
U.S. Rao Chakradhara
Department of Pharmacology, K.S. Hegde Medical Academy, Mangalore 575 018, India
K.S. Karanth
Department of Pharmacology, K.S. Hegde Medical Academy, Mangalore 575 018, India
International Journal of Pharmacology
Year: 2009 | Volume: 5 | Issue: 5 | Page No.: 327-332







ABSTRACT
Ethanol withdrawal increases sensitivity to painful stimuli. N-methyl-D-aspartate glutamate receptors may have a role in alcohol dependence and the development of withdrawal signs and symptoms. This study examined the effect of oral administration of dextromethorphan on the hyperalgesia induced by ethanol withdrawal, using hot plate assay and chemical induced writhing in rats. Wistar albino rats (250-300 g) were divided into three groups of six animals each. All the groups received 7.5% v/v alcohol and food ad libitum, for 10 days and were given saline (10 mL kg-1) or dextromethorphan dissolved in saline (32 and 64 mg kg-1) orally once daily for 10 days. Ethanol was withdrawn on day 10. The reaction times on the hot plate were measured on Day 0, at 6, 12 h after ethanol withdrawal. The ethanol withdrawal signs were rated immediately before testing. Chemical assay using acetic acid (1% v/v) was done on day 0 and at 12 h after ethanol withdrawal. Chronic exposure to ethanol produced antinociception while withdrawal produced hyperalgesia. Repeated administration of dextromethorphan prevented ethanol withdrawal induced hyperalgesia and significantly reduced total ethanol withdrawal scores at both the doses tested. This study demonstrated that dextromethorphan can attenuate hyperalgesia during ethanol withdrawal and suggests the role of N-methyl-D-aspartate receptors in ethanol withdrawal induced hyperalgesia.
PDF Abstract XML References Citation
How to cite this article
U.S. Rao Chakradhara and K.S. Karanth, 2009. Dextromethorphan Attenuates Ethanol Withdrawal Induced Hyperalgesia in Rats. International Journal of Pharmacology, 5: 327-332.
DOI: 10.3923/ijp.2009.327.332
URL: https://scialert.net/abstract/?doi=ijp.2009.327.332
DOI: 10.3923/ijp.2009.327.332
URL: https://scialert.net/abstract/?doi=ijp.2009.327.332
INTRODUCTION
Ethanol is one of the most widely abused substances and is known to cause a withdrawal syndrome associated with increased sensitivity to pain, hyperalgesia (hypersensitivity to noxious stimuli), anxiety, agitation and decreased seizure threshold (Lieber, 1995; Dina et al., 2006). Central sensitization in spinal nociceptive pathways has been proposed to cause hyperalgesia (Yaksh et al., 1999; Mao and Mayer, 2001). Very little is known about Ethanol Withdrawal (EW) induced hyperalgesia and is known to be reversed by either ethanol or drugs like COX2 inhibitors, theophylline and nitrendipine (Pohorecky and Shah, 1987; Gatch, 1999, 2002; Gatch and Lal, 1999; Dhir et al., 2005). This suggests the involvement of cAMP, calcium channels in eliciting hyperalgesia due to EW.
N-methyl-D-aspartate (NMDA) glutamate receptors are known to have a role in alcohol dependence and withdrawal (Roberto et al., 2006; Hendricson et al., 2007; Narita et al., 2007a). Hyper-responsiveness seen during EW followed by chronic administration or brief exposure to ethanol is dependent on NMDA receptors (Li and Kendig, 2003). Thus EW appears to be triggered by NMDA receptors activation during ethanol exposure especially in the motor pathway (Li and Kendig, 2003). It is proposed that a synergistic effect of increased NMDA receptor function and Voltage-Sensitive Calcium Channel (VSCC) function may also contribute to EW induced hyper excitability (Davis and Wu, 2001; Narita et al., 2007b; Hoffman and Tabakoff, 1996). Thus, NMDA receptor antagonists may be used to block the EW syndrome and are also known to prevent development of tolerance to antinociceptive effects of morphine (Cha et al., 2003). Dextromethorphan (DM, a commonly available over-the-counter anti-tussive preparation) and its metabolite Dextrorphan are non-selective NMDA receptor antagonists. Both DM and its metabolite have been found to be effective in reducing EW induced locomotor hyperactivity and audiogenic seizures in rats (Erden et al., 1999). However, no information available about the role of NMDA receptors on EW induced hyperalgesia and allodynia. We hypothesize that EW induced hyperalgesia may be due to the increased NMDA receptor activation during ethanol exposure which is revealed when the depressant effects of ethanol are removed. In an attempt to provide the behavioral evidence for the hypothesis we studied the effect of oral administration of DM on EW induced hyperalgesia in rats.
MATERIALS AND METHODS
This study was conducted at the Department of Pharmacology, K.S. Hegde Medical Academy, Mangalore, India during the period February 2006 to November 2006. Hotplate test (Eddy and Leimback, 1953) and acetic acid induced writhing (Seigmund and Lu, 1957) were employed for nociception testing in rats. The animals were confirmed for EW according to the scores defined earlier (Gatch, 2002).
Animals: Two experiments were performed for hot plate or chemical nociception assays using Wistar albino rats (250-300 g). In each experiment three groups of rats with 6 animals in each were employed. Animal in group 1 were kept as controls and those of groups 2 and 3 served as DM treatment groups with low dose (32 mg kg-1) or high dose (64 mg kg-1), respectively. All the rats were housed and maintained on 12:12 h light and dark cycle (Light on at 7:00 am). All housing procedures were in accord with the guidelines of the Institute of Laboratory Animal Resources, National Research Council (USA). The study was approved by Institutional Animal Ethics Committee of K.S. Hegde Medical Academy (Mangalore, India).
Ethanol administration and withdrawal: Animals in all the three groups received 7.5% ethanol solution in water and food orally ad libitum and were administered either saline 10 mL kg-1 (group1) or DM (dissolved in saline) at the dose of 32 mg kg-1 (group 2) or 64 mg kg-1 (group 3) orally, once daily between 8:00-8:45 am for 10 days. On day 10, the last doses of the drug or saline were administered just before the standardization of EW and all the rats had free access to ethanol until 8:00 am. The rats were gavaged with a dose of ethanol of 3 mL kg-1 to standardize the starting time of EW. During the withdrawal period rats were given free access to food and water ad libitum. Daily ethanol consumption was also measured during the study period. Immediately before performing the nociception assay animals were rated and confirmed for EW status based on the scoring scale adapted for rats as described earlier (Gatch, 2002).
Nociception assays: Two models, viz. eddy’s hot plate test (thermal method) (Eddy and Leimback, 1953) and acetic acid induced writhing response (chemical method) (Seigmund and Lu, 1957) were employed to study the nociceptive behavior of animals.
Hotplate tests (thermal method): The test was performed using eddy’s hot plate maintained at a temperature of 55±1°C. The basal reaction time of all animals towards thermal heat was recorded on the first day of the study before the administration of DM or saline and at 6, 12 h after EW. During each testing session, three readings were taken at an interval of 15 min for each animal. The time taken by the rat for paw licking or jumping was noted. A cut off period of 15 sec was set to avoid damage to the paws.
Acetic acid induced writhing test (chemical method): Acetic acid (1% v/v) was administered intraperitoneally to all the groups and nociceptive behavior was recorded by counting the number of times the rat writhed after the injection of acetic acid for a period of 15 min. A writhing is indicated by abdominal constriction and full extension of hind limb. Chemical induced nociception assay was performed on the first day to check the typical response behaviour by animals and only those animals showing typical writhing response were included in the study. The test was performed at 12 h after the EW.
Statistical analysis: GraphPad Instat® version 3.06 (San Diego, USA) was used for statistical analysis. The average of three readings obtained by the hot plate method and by chemical nociception was compared among the groups by one way ANOVA followed by Tukey-Kramer multiple comparisons post hoc test. The data were expressed as Mean±SD; p<0.05 was considered statistically significant.
RESULTS
Nociception assay: The baseline reaction times on eddy’s hot plate were comparable across test groups. No significant difference in basal reaction times was found between the control (4.7±0.4) and the DM treatment groups (group 2, 4.7±0.5; group 3, 4.7±0.8) (Fig. 1). The exposure to ethanol for 10 days followed by withdrawal produced hyperalgesia, which was reflected by lowered reaction times on eddy’s hot plate (2.8±0.3 at 6 h after EW and 2.2±0.3 at 12 h after EW), increase in the number of writhings (33.3±3.1) and decrease in the onset of time for writhing (1.3±0.3) in the control group.
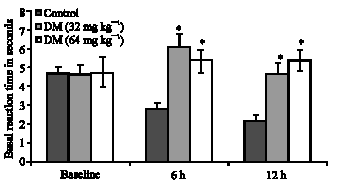 | |
| Fig. 1: | Effect of dextromethophan on basal reaction time in eddy’s hotplate method (thermal) before starting the treatment and at 6 and 12 h after ethanol withdrawal. Values are represented as Mean±SD. The temperature used in eddy’s hot plate was 55±1°C. Three different readings were taken for each animal in a group (n = 6 in each group) at an interval of 15 min and the average was taken for each testing period. The mean values for each group represents the sum of the average of the readings for all animals in that group. *The values are significant compared to that of control group (p<0.001) |
| Table 1: | Effect of Dextromethorphan on chemical induced nociception 12 h after ethanol withdrawal |
 | |
| Saline 10 mL kg-1 was given for control rats. Values are represented as Mean±SD; 1% v/v acetic acid was used for inducing pain in animals. *Significant difference compared to the control group (p<0.001). A significant difference in the time of onset of writhing was observed between DM 32 mg kg-1 and DM 64 mg kg-1 groups (p<0.001). But no significant difference in No. of writhings was observed between these two groups | |
Repeated administration of DM in two doses (32 and 64 mg kg-1) prevented the EW induced hyperalgesia at 6 and 12 h after EW (Fig. 1). The time taken by the control rats to lick the paw or jumping response in eddy’s hot plate was significantly lower compared to DM treatment groups at 6 and 12 h after EW (Fig. 1). But there was no significant difference in basal reaction times was observed between DM 32 mg kg-1 treated group (6.1±0.7 at 6 h after EW and 4.6±0.6 at 12 h after EW) and DM 64 mg kg-1 treated group (5.4±0.6 at 6 h after EW and 5.4±0.5 at 12 h after EW). Similarly, in chemical nociception assay DM significantly lowered the number of writhings and extended the time of onset of writhing at both the doses tested (32 and 64 mg kg-1). A significant difference in the time of onset of writhing was observed between DM 32 mg kg-1 (5.2±0.8) and DM 64 mg kg-1 (7.8±1.0) groups (p<0.001), But no significant difference in number of writhings was observed between these two groups. The details of number of writhings and time of onset of writhing in each group tested are given in Table 1.
Ethanol consumption and ethanol withdrawal signs: The DM decreased ethanol consumption during the study period (Fig. 2). Exposure to ethanol for 10 days followed by withdrawal produced mild to moderate withdrawal signs, which was confirmed by the scoring for the withdrawal signs, with highest scores in control group with a rating scale 0-25. The consumption of ethanol in DM-treated animals was not reduced to such extent to witness no EW signs after cessation of ethanol supply on day 10. The EW scores in the control group were 5.63±0.3 at 6 h after EW and 5.3±0.4 at 12 h after EW (maximum possible score is 25). Signs of EW were observed including increased locomotor activity, stereotype behavior, abnormal posture and gait, urination, defecation on handling, mild tremors and tail stiffness. No seizures or mortality was observed in any group.
| Table 2: | Effect of DM on ethanol withdrawal score at 6 and 12 h after ethanol withdrawal |
 | |
| Saline 10 mL kg-1 was given for control rats. Values are represented as Mean±SD. *p<0.01 compared to that of control group. DM reduced the EW scores at 6 h after EW at the both the doses tested (p<0.001). DM also reduced EW scores in a dose dependant manner with significant difference between the dose groups i.e., 32 and 64 mg kg-1 (p<0.05). No significant reduction in EW scores by DM was observed at 12 h after EW. Total number of animals in each group was 12, which includes 6 animals from chemical nociception assay and 6 animals from hotplate assay | |
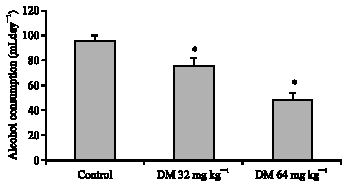 | |
| Fig. 2: | Effect of dextromethorphan on ethanol consumption. Ethanol consumption for each group was measured in milliliters during the total period of 10 days. This was converted to consumption of ethanol in milliliters per day for each group. Total number of animals in each group was 12, which includes 6 animals from chemical nociception assay and 6 animals from hotplate assay. *The values are significant compared to that of control (p<0.001). DM dose dependently decreased ethanol consumption and there is a significant reduction in ethanol consumption observed in DM 64 mg kg-1 group compared to DM 32 mg kg-1 (p<0.001) |
Higher EW scores were found at 12 h after the EW as compared with the scores at 6 h after EW (Table 2). DM decreased the EW scores only at 6 h after EW, but not at 12 h after EW (Table 2).
DISCUSSION
The major finding of the study is the attenuation of EW induced hyperalgesia by DM. This was evident as a significant rise in basal reaction time in the hot plate method and delayed onset of writhing and lower number of writhings in DM treated groups as compared with the control group. The mechanisms underlying the EW syndrome are not clear. Studies have shown that excitatory aminoacid neurotransmitters are involved in eliciting the signs and symptoms of EW (Rossetti and Carboni, 1995). A earlier study has shown that DM attenuates the EW syndrome in rats, suggesting that it can be a useful drug in the treatment of EW (Erden et al., 1999). On withdrawal of ethanol, a state of hyperalgesia was reported in rats (Gatch and Lal, 1999; Gatch, 1999). Few studies have reported the role of GABA, adenosine and calcium channels in EW induced hyperalgesia (Gatch, 1999, 2002; Gatch and Selvig, 2002). We confirmed the appearance of EW in rats and scored it accordingly. The duration of Ethanol administration was only for 10 days. However, control group and treatment groups showed marked EW signs, confirming a withdrawal state. The DM was able to lower EW scores at 6 h after EW. This may be because of the blockade of NMDA receptor activation or up regulation during exposure to ethanol and after the EW by DM. But no difference in EW scores has been found at 12 h after EW which may be due to the unavailability of the DM to block NMDA receptors and might have been due to increased sensitization of the NMDA receptors at 12 h after EW after continuous blockade by DM. The DM attenuated hyperalgesia at both 6 and 12 h after EW, but was unable to decrease EW scores at 12 h after EW suggesting different mechanisms involved in preventing EW syndrome and EW induced hyperalgesia. Prevention of ethanol withdrawal syndrome might have been due to the blockade of NMDA receptors, where as the attenuation of hyparlgesia might be due to the blockade of NMDA receptors and as well as stimulation of opiate receptors.
Dextromethorphan, NMDA receptor antagonist is a safe drug and found to antagonize the morphine withdrawal syndrome in rats (Koyuncuoglu et al., 1990; Koyuncuoglu, 1995). However, the mechanism by which DM attenuates the EW symptoms including hyperalgesia is not known. A study has reported the cross tolerance between morphine and ethanol to their antinociceptive effects (Malec et al., 1987), suggesting the existence of common elements or pathways of development of tolerance and dependence to both these agents. In present study, we could demonstrate that the possible mechanism by which DM attenuates the EW induced hyperalgesia is by antagonizing the NMDA receptors and probably by stimulating opiate receptors resulting in antinociception. Chronic ethanol administration is known to upregulate NMDA receptor function which may contributes to ethanol tolerance and EW symptoms (Krystal et al., 2003). In this study, we examined the effect of co-administration of DM at two different dose levels along with ethanol on EW induced hyperalgesia. Blockade of seizures and decreased locomotor activity during EW by DM demonstrates its utility in the treatment of EW syndrome (Erden et al., 1999). Ethanol and NMDA receptors antagonists share physiologic and rewarding pathways (Krystal et al., 2003). The decreased ethanol consumption in the DM treated groups in the present study demonstrates the role of NMDA receptors and other targets of DM in rewarding pathways. The decrease in basal reaction time in DM treated group may be due to the decreased locomotor activity as compared to that of control group but this possibility is ruled out using chemical induced nocicetion assay. DM attenuated hyperalgesia induced by EW in both thermal and chemical nociception assays suggesting that antinociception activity observed may not be related to the decreased locomotor activity by DM.
In this study, repeated administration of DM not only attenuated the development of hyperalgesia during EW, but also decreased the EW scores and ethanol consumption in the DM treated animals. The blockade of hyperalgesia may not be attributed to any acute antinociceptive effects of DM as the last dose of DM was given 6 h before the EW. Thus the findings suggest that this effect might be due to the changes in the NMDA receptor density and function. However, we cannot rule out the involvement opiate receptors and other unknown mechanisms of antinociceptive effect of DM during EW. Decrease in ethanol intake also might not be responsible for the antinociceptive effect of DM as decrease in ethanol intake was unable to prevent EW induced hyperalgesia and other symptoms in DM treated animals but has only decreased the EW scores at 6 h. Dextromethorphan was able to block hyperalgesia to same extent at both the doses tested during EW, whereas DM treatment decreased ethanol intake dose dependently. This suggests that the blockade of hyperalgesia is not due to the reduced ethanol intake. Activation of mesolimbic dopamine neurons, arising from the cell bodies of the ventral tegmental area, which are projecting to the nucleus accumbens, play an important role in mediating the suppression of tonic pain (Altier and Stewart, 1999). The hyperalgesia seen during EW may be due to an adaptive up regulation of NMDA receptors during chronic alcohol ingestion followed by altered dopaminergic activity in mesolimbic region, which may trigger the response to nociceptive stimuli. DM might have attenuated the hyperalgesia by blocking the upregulated NMDA receptors or by activating the opiate receptors. Further studies are needed to explore and establish the utility of DM in prevention of EW induced hyperalgesia and in the prevention or treatment of EW syndrome. The advantage of DM over the existing treatment of EW is that it can be given orally and is safe with no abuse liability like other opioids (Koyuncuoglu et al., 1990).
To summarize, we have demonstrated that DM blocks the EW induced hyperalgesia and also decreases ethanol consumption, thus suggesting the role for NMDA receptors in rewarding pathways, EW induced hyperalgesia and EW syndrome. Benzodiazepines (BDZ), the treatment option for EW causes sedation and dependence, which are their major limitations. Results from the present study open doors to investigate the use of DM and other NMDA antagonists in the treatment of ethanol addicts and EW syndrome either in combination therapy with BDZ or as monotherapy. The combination therapy of NMDA antagonists and low dose BDZ may have better efficacy and lower side effects, which needs to be investigated. Furthermore, we also hypothesize that DM abuse is more prevalent in alcohol addicts due to the comfortable effects such as attenuation of EW syndrome and hyperalgesia during EW period. But there are no documented reports available even though some limited reports of DM abuse have been reported earlier (Walker and Yatham, 1993; Wolfe and Caravati, 1995).
CONCLUSION
This study demonstrated that dextromethorphan can attenuate hyperalgesia during ethanol withdrawal. The study also provided behavioral evidence in agreement with earlier work on the role of NMDA receptors in the development of EW induced symptoms and hyperalgesia. Further clinical studies are required to establish the treatment options in EW syndromes with NMDA receptor antagonists either as monotherapy or as combination with benzodiazepines.
ACKNOWLEDGMENTS
The experimental work was performed at KS Hegde Medical Academy, Mangalore, India. The study was supported financially by Institute Research Fund, KSHEMA, Mangalore, India. Dextromethorphan pure compound was provided by Wockhardt Pharma Ltd., Mumbai, India.
REFERENCES
- Gatch, M.B. and M. Selvig, 2002. Theophylline blocks ethanol withdrawal-induced hyperalgesia. Alcohol. Alcohol., 37: 313-317.
PubMed - Hoffman, P.L. and B. Tabakoff, 1996. Alcohol dependence: A commentary on mechanism. Alcohol. Alcohol., 31: 333-340.
PubMed - Koyuncuoglu, H., 1995. The combination of tizanidine markedly improves the treatment with dextromethorphan of heroin addicted outpatients. Int. J. Clin. Pharmacol. Ther., 33: 13-19.
PubMed - Koyuncuoglu, H., M. Gungor, H. Sagduyu and F. Aricioglu, 1990. Suppression by ketamine and dextromethorphan of precipitated abstinence syndrome in rats. Pharmacol. Biochem. Behav., 35: 829-832.
PubMed - Li, H.F. and Kendig J.J., 2003. Ethanol withdrawal hyper-responsiveness mediated by NMDA receptors in spinal cord motor neurons. Br. J. Pharmacol., 139: 73-80.
CrossRef - Malec, D., J. Kotlinska and R. Langwinski, 1987. Cross-tolerance between morphine and ethanol and their antinociceptive effects. J. Stud. Alcohol., 48: 507-510.
PubMed - Mao, J. and D.J. Mayer, 2001. Spinal cord neuroplasticity following repeated opioid exposure and its relation to pathological pain. Ann. N.Y. Acad. Sci., 933: 175-184.
PubMed - Narita, M., K. Miyoshi, M. Narita and T. Suzuki, 2007. Changes in function of NMDA receptor NR2B subunit in spinal cord of rats with neuropathy following chronic ethanol consumption. Life Sci., 80: 852-859.
PubMed - Walker, J. and L.N. Yatham, 1993. Benylin (dextromethorphan) abuse and mania. BMJ., 306: 896-896.
PubMed - Wolfe, T.R. and E.M. Caravati, 1995. Massive dextromethorphan ingestion and abuse. Am. J. Emergency Med., 13: 174-176.
PubMed - Yaksh, T.L., X.Y. Hua, I. Kalcheva, N. Nozaki-Taguchi and M. Marsala, 1999. The spinal biology in humans and animals of pain states generated by persistent small afferent input. Proc. Natl. Acad. Sci. USA., 96: 7680-7686.
PubMed