
A. Aida Azlina
Department of Pharmacology,Faculty of Medicine, Universiti Kebangsaan Malaysia, Jalan Raja Muda Aziz, 50300 Kuala Lumpur, Malaysia
H.S. Farihah
Department of Anatomy, Faculty of Medicine, Universiti Kebangsaan Malaysia, Jalan Raja Muda Aziz, 50300 Kuala Lumpur, Malaysia
H.M.S. Qodriyah
Department of Pharmacology,Faculty of Medicine, Universiti Kebangsaan Malaysia, Jalan Raja Muda Aziz, 50300 Kuala Lumpur, Malaysia
M.F. Nur Azlina
Department of Pharmacology,Universiti Kebangsaan Malaysia, Jalan Raja Muda Aziz, 50300 Kuala Lumpur, Malaysia
International Journal of Pharmacology
Year: 2009 | Volume: 5 | Issue: 6 | Page No.: 362-369







ABSTRACT
The 11β-hydroxysteroid dehydrogenase type 1 (11β-HSD1) convert inactive circulating 11-keto steroids into active glucocorticoids, amplifying local glucocorticoid action. It is elevated in adipose tissue in obese humans and rodents, suggesting that adipose tissue glucocorticoid excess may be the causative factor for obesity. This study was conducted to evaluate the effects of Piper sarmentosum (PS) water extract and glycyrrhizic acid (GCA) on 11β-HSD1 bioactivity in ovariectomized induced obese rats. Fourty-two female Sprague-Dawley rats were randomly divided into six groups; four treatments (PS, GCA, CTRL and SHM) and two basal (B-CTRL and B-SHM). All groups underwent ovariectomy excluding SHM and B-SHM which underwent sham operation. Basal groups were sacrificed on the first day of treatment, while ovariectomized groups were given PS extract (0.125 g kg-1), GCA (0.120 g kg-1) and water (CTRL), respectively, while SHM received only water. Blood pressure was measured monthly while body weight weekly. After five months, rats were sacrificed and liver, heart and visceral adipose tissues were taken for analysis. Piper sarmentosum (PS) and GCA group showed a significant reduction in enzyme activity but no difference in body weight compared to CTRL group. Meanwhile only the blood pressure in GCA group was significantly higher after three months of treatment as compared to CTRL group but no difference after five months. In conclusion, both PS water extract and GCA have the ability to reduce 11β-HSD1 enzyme activity but only GCA cause an increased in blood pressure.
PDF Abstract XML References Citation
How to cite this article
A. Aida Azlina, H.S. Farihah, H.M.S. Qodriyah and M.F. Nur Azlina, 2009. Effects of Piper sarmentosum Water Extract on 11-β Hydroxysteroid Dehydrogenase Type 1 Bioactivity in Ovariectomy-Induced Obese Rats. International Journal of Pharmacology, 5: 362-369.
DOI: 10.3923/ijp.2009.362.369
URL: https://scialert.net/abstract/?doi=ijp.2009.362.369
DOI: 10.3923/ijp.2009.362.369
URL: https://scialert.net/abstract/?doi=ijp.2009.362.369
INTRODUCTION
Obesity can be defined as an accumulation of body fat (Gallagher et al., 1996) and it is associated with many diseases like hypertension, diabetes mellitus, coronary heart disease and others. It occurs as a result of either by hypertrophy of existing adipocytes or differentiation of preadipocytes through to adipocytes. A series of epidemiological studies have suggested that for a given body mass index, mortality is higher if adipose tissue is deposited centrally (visceral or omental obesity) as compared with generalized obesity (Björntop, 1991). It has been reported that visceral obesity is caused by excessive production of glucocorticoid exclusively within visceral adipose tissue (Masuzaki et al., 2001).
Glucocorticoids are regulated at the pre-receptor level by the microsomal enzyme 11β-hydroxysteroid dehydrogenase (11βHSD), which interconverts active glucocorticoids (cortisol in men; corticosterone in rodents) to inactive glucocorticoids (cortisone in men; 11-dehydrocorticosterone) (Stewart and Krozowski, 1999). Two different isoforms of 11βHSD enzymes have been identified; the 11β-HSD1 has a low affinity NADP(H)-dependent enzyme that activates the glucocorticoid, while the 11β-HSD2 is a high affinity NAD-dependent enzyme that inactivates the glucocorticoid.
The 11β-hydroxysteroid dehydrogenase type 1 (11β-HSD1) acts in vivo as a reductase, converting inactive to the active glucocorticoid (Katz et al., 1999). The link between obesity and the 11β-HSD1 enzyme had been investigated in many studies. It had been reported that 11β-HSD1 activity is increased in animal models of obesity (Livingstone et al., 2000). The 11β-HSD1 knock-out mice were found to be resistant to high fat diet-induced metabolic syndrome (Kotelevtsev et al., 1997; Morton et al., 2001) and mice with specifically over expressing 11β-HSD1 in adipose tissue showed all the typical features of the metabolic syndrome (Masuzaki et al., 2001, 2003). Moreover, both activity (Rask et al., 2001, 2002) and mRNA expression (Paulmyer-Lacroix et al., 2002) of 11β-HSD1 have been reported to be elevated in subcutaneous adipose tissues of obese human subjects. Therefore, the present study investigates the effects of water extracts of Piper sarmentosum and glycyrrhizic acid (GCA) on 11β-HSD1 bioactivity in ovariectomized induced obese rats.
For centuries, people have used herbs for the treatment of many ailments including chronic disease because of its trusted remedies that have been passed from generation to generation. One of the herbs that have great potential is Piper sarmentosum. Piper sarmentosum or known as ‘kaduk’ has been used widely in folk medicine especially in Southeast Asia region but very few studies have been documented scientifically.
In Malay traditional society, water decoction of the Piper sarmentosum leaves are being used for the treatment of cough, headache, waist pain and arthritis, while the water decoction of the roots are used in treatment of menstrual pain and to improve urination. Moreover, the pounded leaves are useful in the treatment of eczema. Most parts of the plant have potential benefits. The water extract of the whole plant had been shown to induce hypoglycemic effect in streptozocin-diabetic rats (Peungvicha et al., 1998). While the methanolic extract of the leaves was found to possess a marked neuromuscular blocking activity in the rat phrenic nerve-hemidiaphragm preparation. Furthermore, the chloroform and methanol extracts of the leaves have the ability to act as an antiplasmodial agent against Plasmodium falciparum and Plasmodium berghei parasites. In addition, the methanolic extracts of the leaves was found to have higher level of antioxidant activity compared to other traditional medicine plants. Therefore, this study was conducted to evaluate the effects of Piper sarmentosum (PS) water extract on 11β-HSD1 bioactivity, blood pressure and body weight in ovariectomy-induced obese rats.
MATERIALS AND METHODS
Preparation of Piper sarmentosum water extract: Fresh leaves (3 kg) of Piper sarmentosum were collected from the Ethnobotanic Garden, Forest Research Institute Malaysia (FRIM) after being identified and confirmed by a plant taxonomist from the Medicinal Plant Division (voucher specimen, FRI 45870), FRIM. All the extraction procedures were performed at the FRIM laboratory. Fresh leaves of the plants were cleaned with tap water and dried at room temperature before been chopped into small pieces. The leaves were then boiled with distilled water (90%, v/v) at 80°C for 3 h. The water extract was then concentrated and followed by freeze-drying to form powder. The powdered extract was stored at 4°C until further use.
Animal preparation: All procedures were carried out in accordance with the institutional guidelines for animal research of the Universiti Kebangsaan Malaysia (FAR/2006/AZLINA/19-APRIL/168-MARCH-2007). Fourty-two female Sprague-Dawley rats weighing 180-200 g were used in this study and placed in Animal House at Faculty of Medicine, Universiti Kebangsaan Malaysia. The rats were maintained in a temperature-controlled room (24°C) and illuminated for 12 h daily (lights on from 0700 to 1900). Two rats were kept in each cage and allowed free access to rat chow (Gold Coin, Klang, Selangor, Malaysia) and tap water ad libitum. The rats were randomly divided into six groups; four treatment groups (PS, GCA, CTRL and SHM) and two basal groups (B-CTRL and B-SHM). All groups underwent an experimental ovariectomy excluding SHM and B-SHM groups which underwent sham operation. Basal groups (B-SHM and B-CTRL) were sacrificed on the first day of treatment started. The ovariectomized groups were given Piper sarmentosum water extract (PS), glycyrrhizic acid (GCA) and water (CTRL), respectively, while the sham-operated (SHM) group received only water by oral feeding daily for 5 months.
Experimental protocol: The procedures were carried out in accordance with the institutional guidelines for animal surgery of the Universiti Kebangsaan Malaysia Animal Ethics Committee (UKMAEC). The rats were anaesthetized with intramuscular injection of ketamine (Troy Laboratories, Australia) and xylazine (Troy Laboratories, Australia) cocktail (1:1). The hairs on the both sides of the body were shaved off from the lowest rib to the hip region. For the rats which underwent a sham operation, only skin and muscles were cut but the ovaries were spared. Meanwhile for rats in the ovariectomized groups, bilateral ovariectomy was performed with an incision 1.5 cm inferior to the palpated rib cage. The ovaries and surrounding tissue were removed; the incision was closed by suturing the muscles and stapling the skin. Antibiotic cream was applied on the wound and the animal was placed on paper towels in a cage until awake. The treatment period was started after 2 weeks post-ovariectomy. Piper sarmentosum and GCA (Sigma, USA) were administered by oral gavage at a dose of 1.25 mg kg-1 (Peungvicha et al., 1998) and 1.20 mg kg-1 (Al-Wahaibi et al., 2007), respectively for five months. After five months of treatment, the rats were sacrificed and the liver, heart and visceral adipose tissues were taken for bioactivity analysis.
Bioactivity of 11β-HSD1 measurement: Enzyme activity was measured according to the method of Al-Wahaibi et al. (2007) with some modification. The liver, heart and visceral adipose tissues were homogenized in 2 mL Kreb-Ringer buffer. Two-hundred μmol of NADP (Sigma, USA) for 11β-HSD1 activity and 12 nM [3H]-corticosterone (Amersham, England) were added to tissue homogenates containing 0.5 mg protein. The total protein content was estimated calorimetrically (Bio-Rad, USA) on aliquots of each homogenates. Krebs-Ringer buffer containing 0.2% glucose (Sigma, USA) and 0.2% bovine serum albumin (Sigma, USA) were added to make up the total assay volume of 250 μL. The mixture was incubated in a shaking water bath at 37°C for 10 min (JE10 TECH, Korea). The reaction was terminated by the addition of 1 mL of ethyl acetate (Merck, Germany), steroids were then extracted. The organic layer was separated by centrifugation at 4°C, 3000 rpm for 10 min (IEC Centra EC4R, USA). The top layer was then transferred into test tube and evaporated using vacuum concentrated (Hetotrap VR10, USA). Steroid residues were dissolved in ethanol (Merck, Germany) containing 11-dehydrocorticosterone (Sigma, USA) and kept in 4°C overnight. The following day, it was separated by thin layer chromatography (Merck, Germany) in chloroform (Merck, Germany) and 95% ethanol (Merck, Germany) in the ratio of 92:8. The fractions corresponding to the steroid were located under ultraviolet lamp absorption 240 nm. It were scraped and transferred into scintillation vials and counted in scintillation fluid using fluid scintillation β-counter. Enzyme activity was calculated as the percentage conversion of [3H]-corticosterone to [3H]11-dehydrocorticosterone.
Body weight and blood pressure measurement: Body weight was taken every week by using the electronic weighing scale (Tanita Model 1144, Japan). Blood pressure was measured by using a physiograph machine. The rat was anaesthetized with diethyl ether (Merck, Germany). After which, the transducer was put at the tail. Then the cuff was inflated and the reading was recorded. This step was performed in triplicate and the average reading was taken as final reading.
Statistical analysis: All data were tested for normal distribution and presented as Mean±SD. Differences in the body weights and blood pressure were analyzed by a one-way Analysis of Variance (ANOVA) followed by Tukey test for multiple group comparisons. A value of p<0.05 was taken as significant. All statistical analysis was conducted using the SPSS version 11.5 software.
RESULTS
After ovariectomy, the 11βHSD1 activity in B-CTRL group were increased in all the three organs especially in liver 5% (p = 0.715) (Fig. 1) followed by visceral adipose tissues 2% (p = 0.055) (Fig. 3) and heart 1% (p = 1.000) (Fig. 2) as compared to B-SHM group.
However, after 5 months of treatment both PS and GCA treated group showed reduction in the 11βHSD1 activities as compared to CTRL group. For PS treated group, the most of the reduction in the activity occurred was in the liver 7% (p = 0.006) (Fig. 1) followed by the visceral adipose tissues 5% (p = 0.144) (Fig. 3) and the heart 5% (p = 0.161) (Fig. 2). Meanwhile in GCA treated group, the most of the reduction in the activity occurred was in the visceral adipose tissues 17% (p<0.001) (Fig. 3) followed by the liver 12% (p<0.001) (Fig. 1) and the heart 3% (p = 0.688) (Fig. 2).
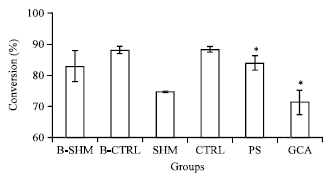 | |
| Fig. 1: | Effects of the oral administration of the water extract of Piper sarmentosum (PS) on the 11βHSD-1 enzyme activity in the liver of ovariectomized induced obese rats. B-SHM and B-CTRL were sacrificed on the day 0 of the treatment, SHM, CTRL, PS and GCA were sacrificed after 5 months of treatment. *Significant different compared with CTRL (*, p<0.05). N = 7 in each group. The data are the Mean±SD |
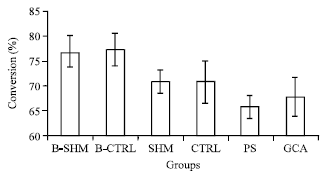 | |
| Fig. 2: | Effects of the oral administration of the water extract of Piper sarmentosum (PS) on the 11βHSD-1 enzyme activity in the heart of ovariectomized induced obese rats. B-SHM and B-CTRL were sacrificed on the day 0 of the treatment, SHM, CTRL, PS and GCA were sacrificed after 5 months of treatment. *Significant different compared with CTRL (*, p<0.05). N = 7 in each group. The data are the Mean±SD |
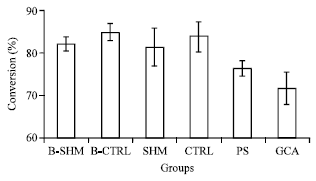 | |
| Fig. 3: | Effects of the oral administration of the water extract of Piper sarmentosum (PS) on the 11βHSD-1 enzyme activity in the visceral adipose tissue of ovariectomized induced obese rats. B-SHM and B-CTRL were sacrificed on the day 0 of the treatment; SHM, CTRL, PS and GCA were sacrificed after 5 months of treatment. *Significant different compared with CTRL (*, p<0.05). N = 7 in each group. The data are the Mean±SD |
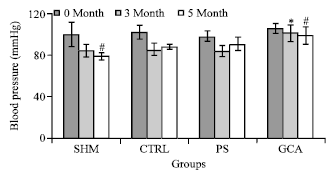 | |
| Fig. 4: | Comparisons of blood pressure between groups. Blood pressure was measured on 0 month, 3 month and 5 month of treatment. *Significant different compared with 3MONTH CTRL (*, p<0.05), #Significant different compared with 5MONTH CTRL (#, p<0.05 N = 7 in each group. The data are the Mean±SD |
The blood pressure measurement, at 0 month of treatment, showed no significant difference between all the four groups. However, only GCA treated group showed a significant higher blood pressure readings after 3 months (p = 0.001) and 5 months (p = 0.014) of treatment as compared to CTRL group (Fig. 4).
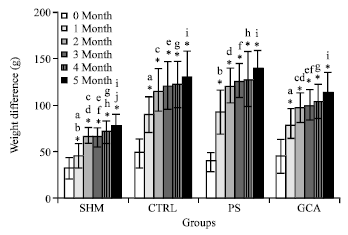 | |
| Fig. 5: | Weight difference between group over time. Same letters are statistically significant (p<0.05) between groups, *Significant difference between 0 MONTH (*, p<0.05). N = 7 in each group. The data are the Mean±SD |
Figure 5 shows that ovariectomy procedure caused obesity in the rats because the body weight increase was almost up to 20% as compared to the SHM group after 1 month of surgery. After 5 months of treatment, only GCA-treated group showed a significant (p<0.05) reduction in body weight difference as compared with CTRL group (Fig. 5).
DISCUSSION
This presence study was done to investigate the effect of ovariectomy on body weight and also on the activity of the 1β-hydroxysteroid dehydrogenase type 1 (11β-HSD1) enzyme in the liver, visceral adipose and heart tissues. It is known that ovariectomy procedure will cause increase in body weight which results in obesity (Chu et al., 1999). Present finding showed that the weight differences increased was almost up to 20% as compared to the SHM group after one month of surgery, which is comparable with Kano and Doi (2006) study. This observation is supported with the finding by Heymsfield et al. (1994) and Poehlman et al. (1995) which had suggested that menopause increased central adiposity. This is consistence with the view that it is associated with changes in fat distribution and estrogen deficiency that is believed to play a role in postmenopausal women fat distribution (Reubinoff et al., 1995). Moreover, estrogen has been found to influence eating behavior and cause weight gain in animals (Heymsfield et al., 1994).
It became evident that excessive glucocorticoid action plays a causal role in the pathogenesis of metabolic syndrome (Rosmond, 2005; Asensio et al., 2004). In particular, the enhanced local conversion of inactive glucocorticoids (cortisone, 11-dehydrocorticosterone) to active glucocorticoids (cortisol, corticosterone) by 11β-HSD1 in metabolically active tissues such as liver, adipose tissue and skeletal muscle has been implicated in these disorders (Tomlinson, 2005; Peterson et al., 2005; Wang, 2006). The pioneer study in 11β- HSD1 have purified the enzyme from the rat liver, followed by other studies that reported that it’s widespread of tissues distribution. In human, 11β-HSD1 was identified in samples from liver, testis, lung, foreskin fibroblast, ovary, colon and kidney with the highest level of expression in the liver and the lowest in the kidney (Tischer et al., 1991). Meanwhile in rats, 11β-HSD1 also has wide tissue distribution including liver, kidney, testis, lung, heart and colon (Agarwal et al., 1989; Krozowski et al., 1992; Brereton et al., 2001).
The effect of ovariectomy on 11β-HSD1 in the liver, visceral and heart tissues was investigated in this study. The enzyme activity was increased after ovariectomized in liver and visceral adipose but no difference in the heart. The increment in visceral adipose tissues was consistence with Rask et al. (2001) reported that 11β-HSD1 activity has increase in adipose tissues from obese human. In another study it was also reported that visceral adipose tissues has more higher 11β-HSD1 activity than subcutaneous adipose tissues and therefore preadipocytes from visceral adipose tissues are more prone to differentiated into adipocytes compared to subcutaneous adipose tissues (Berger et al., 2001). Similarly, the increment of hepatic 11β-HSD1 activity was comparable with Low et al. (1993) study where they reported that gonadectomy resulted in a significant increased in enzyme activity in the female rat liver. Explanation for the unaffected 11β-HSD1 activity in the heart was most probably because of the 11β-HSD1 activity in the heart was restricted to the interstitial cells of the endocardium and isolated cells surrounding cardiac vessels (Brereton et al., 2001).
There are minimum numbers of scientific studies which reports on the effects of Piper sarmentosum although it is widely been used as alternative medicine. The results of the present study demonstrated that the water extract of Piper sarmentosum had the ability to reduce the 11β-HSD1 enzyme activity in the liver, visceral adipose tissues and a minute effect in the heart tissues of ovariectomy-induced obese rats. Several studies have reported that Piper sarmentosum water extract had hypoglycemic effects in rats and in alloxan diabetic rabbits (Peungvicha et al., 1998). In our earlier study we found that Piper sarmentosum water extract reduced the blood glucose level and increase the adiponectin plasma level in ovariectomy-induced obese rats (Aida Azlina et al., 2009). Treatment of obese mouse strains with selective 11β-HSD1 inhibitors resulted in significant reduced visceral fat deposits, accompanied with improvement of glucose tolerance and insulin sensitivity (Alberts et al., 2002, 2003). As hypothesized, the activity of the 11β-HSD1 enzyme in PS and GCA treated groups were significantly (p<0.05) reduced in all the three organs as compared to CTRL group. Therefore, there is a possibility that Piper sarmentosum water extract has the ability to reduce the activity of the 11βHSD1 enzyme.
After five months of treatment, significant (p<0.05) reduction in body weight only occurred in GCA treated group. For PS treated group, there was no dissimilarity as compared to CTRL group. This result was not shown to be correlated with the bioactivity of 11βHSD1. Explanation for this probably because of the reduction of 11βHSD1 enzyme activities is site specific which is occurs more at the visceral adipose tissues compared with subcutaneous fat, when in fact, body weight measurement were included the subcutaneous adipose tissues as well. This finding were supported with other studies (Hauner et al., 1989; Fried et al., 1993) which have reported that the expression of 11βHSD1 enzyme was highest occurred in the visceral compared with the subcutaneous adipose tissues.
Glycyrrhetinic acid, the main metabolite of glycyrrhizic acid (GCA) is potent but not selective inhibitors for 11β-HSDs isoforms (Monder et al., 1989). It has been reported that it can reduce the thickness of subcutaneous thigh fat through tropical application (Armanini et al., 2005). However, prolonged ingestion of glycyrrhizic acid can cause hypertension because it may develop pseudo aldosterone-like effects (Heikens et al., 1995).
Present finding on blood pressure agrees with Heikens et al. (1995) finding that glycyrrhizic acid may induce hypertension by inhibition of cortisol metabolism which results in the pseudo-aldosterone- like effects. On the zero month of treatment, there was no significant difference between the sham-operated and ovariectomized rats. However, the blood pressure of the GCA treated group was significantly higher after three months of treatment compared to other group which confirms the hypothesis that glycyrrhizic acid increases the blood pressure. Krozowski and Funder (1983), Edwards et al. (1988) and Funder et al. (1988) proposed that conversion of cortisol (corticosterone for rodents) to cortisone (11-dehydrocorticosterone) represent the physiological mechanism conferring specificity for aldosterone upon the mineralocorticoid receptors. Since, cortisol normally circulates at levels 100-1000 time compared to aldosterone, this leads to signs of mineralocorticoid excess, even though aldosterone secretion is suppressed. Thus, in prolonged glycyrrhizic acid ingestion, it permits cortisol to occupy the mineralocorticoid receptors which leads to hypertension.
CONCLUSION
As a conclusion, both Piper sarmentosum water extract and glycyrrhizic acid reduces the activity of the 11β-hydroxysteroid dehydrogenase type 1 enzyme, which suggest that Piper sarmentosum water extract may have the ability to inhibit the 11βHSD1 enzyme activity. However, only glycyrrhizic acid can reduce the body weight as well as cause an increase in the blood pressure in overiectomized induced obese rats.
ACKNOWLEDGMENTS
The researchers would like to thank Pn. Azizah Osman, Pn Sinar Suriya Muhamad, Pn. Mazliadiyana Mazlan, Pn Norhayati Md Yasin and all the technical staff of the department of Pharmacology, UKM for their help. We would also like to thank the Ministry of Science, Technology and Innovation for the grant IRPA 06-02-02-10026EAR; and to Universiti Kebangsaan Malaysia for the research grant FF-164-2006. We are also grateful to the Forest Research Institute Malaysia (FRIM) for the supply of Piper sarmentosum extract.
REFERENCES
- Agarwal, A.K., C. Monder, B. Eckstein and P.C. White, 1989. Cloning and expression of rat cDNA encoding corticosteroid 11beta-hydroxysteroid dehydrogenase. J. Biol. Chem., 264: 18939-18943.
Direct Link - Aida Azlina, A., S. Farihah, M.S. Qodriyah and M.F. Nur Azlina, 2009. Effects of Piper sarmentosum (Kaduk) water extract on adiponectin and blood glucose levels in ovariectomy-induced obese rats. Res. J. Med. Plants (In Press).
Direct Link - Alberts, P., L. Engblom, N. Edling, M. Forsgren and G. Klingstrom et al., 2002. Selective inhibition of 11beta-hydroxysteroid dehydrogenase type 1 decreases blood glucose concentrations in hyperglycemic mice. Diabetologia, 45: 1528-1532.
PubMedDirect Link - Alberts, P., L. Engblom, N. Edling, M. Forsgren and G. Klingstrom et al., 2003. Selective inhibition of 11beta-hydroxysteroid dehydrogenase type 1 improves hepatic insulin sensitivity in hyperglycemic mice. Endocrinology, 144: 4755-4762.
PubMedDirect Link - Armanini, D., D. Nacamulli, F. Francini-Pesenti, G. Battagin, E. Ragazzi and C. Fiore, 2005. Glycyrrhetinic acid, the active principle of licorice, can reduce the thickness of subcutaneous thigh fat through topical application. Steroid, 70: 538-542.
CrossRef - Asensio, C., P. Muzzin and F. Rohner-Jeanrenaud, 2004. Role of glucocorticoids in the physiopathology of excessive fat deposition and insulin resistance. Int. J. Obes. Relat. Metab. Disord., 28: 45-52.
PubMedDirect Link - Bjorntop, P., 1991. Adipose tissue distribution and function. Int. J. Obes., 15: 67-81.
PubMedDirect Link - Brereton, P.S., R.R.V. Driel, F.R. Suhaimi, K. Koyama, R. Dilley and Z. Krozowski, 2001. Light and electron microscopy localization of the 11beta-hydroxysteroid dehydrogenase type 1 enzyme in the rat. Endocrinology, 142: 1644-1651.
PubMedDirect Link - Chu, S.C., Y.C. Chou, J.Y. Liu, C.H. Chen, J.C. Shyu and F.P. Chou, 1999. Fluctuation of serum leptin level in rats after ovariectomy and the influence of estrogen supplement. Life Sci., 64: 2299-2306.
CrossRefDirect Link - Edwards, C.R., P.M. Stewart, D. Burt, L. Brett and M.A. McIntyre(et al)., 1988. Localization of 11 beta-hydroxysteroid dehydrogenase-tissue specific protector of the mineralocorticoid receptor. Lancet, 2: 986-989.
PubMedDirect Link - Fried, S.K., C.D. Russell, N.L. Grauso and R.E. Brolin, 1993. Lipoprotein lipase regulation by insulin and glucocorticoid in subcutaneous and omental adipose tissue of obese women and men. J. Clin. Invest., 92: 2191-2198.
PubMedDirect Link - Funder, J.W., P.T. Pearce, R. Smith and A.I. Smith, 1988. Mineralocorticoid action: Target tissue specificity is enzyme, but not receptor, mediated. Science, 242: 583-585.
CrossRefDirect Link - Gallagher, D., M. Visser and D. Sepulveda, 1996. How useful is body mass index for comparison of body fatness across age, sex and ethnic group? Am. J. Epidemiol., 143: 228-229.
Direct Link - Hauner, H., G. Entenmann, M. Wabitsch, D. Gaillard, G. Ailhaud, R. Negrel and E.F. Pfeiffer, 1989. Promoting effect of glucocorticoids on the differentiation of human adipocyte precursor cells cultured in a chemically defined medium. J. Clin. Invest., 84: 1663-1670.
CrossRefDirect Link - Heikens, J., E. Fliers, E. Endert, M. Ackerman and G. van Montfrans, 1995. Liquorice-induced hypertension: A new understanding of an old disease: Case report and brief review. Neth. J. Med., 47: 230-234.
PubMedDirect Link - Kano, S. and M. Doi, 2006. NO-1886 (ibrolipim), a lipoprotein lipase-promoting agent, accelerates the expression of UCP3 messenger RNA and ameliorates obesity in ovariectomized rats. Metabol. Clin. Exp., 55: 151-158.
Direct Link - Katz, J.R., V. Mohamed-Ali, P.J. Wood, J.S. Yudkin and S.W. Coppack, 1999. An in vivo study of the cortisol-cortisone shuttle in subcutaneous abdominal adipose tissue. Clin. Endocrinol. (Oxf.), 50: 63-68.
Direct Link - Kotelevtsev, Y., M.C. Holmes, A. Burchell, P.M. Houston and D. Schmoll et al., 1997. 11beta-hydroxysteroid dehydrogenase type 1 knockout mice show attenuated glucocorticoid inducible responses and resist hyperglycemia on obesity or stress. Proc. Natl. Acad. Sci. USA., 94: 14924-14929.
PubMedDirect Link - Krozowski, Z.S. and J.W. Funder, 1983. Renal mineralocorticoid receptors and hippocampal corticosterone binding species have identical intrinsic steroid specificity. Proc. Natl. Acad. Sci. USA., 80: 6056-6060.
PubMedDirect Link - Krozowski, Z.S., V. Obeyesekere, R. Smith and W. Mercer, 1992. Tissue-specific expression of an 11beta-hydroxysteroid dehydrogenase with a truncated N-terminal domain. A potential mechanism for differential intracellular localization within mineralcorticoid target cells. J. Biol. Chem., 267: 2569-2574.
- Livingstone, D.E.W., G.C. Jones, K. Smith, R. Andrew, C.J. Kenyon and B.R. Walker, 2000. Understanding the role of glucocorticoids in obesity: Tissue-specific alterations of corticosterone metabolism in obese Zucker rats. Endocrinology, 141: 560-563.
PubMedDirect Link - Low, S.C., S.N. Assard, V. Rajan, K.E. Chapman, C.R. Edwards and J.R. Seckl, 1993. Regulation of 11beta-hydroxysteroid dehydrogenase by sex steroids in vivo: Further evidence for the existence of a second dehydrogenase in rat kidney. J. Endocrinol., 139: 27-35.
CrossRefDirect Link - Masuzaki, H., J. Peterson, H. Shinyama, N.M. Morton, J.J. Mullins, J.R. Seckl and F.S. Flier, 2001. A transgenic model of visceral obesity and the metabolic syndrome. Science, 294: 2166-2170.
CrossRefPubMedDirect Link - Monder, C., P.M. Stewart, V. Lakshmi, R. Valentino, D. Burt and C.R. Edwards, 1989. Licorice inhibits corticosteroid 11beta-HSD of rat kidney and liver in vivo and in vitro studies. Endocrinology, 125: 1046-1053.
Direct Link - Morton, N.M., M.C. Holmes, C. Fievet, B. Staels, A. Tailleux, J.J. Mullins and J.R. Seckl, 2001. Improved lipid and lipoprotein profile, hepatic insulin sensitivity and glucose tolerance in 11beta-hydroxysteroid dehydrogenase type 1 null mice. J. Biol. Chem., 276: 41293-41300.
PubMedDirect Link - Paulmyer-Lacroix, O., S. Boullu, C. Oliver, M.C. Alessi and M. Grino, 2002. Expression of the messenger RNA coding for 11beta-HSD type 1 in adipose tissue from obese patients: An in situ hybridization study. J. Clin. Endocrinol. Metab., 87: 2701-2705.
PubMedDirect Link - Peterson, J.M., J.R. Seckl and J.J. Mullins, 2005. Genetic manipulation of 11beta-hydroxysteroid dehydrogenases in mice. Am. J. Physiol. Regul. Integr. Comp. Physiol., 289: 642-652.
Direct Link - Poehlman, E.T., M.J. Toth and A.W. Gardner, 1995. Changes in energy balance and body composition at menopause: a controlled longitudinal study. Ann. Int. Med., 123: 673-675.
Direct Link - Rask, E., T. Olsson, S. Soderberg, R. Andrew, D.E.W. Livingstone, O. Johnson and B.R. Walker, 2001. Tissue-specific disregulation of cortisol metabolism in human obesity. J. Clin. Endocrinol. Metab., 86: 1418-1421.
Direct Link - Rask, E., B.R. Walker, S. Soderberg, D.E.W. Livingstone and M. Eliasson(et al)., 2002. Tissue specific changes in peripheral cortisol metabolism in obese women: Increased adipose 11beta-HSD type 1 activity. J. Clin. Endocrinol. Metab., 87: 3330-3336.
PubMedDirect Link - Rosmond, R., 2005. Role of stress in the pathogenesis of the metabolic syndrome. Psychoneuroendocrinology, 30: 1-10.
CrossRef - Reubinoff, B.E., J. Wurtman and L. Rojanski, 1995. Effects of hormone replacement therapy on weight, body fat composition, fat distribution and food intake in early postmenopausal women: A prospective study. Fertil. Steril., 64: 963-968.
Direct Link - Tischer, E., R. Mitchell, T. Hartman, M. Silva, D. Gospodarowicz, J.C. Fiddes and J.A. Abraham, 1991. The human gene for vascular endothelial growth factor. Multiple protein forms are encoded through alternative exon splicing. J. Biol. Chem., 266: 11947-11954.
PubMedDirect Link - Tomlinson, J.W., 2005. 11Beta-hydroxysteroid dehydrogenase type 1 in human disease: A novel therapeutic target. Minerva. Endocrinol., 30: 37-46.
PubMedDirect Link - Wang, M., 2006. Inhibitors of 11beta-hydroxysteroid dehydrogenase type 1 for the treatment of metabolic syndrome. Curr. Opin. Invest. Drugs, 7: 319-323.
Direct Link