), but few of these studies related to lung injury. For example combination of N-acetylcystein plus DFO decreased bronchoalveolarlavage (BAL) fluid protein such as carbonyl protein, inflammatory cells, oxidative damage variables and proinflammatory cytokines in rat after induction of acute lung injury by instillation of lipopolysaccaride. Ritter and coworkers concluded that this combination significantly attenuated lung oxidative damage, mitochondrial superoxide production and histopathological alterations in lung (Ritter et al., 2006). In another study, 42 male Wistar rats exposed to coal dust by intratracheal instillation. N-acetylcystein (20 mg/kg/day) alone or in combination with DFO (20 mg/kg/day) could decrease the inflammatory response and the oxidative stress parameters in these rats (Pinhoa et al., 2005). Aerosolized DFO could prevent severe pulmonary failure in sheeps exposed to burning cotton towlling (Lalonde et al., 1994). DFO due to very poor oral bioavailibilty and a short half-life must be administered by subcutaneous or intravenous infusion. New long acting oral agent such as deferasirox is a tridentate molecule containing 3 active binding sites for iron and has a binding ratio with 2:1 Fe3+ (Stumpf, 2007). This drug has better side effect profile in comparison with deferoxamin because of its small size and lipophilic structure that could chelate intracellular iron better than deferoxamine. So, it could be a promising agent for chelation of iron in situation other than thallasseamia (Vanorden and Hagemann, 2006). Currently authors of this paper have a clinical trial ongoing about usefulness of DFO in clinic that its primary results have been promising and is thought to be completed by end of 2010.
CONCLUSION
Oxygen therapy is one of the necessary parts of management of critically ill patients, especially with respiratory failure, but oxygen act as two edge sword in these situations and through oxidative stress process damage to major macromolecules that lead to cell death. Different pathways and molecules that involved in this process are the target of different drugs for prevention or treatment of harmful effect of oxygen. Despite favorable effects of these drugs, they are not completely successful in this condition. Regarding adequate evidence about role of iron in mediating of oxidative stress in hyperoxic conditions, protective effects of iron chelators should be considered reasonable. There are some evidences about positive effects of desferrioxamine alone or in combination when tested in animal models of lung injury or in human but they are not convincing. Further studies are necessary to clarify the importance of intervention with desferrioxamine or new long acting oral agent deferasirox and their risk/benefit in hyperoxia-induced lung injury.
REFERENCES
- Abdollahi, M., A. Dehpour and P. Kazemian, 2000. Alteration by cadmium of rat submandibular gland secretory function and the role of the L-arginine/nitric oxide pathway. Pharmacol. Res., 42: 591-597.
CrossRef - Abdollahi, M., A.R. Dehpour and F. Shafayee, 2000. L-Arginine/nitric oxide pathway and interaction with lead acetate on rat submandibulary gland function. Pharmacol. Toxicol., 87: 198-203.
CrossRef - Abdollahi, M., A. Ranjbar, S. Shadnia, S. Nikfar and A. Rezaie, 2004. Pesticides and oxidative stress: A review. Med. Sci. Monit., 10: RA141-RA147.
PubMedDirect Link - Ahmad, A., S. Ahmad, L.Y. Chang, J. Schaack and C.W. White, 2006. Endothelial Akt activation by hyperoxia: Role in cell survival. Free Radic. Biol. Med., 40: 1108-1118.
PubMed - Ansari, G., M. Mojtahedzadeh, F. Kajbaf, A. Najafi and M.R. Khajavi et al., 2008. How does blood glucose control with metformin influence intensive insulin protocols? Evidence for involvement of oxidative stress and inflammatory cytokines. Adv. Ther., 25: 681-702.
CrossRef - Aslam, M., R. Baveja, O.D. Liang, A. Fernandez-Gonzalez, C. Lee, S.A. Mitsialis and S. Kourembanas, 2009. Bone marrow stromal cells attenuate lung injury in a murine model of neonatal chronic lung disease. Am. J. Respir. Crit. Care Med., 11: 1122-1130.
PubMed - Auten, Jr. R.L., S.N. Mason, D.T. Tanaka, K. Welty-Wolf and M.H. Whorton, 2001. Anti-neutrophil chemokine preserves alveolar development in hyperoxia-exposed newborn rats. Am. J. Physiol. Lung Cell Mol. Physiol., 281: L336-L344.
Direct Link - Avissar, N., J.N. Finkelstein, S. Horowitz, J.C. Willey and E. Coy et al., 1996. Extracellular glutathione peroxidase in human lung epithelial lining fluid and in lung cells. Am. J. Physiol., 14: L173-L182.
PubMed - Beckett, W.S. and N.D. Wong, 1988. Effect of normobaric hyperoxia on airways of normal subjects. J. Applied Physiol., 64: 1683-1687.
PubMed - Birgens, H.S., L.O. Kristensen, N. Borregaard, H. Karle and H.E. Hansen, 1998. Lactoferrin-mediated transfer of iron to intracellular ferritin in human monocytes. Eur. J. Haematol., 41: 52-57.
PubMed - Bryan, C.L., R.E. Lewis, S.L. Owens, B. Emanuel and G. Jekinson, 1993. Allopurinol inhibition of neutrophilic alveolar response during hyperoxia. J. Applied Physiol., 75: 357-363.
PubMed - Cellier, M., G. Prive, A. Belouchi, T. Kwan, V. Rodrigues, W. Chia and P. Gros, 1995. Nramp defines a family of membrane proteins. Proc. Nat. Acad. Sci. USA., 92: 10089-10093.
CrossRef - Chow, C.W., M.T. Herrera Abreu, T. Suzuki and G.P. Downey, 2003. Oxidative stress and acute lung injury. Am. J. Respir. Cell Mol. Biol., 29: 427-431.
Direct Link - Cross, C.E., A. van der Vliet, C.A. O'Neill, S. Louie and B. Halliwell, 1994. Oxidants, antioxidants and respiratory tract lining fluids. Environ. Health Perspect., 102: 185-191.
CrossRefPubMedDirect Link - Das, D.K., D. Bandyopadyay, S. Hoory and H. Steinberg, 1988. Role of polymorphonuclear leukocytes in hyperoxic lung injury. Prevention of neutrophil influx into the lung endothelium during oxygen exposure by ibuprofen. Biomed. Biochim. Acta., 47: 1023-1036.
PubMed - Davies, K.J., A. Sevanian, S.F. Muakkassah-kelly and P. Hochstein, 1986. Uric acid-iron ion complexes. A new aspect of the antioxidant function of uric acid. Biochem. J., 235: 747-754.
PubMed - Davis, J.M., W.N. Rosenfeld, R.J. Sanders and A. Gonenne, 1993. Prophylactic effects of recombinant human superoxide dismutase in neonatal lung injury. J. Applied Physiol., 74: 2234-2241.
PubMed - Davis, W.B., S.I. Rennard, P.B. Bitterman and R.G. Crystal, 1983. Pulmonary oxygen toxicity. Early reversible changes in human alveolar structures induced by hyperoxia. NEJM., 309: 878-883.
PubMed - Deboer, D.A. and R.E. Clark, 1992. Iron chelation in myocardial preservation after ischemia-reperfusion injury: The importance of pretreatment and toxicity. Ann. Thorac. Surg., 53: 412-418.
PubMed - Drossos, G., A. Lazou, P. Panagopoulos and S. Westaby, 1995. Deferoxamine cardioplegia reduces superoxide radical production in human myocardium. Ann. Thorac. Surg., 59: 169-172.
CrossRef - Du, J., A.L.Z. Du, J.J Jiang, L.F. Tang and X.L. Wu, 2006. Effect of inhaled nitric oxide on surfactant protein A and mannose binding ability in the lung of neonatal rats with hyperoxia-induced lung injury. Zhongguo Dang Dai Er Ke Za Zhi., 8: 486-490.
PubMed - Emerit, J., C. Beaumont and F. Trivin, 2001. Iron metabolism, free radicals and oxidative injury. Biomed. Pharmacother., 55: 333-339.
Direct Link - Fogg, S., A. Agrawal, H. Nick and G. Visner, 1999. Iron regulates hyperoxia-dependent human Heme oxygenase 1 gene expression in pulmonary endothelial cell. Am. J. Respir. Cell Mol. Biol., 20: 794-804.
PubMed - George, C.L., G. Fantuzzi, S. Bursten, L. Leer and E. Abraham, 1999. Effects of lisofylline on hyperoxia-induced lung injury. Am. J. Physiol., 276: L776-L785.
Direct Link - Ghio, A., 2009. Disruption of iron homeostasis and lung disease. Biochem. Biophys. Acta, 1790: 731-739.
CrossRefPubMedDirect Link - Ghio, A., J. Carter, J. Richards, L. Brighton, J. Lay and R. Delvin, 1998. Disruption of normal iron homestasis after bronchial instillation of an iron-containing particle. Am. J. Physiol. Lung Cell Mol. Physiol., 274: L396-L403.
Direct Link - Ghio, A.J., P.J. Fracica, S.L. Young and C.A. Piantadosi, 1994. Synthetic surfactant scavenges oxidants and protects against hyperoxic lung injury. J. Applied Physiol., 77: 1217-1223.
PubMed - Hageman, J.R., S.E. Lee, J. Zemaitis, L.J. Smith and C.E. Hunt, 1989. Prostaglandin E1 infusion fails to prevent hyperoxic lung injury in adult rabbits. Crit. Care Med., 17: 339-344.
PubMed - Hallman, M., A. Sarnesto and K. Bry, 1994. Interaction of transferrin saturated with iron with lung surfactant in respiratory failure. J. Applied Physiol., 77: 757-766.
PubMed - Harrison, P.M and P. Arosio, 1996. The ferritins: Molecular properties, iron storage, function and cellular regulation. Biochem. Biophys. Acta, 1275: 161-203.
CrossRef - Hershko, C., 1994. Control of disease by selective iron depletion: A novel therapeutic strategy utilizing iron chelators. Baillieres Clin. Haematol., 7: 965-1000.
CrossRef - Jenkinson, S.G., R.J. Roberts, R.A. de Lemos, R.A. Lawrence and J.J. Coalson et al., 1991. Allopurinol-induced effects in premature baboons with respiratory distress syndrome. J. Applied Physiol., 70: 1160-1167.
PubMed - Lalonde, C., K. Ikegani and R. Demling, 1994. Aerosolized deferoxamine prevents lung and systemic injury caused by smoke inhalation. J. Applied Physiol., 77: 2057-2064.
PubMed - Langley, S.C. and F.J. Kelly, 1993. N-acetylcysteine ameliorates hyperoxic lung injury in the preterm guinea pig. Biochem. Pharmacol., 45: 841-846.
CrossRef - Lavaute, T., S. Smith, S. Cooperman, K. Iwai, E. Meyron-Holtz, S.K. Drake and G. Mitter, 2001. Targeted deletion of the gene encoding iron regulatory protein-2 causes misregulation of iron metabolism and neurodegenerative disease in mice. Nat Genet., 27: 209-214.
PubMed - Lee, J.H., D.K. Sung, S.H. Koo, B.K. Shin and Y.S. Hong et al., 2007. Erythropoietin attenuates hyperoxia-induced lung injury by down-modulating inflammation in neonatal rats. J. Korean Med. Sci., 22: 1042-1047.
CrossRef - Lee, P.J. and A.M. Choi, 2003. Pathways of cell signaling in hyperoxia. Free Radic. Biol. Med., 35: 341-350.
CrossRef - Li, W.B., L.W. Chang, Z.H. Rong, H.C. Liu and Q.S. Zhang et al., 2008. Retinoic acid diminished the expression of lung tissue matrix metalloproteinase-2 and matrix metalloproteinase-9 in hyperoxia-exposed premature rats through regulating mitogen-activated protein kinases. Zhonghua Er Ke Za Zhi., 46: 347-353.
PubMed - Lloyd, R.V., P.M. Hanna, P.M. and R.P. Mason, 1997. The origin of the hydroxyl radical in the Fenton reaction. Free Radic. Biol. Med., 22: 885-888.
CrossRef - Lyer, S. and B. Lonnerdal, 1993. Lactoferrin, lactoferrin receptors and iron metabolism. Eur. J. Clin. Nutr., 47: 232-241.
PubMed - Maines, M.D., 1988. Heme oxygenase: Function, multiplicity, regulatory mechanisms and clinical applications. FASEB J., 2: 2557-2568.
PubMed - Mateos, F., J. Brock and J.L. Perez-Arellano, 1998. Iron metabolism in the lower respiratory tract. Thorax, 53: 594-600.
CrossRefPubMedDirect Link - Matthew,.E., L. Kutcher and J. Dedman, 2003. Protection of lungs from hyperoxic injury: Gene expression analysis of cyclosporin A therapy. Physiol. Genomics., 14: 129-138.
PubMed - Menasche, P., C. Grousset, Y. Gauduel, C. Mouas and A. Piwnica, 1987. Prevention of hydroxyl radical formation: A critical concept for improving cardioplegia. Protective effect of deferoxamine. Circulation, 76: V180-V185.
PubMed - Mahmoodpoor, A., M. Mojtahedzadeh, A. Najafi, A. Ahmadi and A. Dehnadi-Moghadam et al., 2010. Examination of setarud (IMODTM) in the management of patients with severe sepsis. DARU-J. Pharm. Sci, 18: 23-28.
Direct Link - Najafi, A., M. Mojtahedzadeh, A. Mahmoodpoor, M. Aghamohammadi and A. Ahmadi et al., 2009. Effect of N-acetylcysteine on microalbuminuria in patients with acute respiratory distress syndrome. Arch. Med. Sci., 5: 408-414.
Direct Link - Mojtahedzadeh, M., M.R. Rouini, F. Kajbaf, A. Najafi and G. Ansari et al., 2008. Advantage of adjunct metformin and insulin therapy in the management of glycemia in critically ill patients. Evidence for nonoccurrence of lactic acidosis and needing to parenteral metformin. Arch. Med. Sci., 4: 174-178.
- Nelson, S.K. and J.M. McCord, 1998. Iron, oxygen radicals and disease. Adv. Mol. Cell. Biol., 25: 157-183.
CrossRef - Obara, H., S. Iwai and T. Takada, 1985. The effect of a high dose of vitamin E on the oxygen injured lung. Masui, 34: 286-291.
PubMed - Ozer, E.A., A. Kumral, E. Ozer, O. Yilmaz and N. Duman et al., 2005. Effects of erythropoietin on hyperoxic lung injury in neonatal rats. Pediatr. Res., 58: 38-41.
PubMed - Paine, R., S.E. Wilcoxen, S.B. Morris, C. Sartori, C.E. Baleeiro, M.A. Matthay and P.J. Christensen, 2003. Transgenic overexpression of granulocyte macrophage-colony stimulating factor in the lung prevents hyperoxic lung injury. Am. J. Pathol., 163: 2397-2406.
PubMed - Pantopoulos, K. and M.W. Hentz, 1998. Activation of iron regulatory protein-1 by oxidative stress in vitro. Proc. Nat. Acad. Sci. USA., 95: 10559-10563.
CrossRef - Paraskevaidis, I.A., E.K. Lliodromitis, D. Vlahakos, D. Tsiapras and A. Nikolaidis et al., 2005. Deferoxamine infusion during coronary artery bypass grafting ameliorates lipid peroxidation and protects the myocardium against reperfusion injury: imediate and long-term significance. Eur. Heart J., 26: 263-270.
CrossRefPubMed - Pedan, D.B., M. Swiersz, K. Ohkubo, B. Hahn, B. Emery and M.A. Kaliner, 1993. Nasal secretion of ozone scavenger uric acid. Am. Rev. Respir. Dis., 148: 455-461.
PubMed - Prass, K., K. Ruscher, M. Karsch, N. Isaev and D. Megow et al., 2002. Desferrioxamine induces delayed tolerance against cerebral ischemia in vivo and in vitro. J. Cereb. Blood Flow Metab., 22: 520-525.
CrossRef - Quinn, D.A., R.K. Moufarrej, A. Volokhov and C.A. Hales, 2002. Interactions of lung stretch, hyperoxia and MIP-2 production in ventilator-induced lung injury. J. Applied Physiol., 93: 517-525.
PubMed - Ritter, C., A.A. da Cunha, I.C. Echer, M. Andrades and A. Reinke et al., 2006. Effects of N-acetylcysteine plus deferoxamine in lipopolysaccharide-induced acute lung injury in the rat. Crit. Care Med., 34: 471-477.
PubMed - Rodriguez-Pierce, M., I.R. Sosenko, P. Whitney and L. Frank, 1994. Propylthiouracil treatment decreases the susceptibility to oxygen radical-induced lung damage in newborn rats exposed to prolonged hyperoxia. Pediatr. Res., 35: 530-535.
PubMed - Romashko, J., S. Horowitz, W.R. Franek, T. Palaia and E.J. Miller et al., 2003. MAPK pathways mediate hyperoxia-induced oncotic cell death in lung epithelial cells. Free Radic. Biol. Med., 35: 978-993.
CrossRef - Salari, P., M. Mojtahedzadeh and M. Abdollahi, 2005. Influence of serum epidermal growth factor on mechanical ventilation and survival in patients with acute respiratory distress syndrome. Therapy, 2: 393-398.
CrossRef - Salari, P., M. Mojtahedzadeh, A. Najafi, S. Sadraie, K. Bahaadini, M. Moharreri, N. Hadavandand M. Abdollahi, 2005. Comparison of the effect of aminophylline and low PEEP vs. high PEEP on EGF concentration in critically ill patients with ALI/ARDS. J. Clin. Pharmacy Therapeutics, 30: 139-144.
Direct Link - Sprong, C., Y.M.W. Janssen and P.J.A. Borm, 1991. The role of reactive oxygen species, cytokines and cytochrome P450 in pulmonary damage due to hyperoxia. Med. Hypotheses., 34: 296-299.
CrossRef - Soltan-Sharifi, M.S., M. Mojtahedzadeh, A. Najafi, M.R. Khajavi and M.R. Rouini et al., 2007. Improvement by N-acetylcysteine of acute respiratory distress syndrome through increasing intracellular glutathione and extracellular thiol molecules and antioxidant power: Evidence for underlying toxicological mechanisms. Hum. Exp., 26: 697-703.
CrossRefDirect Link - Su, W.Y., R. Folz, J.S. Chen, D.J. Capro and L.Y. Chang, 1997. Extacellular superoxide dismutase mRNA expressions in human lung by in situ hybridization. Am. J. Respir. Cell Mol. Biol., 16: 162-170.
PubMed - Thomson, A.M., J.T. Rogers and P.J. Leedman, 1999. Iron regulatory proteins,iron-responsive elements and ferritin mRNA translation. Int. J. Biochem. Cell Biol., 31: 1139-1152.
CrossRef - Town, G.I., G.J. Phillips, M. Landreau, J. Louden, S.T. Holgate and F.J. Kelly, 1993. Dexamethasone treatment fails to reduce oxygen-induced lung injury in the preterm guinea pig. Effects on pulmonary inflammation and antioxidant status. Biochem. Pharmacol., 46: 1565-1572.
CrossRef - Van der Vliet, A., C.A. O'Neill, C.E. Cross, J.M. Koostra, W.G. Volz, B. Halliwell and S. Louie, 1999. Determination of low-molecular-mass antioxidant concentrations in human respiratory tract lining fluids. Am. J. Physiol., 276: L289-L296.
PubMed - Van Klaveren, R.J., D. Dinsdale, J.L. Pype, M. Demedts and B. Nemery, 1997. N-acetylcysteine does not protect against type II cell injury after prolonged exposure to hyperoxia in rats. Am. J. Physiol., 273: L548-L555.
Direct Link - Van Snick, J.L., B. Markowetz and P.L. Masson, 1977. The ingestion and digestion of human lactoferrin by mouse peritoneal macrophages and the transfer of its iron into ferritin. J. Exp. Med., 146: 817-827.
PubMed - Vazin, A., M. Mojtahedzadeh, E. Salehifar, N. Rastkari, S. Khalaj, A. Rezaie and M. Abdollahi, 2005. Future drugs for treatment of acute respiratory distress syndrome. Int. J. Pharmacol., 1: 9-16.
CrossRef - Vazin, A., M. Mojtahedzadeh, A. Najafi, A. Khalilzadeh and M. Abdollahi, 2005. Relationship between duration, fatality rate and severity of disease and serum epidermal growth factor in human acute lung injury. Therapy, 2: 255-259.
CrossRef - Veness-Meehan, K.A., F.G. Bottone and A.D. Stiles, 2000. Effects of retinoic acid on airspace development and lung collagen in hyperoxia-exposed newborn rats. Pediatr. Res., 48: 434-444.
PubMed - Voget, B.A., J. Alam, A.J. Croatt, G.M. Veraallott and K.A. Nath, 1995. Acquired resistance to acute oxidative stress: Possible role of heme oxygenase and ferritin. Lab. Invest., 72: 474-483.
PubMed - Walther, F.J., R. David-Cu and S.L. Lopez, 1995. Antioxidant-surfactant liposomes mitigate hyperoxic lung injury in premature rabbits. Am. J. Physiol. Lung Cell Mol. Physiol., 269: L613-L617.
Direct Link - Wesselius, L., M.E. Nelson and B.S. Shikne, 1994. Increased release of ferritin and iron by iron-loaded alveolar macrophages in cigarette smokers. Am. J. Respir. Crit. Care Med., 150: 690-695.
PubMed - Wesselius, L., W. Wade, K. Baily, S. Vamos, A. O'brien-lander and Th.Weigmann, 1996. Iron uptake promotes hyperoxic injury to Alveolar macrophages. Am. J. Respir. Crit. Care Med., 159: 100-106.
Direct Link - Yang, F., J. Coalson, H. Bobb, J. Carter, J. Banu and A. Ghio, 1999. Resistance of hypotransferrinemic mice to hyperoxia-induced lung injury. Am. J. Physiol. Lung Cell Mol. Physiol., 277: L1214-L1223.
PubMedDirect Link - Yang, F., X. Wang, D.J. Haile, C. Piantadosi and A. Ghio, 2002. Iron increases expression of iron-export protein MTP1 in lung cells. Am. J. Physiol. Lung Cell Mol. Physiol., 283: L932-L939.
PubMedDirect Link - Zimova-Herknerova, M., J. Myslivek and P. Potmisil, 2008. Retinoic acid attenuates the mild hyperoxic lung injury in newborn mice. Physiol. Res., 57: 33-40.
PubMed - Kajbaf, F., M. Mojtahedzadeh and M. Abdollahi, 2007. Mechanisms underlying stress-induced hyperglycemia in critically ill patients. Therapy, 4: 97-106.
CrossRefDirect Link