
K.N. Kim
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
Y.M. Ham
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
J.Y. Moon
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
M.J. Kim
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
D.S. Kim
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
W.J. Lee
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
N.H. Lee
Jeju Seaweed Industry Development Agency, Korea
C.G. Hyun
Jeju Biodiversity Research Institute (JBRI), Jeju High-Tech Development Institute (HiDI), Jeju 699-943, Korea
International Journal of Pharmacology
Year: 2009 | Volume: 5 | Issue: 5 | Page No.: 298-306







ABSTRACT
The cytotoxic activity of a variety of Jeju coastal seaweeds against a panel of tumour cell lines was tested in vitro using a calorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Twenty-three extracts were screened against HL-60 (human promyelocytic leukaemia cell line), HT-29 (human colon carcinoma cell line), B16F10 (murine melanoma cell line) and A549 (human lung cancer cell line). The ethyl acetate (EtOAc) extracts of Sargassum thunbergii and Dictyopteris divaricata showed excellent cytotoxic activity against the HL-60 cell line. Furthermore, the S. thunbergii extract also exhibited good cytotoxic activity against the HT-29 and B16F10 cell lines. To explore the mechanisms of cytotoxicity of S. thunbergii and D. divaricata, we used several measures of apoptosis to determine whether these processes were involved in EtOAc fraction-induced HL-60 cell death. We found that EtOAc fractions induced cell shrinkage, cell membrane blebbing and formation of apoptotic bodies. In addition, HPLC fingerprinting of the S. thunbergii extract revealed that fucosterol was a standard component of these fractions. These results suggest that S. thunbergii and D. divaricata have great potential value as food additives, medicinal supplements for patients with chronic diseases and preventive agents against cancer.
PDF Abstract XML References Citation
How to cite this article
K.N. Kim, Y.M. Ham, J.Y. Moon, M.J. Kim, D.S. Kim, W.J. Lee, N.H. Lee and C.G. Hyun, 2009. In vitro Cytotoxic Activity of Sargassum thunbergii and Dictyopteris divaricata (Jeju Seaweeds) on the HL-60 Tumour Cell Line. International Journal of Pharmacology, 5: 298-306.
DOI: 10.3923/ijp.2009.298.306
URL: https://scialert.net/abstract/?doi=ijp.2009.298.306
DOI: 10.3923/ijp.2009.298.306
URL: https://scialert.net/abstract/?doi=ijp.2009.298.306
INTRODUCTION
In the regulation of development and homeostasis in multicellular organisms, cell death is as important as cell proliferation. Although, physiological cell death is usually mediated via apoptosis, uncontrolled growth and proliferation and inhibited apoptosis are major characteristics of cancer cells. Apoptosis is a gene-regulated phenomenon that is induced by many chemotherapeutic agents in cancer treatment. The induction of apoptosis in tumour cells is considered to be useful not only for the management and treatment of cancer, but also for its prevention (Hersey and Zhang, 2001; Taraphdar et al., 2001; Poncet and Kroemer, 2002). A wide variety of natural substances that can induce apoptosis in various tumour cells have been recognized. Thus, it is of considerable importance to be able to isolate apoptotic inducers (either in the form of crude extracts or as components) from bioresources (Afshari et al., 2008). Much previous research has studied the biological effects of natural marine products, including their antioxidant, antibacterial, anti-malarial, antiviral, anti-inflammatory and cytotoxic activities (Skropeta, 2008; Blunt et al., 2009; Gademann and Kobylinska, 2009; Gulder and Moore, 2009; Mayer et al., 2009; Napolitano et al., 2009). However, only a few studies have examined the biological potential of the marine algae (seaweeds) (Afolayan et al., 2008; Li et al., 2008; Jung et al., 2009; Kim et al., 2009; Zhang and Kim, 2009). Jeju Island, the largest island in Korea, is located in the southwest Korea Strait and is famed for its distinctive environment. In particular, the sea levels around this island are known to fluctuate rapidly as a result of global warming. Therefore, in response to this unusual environment, the seaweeds that are present on Jeju Island may possess substantial endogenous protective mechanisms. Thus, the aim of this study was to evaluate the potential of certain Jeju seaweed species as sources of new cytotoxic compounds. To achieve this, the growth of a variety of tumour cell lines, HL-60 (human promyelocytic leukaemia cell line), HT-29 (human colon carcinoma cell line), B16F10 (murine melanoma cell line) and A549 (human lung cancer cell line) were assessed in vitro using the calorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay.
MATERIALS AND METHODS
Preparation of Jeju seaweed extracts: Dried samples of Jeju seaweeds were prepared on Jeju Island from 2006 to 2007 and then identified by Dr. Wook Jae Lee. Previously, sample specimens were deposited at the herbarium of the Jeju Biodiversity Research Institute (JBRI), Jeju, Republic of Korea. The materials for extraction were cleaned, dried at room temperature for 2 weeks and then ground into a fine powder. The dried algae (100 g) were extracted with 80% ethanol (EtOH; 2 L) at room temperature for 24 h and then evaporated in a vacuum. The evaporated EtOH extract was then suspended in water (4 L) and partitioned with ethyl acetate (EtOAc; 4 L); this partitioning was repeated three times.
Cell culture: HL-60, HT-29 and A549 cells were grown in RPMI-1640 medium and B16F10 cells were grown in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% (v/v) heat-inactivated Foetal Bovine Serum (FBS), penicillin (100 U mL-1) and streptomycin (100 μg mL-1). Cultures were maintained in an incubator at 37°C and 5% CO2 (Manthey and Guthrie, 2002).
Cell growth inhibitory assay: The cytotoxicity of each sample against the tumour cells was determined using a calorimetric MTT assay. Suspension cells (HL-60 cells) were seeded (5x104 cells mL-1) together with various concentrations (13, 25 and 50 μg mL-1) of the sample and then incubated for a period of up to 72 h before MTT treatment. Attached cells (HT-29, A549 and B16 cells) were seeded in a 96-well plate at a concentration of 2x104 cells mL-1. The cells were treated with sample (100 μg mL-1) 16 h after seeding and incubated for 72 h at 37°C. MTT stock solution (50 μL; 2 mg mL-1 in PBS) was then added to each well to produce a total reaction volume of 250 μL. After incubating for 4 h, the plates were centrifuged at 2,000 rpm for 10 min and the supernatant was aspirated. The formazan crystals in each well were dissolved in DMSO and the amount of purple formazan was determined by measuring the absorbance at 540 nm (Idibie et al., 2007).
Cytotoxicity assay of HaCaT cells: Human keratinocyte HaCaT cells were cultured in DMEM containing 10% FBS (Gibco) and penicillin-streptomycin at 37°C in a humidified 95% air/5% CO2 atmosphere. Cells were seeded on 48-well plates and drug treatment began 24 h after seeding. After HaCaT cells were incubated with 100 μg mL-1 concentrations of EtOAc fractions of various seaweeds for 24 h at 37°C in 5% CO2 atmosphere, MTT (1 mg mL-1 in PBS) was added to each well in a 1/10 volume of media. Cells were incubated at 37°C for 4 h and DMSO was added to dissolve the formazan crystals. The absorbance was then measured at 570 nm with a spectrophotometer (Power Wave, Bio-tek).
Nuclear staining with Hoechst 33342: The nuclear morphology of the cells was studied using the cell-permeable, DNA-specific fluorescent dye Hoechst 33342. Cells with homogeneously stained nuclei were considered to be viable, whereas the presence of chromatin condensation and/or fragmentation was indicative of apoptosis (Gschwind and Huber, 1995; Lizard et al., 1995). The HL-60 cells were placed in 24-well plate at a concentration of 1x105 cells mL-1. The cells were treated with various concentrations (13, 25, 50 and 100 μg mL-1) of the sample and further incubated for 24 h. Subsequently, Hoechst 33342 was added to the culture medium to give a final concentration of 10 μg mL-1 and the plate was incubated for an additional 10 min at 37°C. To ascertain the degree of nuclear condensation, the stained cells were then observed under a fluorescence microscope equipped with a CoolSNAP-Pro colour digital camera.
Flow cytometry analysis: To determine the proportion of apoptotic sub-G1 hypodiploid cells, flow cytometry analysis was performed (Nicoletti et al., 1991). The HL-60 cells were placed in a 6-well plate at a concentration of 1.0x105 cells mL-1 and treated with various concentrations (13, 25, 50 and 100 μg mL-1) of the sample. After 24 h, the cells were harvested at the indicated time and fixed in 1 mL of 70% ethanol for 30 min at 4°C. The cells were washed twice with PBS and then incubated in the dark in 1 mL of PBS containing 100 μg PI and 100 μg RNase A for 30 min at 37°C. Flow cytometric analysis was performed using a FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA, USA). The effect on the cell cycle was determined by measuring changes in the percentage of the cells engaged in each phase of the cell cycle; from these data, histograms were generated using the computer programmes Cell Quest and Mod-Fit (Wang et al., 1999).
HPLC fingerprint of S. thunbergii and D. divaricata EtOAc extracts: As fucosterol was previously reported to be a seaweed component that is an effective anti-cancer agent (Lee et al., 2003), we searched for this compound in the EtOAc extracts of S. thunbergii and D. divaricata. The chromatographic analyses of S. thunbergii and D. divaricata EtOAc extracts was performed using High-Performance Liquid Chromatography (HPLC) with an AllianceTM Waters 2695 separation module; this was coupled to a Waters 2998 photodiode array detector that utilized a Capcell pak 4.6x250 mm C18 column (particle size 5 μm, Shiseido Chemicals, Tokyo) at a flow rate of 1.0 mL min-1. The column was placed in a column oven at 25°C (Yoon et al., 2009). The mobile phase consisted of acetonitrile (CAN) and water. A gradient program was used as follows: a linear gradient from 70 to 100% ACN in the first 3 min and an isocratic elution with 100% ACN for 47 min.
RESULTS AND DISCUSSION
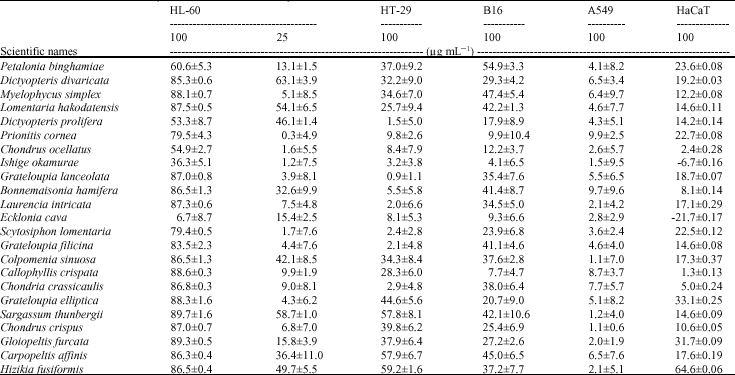
Cytotoxic activities of Jeju seaweeds against cancer cell lines: The algae source, provenance of the algae, harvest time or developmental stage, extraction technique, use of fresh or dried algae material and test methodology are all factors that influence the anti-tumour activity and therefore must be taken into account whenever anti-tumour assays are performed using these seaweeds. In this study, we tested the cytotoxic effects of 23 extracts of seaweeds, collected from Jeju Island, on HL-60, HT-29, B16 and A549 cells. These cells were subjected to 72 h of exposure at either a 25 or 100 μg mL-1 concentration (Table 1). As a control group, some cells were incubated with the same concentration of DMSO. The percentage inhibition of seaweed extracts (25 μg mL-1) from Dictyopteris divaricata, Lomentaria hakodatensis, Dictyopteris prolifera, Colpomenia sinuosa, Sargassum thunbergii and Hizikia fusiformis, on HL-60 cell lines was over 40%. Of the 23 seaweeds, D. divaricata (JBR246) and S. thunbergii (JBR511) displayed the strongest cytotoxic effects on human HL-60. Furthermore, S. thunbergii, Carpopeltis affinis and H. fusiformis also demonstrated cytotoxic effects greater than 50% on HT-29 cell lines. We compared the cytotoxic effects of seaweed extracts on cancer cells with the effects on normal cells (human keratinocytes, HaCaT). As shown in Table 1, with the exceptions of H. fusiformis, Grateloupia elliptica and Gloiopeltis furcata, the viability of human keratinocytes (HaCaT) that were exposed to concentrations up to 100 μg mL-1 for periods of 72 h was barely affected. These results suggest that Jeju seaweeds have irreversible cytotoxic effects on HL-60 cells but not on normal cells. To determine the concentration-dependent effects of JBR246 and JBR511 on the HL-60 cells, the MTT conversion assay was performed. Cells were exposed to various concentrations of JBR246 and JBR511 (13, 25, 50 and 100 μg mL-1) for 72 h. Cells treated with 0.1% DMSO were used as a control. As shown in Fig. 1, both JBR246 and JBR511 exerted a cytotoxic effect on HL-60 cells in a dose-dependent manner. The IC50 values of JBR246 and JBR511, calculated from the graph, were 23.46 and 15.15 μg mL-1, respectively.
| Table 1: | Growth inhibition activity of EtOAc fraction from Jeju seaweeds on tumour cells |
 | |
| The cytotoxic effects of 23 extracts of edible seaweeds, collected from Jeju Island, on HL-60 (human promyelocytic leukaemia cell line), HT-29 (human colon carcinoma cell line), B16F10 (murine melanoma cell line) and A549 (human lung cancer cell line) cells were tested. These cells were subjected to exposure for 72 h at 25 μg mL-1 or 100 μg mL-1 concentrations. The cytotoxic effects of seaweed extracts on cancer cells were compared with the effects on normal cells (human keratinocytes, HaCaT) as a control | |
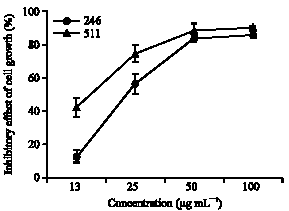
Observation of JBR246- and JBR511-induced morphological changes in HL-60 cells under the fluorescent microscope: To test whether the decrease in cell viability observed after treatment with JBR246 and JBR511 was due to apoptosis, the HL-60 cells were stained with Hoechst 33342 dye. This dye stains the condensed chromatin of apoptotic cells more brightly than it stains the chromatin of normal cells. As shown in Fig. 2, Hoechst staining, which correlates with the presence of cells with a typical apoptotic nuclear morphology (nuclear shrinkage, DNA condensation and fragmentation), was present in the JBR246- and JBR511-treated cells, but not in the untreated controls.
 | |
| Fig. 1: | Inhibitory effect of JBR246 and JBR511 fractions against the growth of HL-60 cells. HL-60 cells were incubated with various concentrations (13, 25, 50 and 100 μg mL-1) of JBR246 and JBR511 extracts for 72 h and the cell viability was examined via MTT assay |
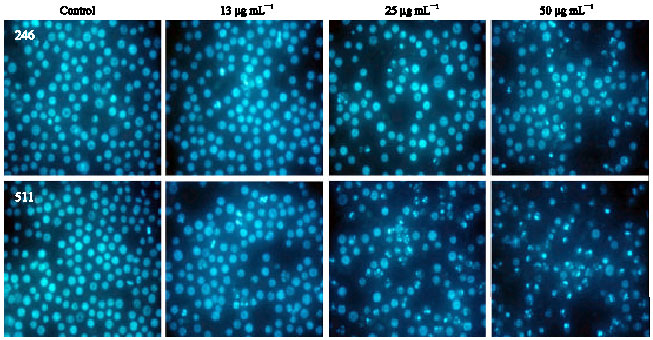
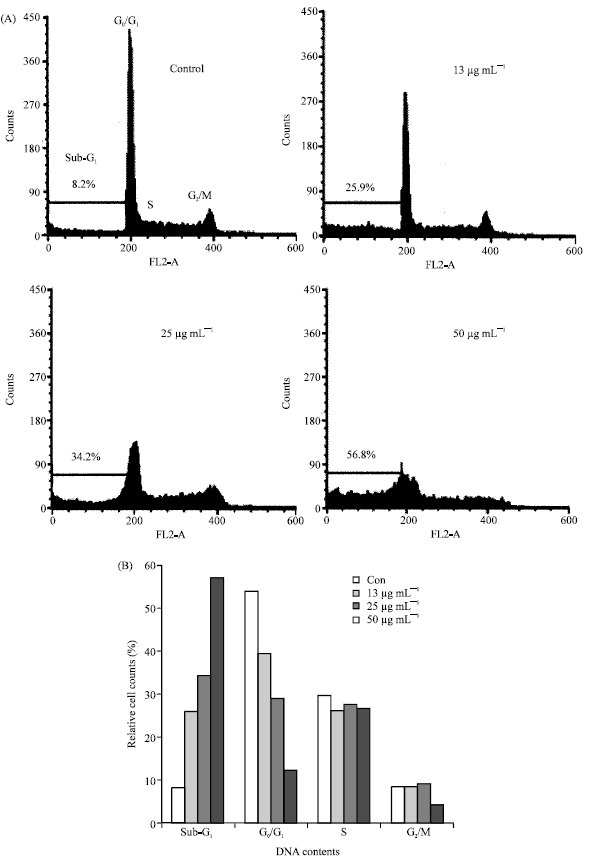
Effects of JBR246 and JBR511 on the population of hypodiploid cells among the HL-60 cells: To test whether JBR246 and JBR511 induced cell death via apoptosis, the percentages of apoptotic cells were measured via flow cytometry. In recent years, flow cytometry has emerged as a rapid and sensitive technique for studying cell membranes and intracellular changes in various cell types (Teng et al., 2006). Flow cytometric analysis was performed to determine the presence of hypodiploid cells. In flow cytometry histograms, apoptotic cells produce DNA fluorescence in the subdiploid regions. Therefore, we used flow cytometric analysis to assess the progression of the cell cycle and to discriminate between cells that were experiencing apoptosis and those that were undergoing necrosis as a result of the lethal effects of JBR246 and JBR511. In HL-60 cells that were treated with JBR246 or with JBR511, there was the appearance of a hypodiploid peak (A0 region), thought to be due to the presence of apoptosing cells and/or apoptotic bodies with a DNA content less than 2n (Fig. 3A, 4A). Quantitation of the apoptotic cells showed a dose-dependent response to exposure to either JBR246 or JBR511 (Fig. 3B, 4B); this result is in accordance with the finding of morphological changes. To determine the percentage of cells in the sub-G1, G0/G1, S and G2/M phases for both the control cells and the cells that were treated with JBR246 and JBR251, quantitative analyses of the histograms for 24 h are shown.
 | |
| Fig. 2: | Effect of JBR246 and JBR511 fractions on morphology of HL-60 cells. HL-60 cells were incubated with various concentrations (13, 25 and 50 μg mL-1) of the JBR246 and JBR511 fractions for 8 h. Cells were stained with Hoechst 33342 and the stained nuclei were imaged under a fluorescent microscope |
 | |
| Fig. 3: | Effect of the JBR246 fraction on the cell cycle pattern and on the fraction of apoptotic cells in the population of HL-60 cells, as measured by flow cytometric analysis. HL-60 cells were treated with various concentrations (13, 25 and 50 μg mL-1) of JBR246 for 72 h. A sub-G1 peak, which is indicative of apoptotic cells was induced in JBR246-treated cells but not in control cells. (A) Histogram of cell cycle patterns of HL-60 cells. (B) Bar graph of cell cycle patterns of HL-60 cells |
 | |
| Fig. 4: | Effect of the JBR511 fraction on the cell cycle pattern and percentage of HL-60 cells undergoing apoptosis, as measured by flow cytometric analysis. HL-60 cells were treated with various concentrations (13, 25 and 50 μg mL-1) of JBR511 for 72 h. A sub-G1 peak, which is indicative of apoptotic cells was induced in JBR511-treated but not in control cells. (A) Histogram of cell cycle patterns of HL-60 cells. (B) Bar graph of cell cycle patterns of HL-60 cells |
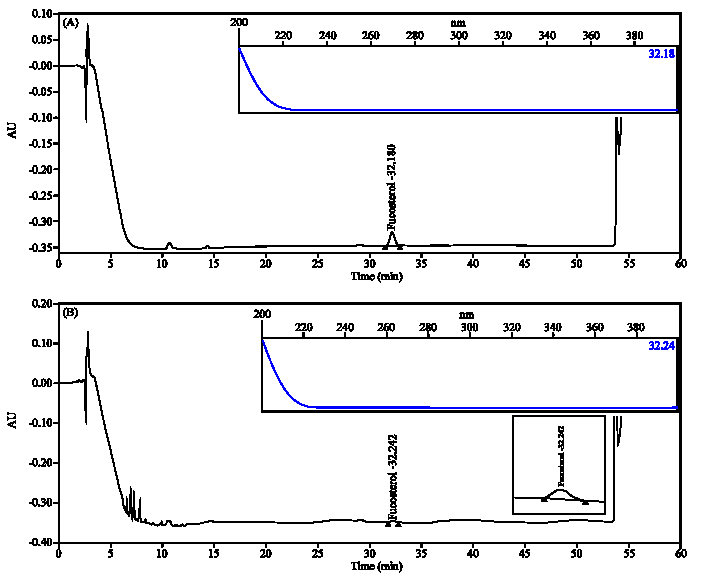 | |
| Fig. 5: | HPLC fingerprinting analysis of the JBR511 EtOAc extract. HPLC profiles of authentic fucosterol and JBR511 EtOAc extract are shown in (A) and (B), respectively |
These data clearly indicate that JBR246 and JBR511 cause an increase in the percentage of cells at the sub-G1 phase and a significant decrease in the percentage of cells at the G0/G1 phases (Fig. 3A, 4A). Present results suggest that cell cycle-induced apoptosis is a mechanism by which JBR246 and JBR511 exert their antiproliferative effects.
HPLC fingerprint of the JBR246 and JBR511 extracts: According to traditional oriental medicine, the therapeutic actions of herbal medicines are based on the integrated interaction of many different ingredients. With the development of analytical technology, the use of chromatographic methods to develop fingerprints for traditional oriental medicines and their raw materials is highly recommended. Thus, interest in HPLC fingerprint analysis has increased not only in Asia, but also in other countries around the world (Zhang et al., 2007; Yoon et al., 2009). Therefore, simple HPLC fingerprints of the algae were explored in this study. In accordance with previous studies that confirmed that fucosterol is a seaweed component with effective anti-tumour properties, substantial amounts of this substance were detected. Using the conditions described in the experimental section, fucosterol was well resolved (with excellent peak shapes) from the JBR511 EtOAc extracts (Fig. 5). The fucosterol content in the EtOAc extracts was 15 ppm. Thought, it was previously reported that D. divaricata harbours several bio-active compounds in addition to fucosterol, including dictyvaric acid, 1,4-epoxymuurolan-5beta-ol, (-)-torreyol, 4beta, 5alpha-dihydroxycubenol, 3-farnesyl-p-hydroxybenzoic acid, chromazonarol, phenyl acetylamine, 4-hydroxybenzoic acid and n-hexadecanoic acid (Song et al., 2005, 2006; Wen et al., 2009). In contrast, fucosterol was not detected in the JBR246 EtOAc extracts. This can be attributed to the presence of only trace amounts of fucosterol or to seasonal and regional differences. Further investigations will focus on the in vivo assessment of the biological activity of the JBR246 and JBR511 extracts and on the chemical identification of the major active components responsible for anti-tumour activity in the efficacious extracts. To the best of our knowledge, this is the first report demonstrating the in vitro anti-cancer activity of the EtOAc extracts of S. thunbergii and D. divaricata and providing a scientific basis for its effects on human health.
CONCLUSION
JBR246 and JBR511 could decrease cell viability in HL-60 cells in a concentration-dependent manner. The IC50 values of JBR246 and JBR511 after 72 h were 23.46 and 15.15 μg mL-1, respectively. Both JBR246 and JBR511 induced a sub-G1 peak in the flow cytometry histogram of treated cells compared to control cells, indicating that apoptotic cell death is involved in JBR246- and JBR511-induced toxicities. It might be concluded that JBR246 and JBR511 could cause cell death in HL-60 cells, in which apoptosis or programmed cell death plays an important role. JBR246 and JBR511 could be also considered as a promising chemotherapeutic agent in cancer treatment in the future. As the amounts of JBR246 and JBR511 used in this study were large, it is possible that the active agents represent only a small component of the extracts. Therefore, further studies are required in order to fractionate the active components and to elucidate the exact mechanism of action.
ACKNOWLEDGMENTS
This research was supported in part by the Program for the RIS (Regional Innovation System; JEJU SEA-GREEN PROJECT) and the Regional Technology Innovation Program (RTI04-02-07), both of which are managed by the Ministry of Knowledge and Economy, Korea.
REFERENCES
- Afolayan, A.F., J.J. Bolton, C.A. Lategan, P.J. Smith and D.R. Beukes, 2008. Fucoxanthin, tetraprenylated toluquinone and toluhydroquinone metabolites from Sargassum heterophyllum inhibit the in vitro growth of the malaria parasite Plasmodium falciparum. Zeitschrift Naturforschung C, 63: 848-852.
CrossRefPubMedDirect Link - Blunt, J.W., B.R. Copp, W.P. Hu, M.H. Munro, P.T. Northcote and M.R. Prinsep, 2009. Marine natural products. Nat. Prod. Rep., 26: 170-244.
PubMed - Gademann, K. and J. Kobylinska, 2009. Antimalarial natural products of marine and freshwater origin. Chem. Rec., 9: 187-198.
CrossRefPubMedDirect Link - Gschwind, M. and G. Huber, 1995. Apoptotic cell death induced by β-amyloid 1-42 peptide is cell type dependent. J. Neurochem., 65: 292-300.
PubMed - Gulder, T.A. and B.S. Moore, 2009. Chasing the treasures of the sea bacterial marine natural products. Curr. Opin. Microbiol., 12: 252-260.
PubMed - Hersey, P. and X.D. Zhang, 2001. How melanoma cells evade trail-induced apoptosis. Nat. Rev. Cancer, 1: 142-150.
PubMed - Idibie, C.A., H. Davids and S.E. Iyuke, 2007. Cytotoxicity of purified cassava linamarin to a selected cancer cell lines. Bioprocess Biosyst. Eng., 30: 261-269.
CrossRefDirect Link - Jung, W.K., Y.W. Ahn, S.H. Lee, Y.H. Choi and S.K. Kim et al., 2009. Ecklonia cava ethanolic extracts inhibit lipopolysaccharide-induced cyclooxygenase-2 and inducible nitric oxide synthase expression in BV2 microglia via the MAP kinase and NF-kappaB pathways. Food Chem. Toxicol., 47: 410-417.
PubMed - Kim, M.M., N. Rajapakse and S.K. Kim, 2009. Anti-inflammatory effect of Ishige okamurae ethanolic extract via inhibition of NF-kappaB transcription factor in RAW 264.7 cells. Phytother. Res., 23: 628-634.
PubMed - Lee, S., Y.S. Lee, S.H. Jung, S.S. Kang and K.H. Shin, 2003. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res., 26: 719-722.
PubMed - Li, Y., S.H. Lee, Q.T. Le, M.M. Kim and S.K. Kim, 2008. Anti-allergic effects of phlorotannins on histamine release via binding inhibition between IgE and Fc epsilonRI. J. Agric. Food Chem., 24: 12073-12080.
PubMed - Lizard, G., S. Fournel, L. Genestier, N. Dhedin and C. Chaput et al., 1995. Kinetics of plasma membrane and mitochondrial alterations in the cells undergoing apoptosis. Cytometry, 21: 275-283.
PubMed - Manthey, J.A. and N. Guthrie, 2002. Antiproliferative activities of citrus flavonoids against six human cancer cell lines. J. Agric. Food Chem., 50: 5837-5843.
PubMed - Mayer, A.M., A.D. Rodriguez, R.G. Berlinck and M.T. Hamann, 2009. Marine pharmacology in 2005-6: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal antituberculosis and antiviral activities affecting the cardiovascular immune and nervous systems and other miscellaneous mechanisms of action. Biochim. Biophys. Acta., 1790: 283-308.
PubMed - Napolitano, J.G., A.H. Daranas, M. Norte and J.J. Fernandez, 2009. Marine macrolides a promising source of antitumor compounds. Anticancer Agents Med. Chem., 9: 122-137.
PubMed - Nicoletti, I., G. Migliorati, M.C. Pagliacci, F. Grignani and C. Riccardi, 1991. A rapid and simple method for measuring thymocyte apoptosis by propidium iodide staining and flow cytometry. J. Immunol. Meth., 139: 271-279.
CrossRefPubMedDirect Link - Poncet, K.M. and G. Kroemer, 2002. Chemotherapy targeting the mitochondrial cell death pathway. Oncogene, 21: 8786-8803.
PubMed - Song, F.H., X. Fan, X.L. Xu, J.L. Zhao, J.J. Han and J.G. Shi, 2005. Chemical constituents of the brown alga dictyopteris divaricata. J. Asian Nat. Prod. Res., 7: 777-781.
PubMed - Song, F.H., X. Fan, X.L. Xu, S.J. Wang, S. Li, Y.C. Yang and J.G. Shi, 2006. Studies on chemical constituents of the brown alga dictyopteris divaricata. Zhongguo Zhong Yao Za Zhi., 31: 125-128.
PubMed - Taraphdar, A.K., M. Roy and R.K. Bhattacharya, 2001. Natural products as inducers of apoptosis: Implication for cancer therapy and prevention. Curr. Sci., 80: 1387-1396.
Direct Link - Afshari, J.T., A. Brook and S.H. Mousavi, 2008. Study of cytotoxic and apoptogenic properties of saffron extract in human cancer cell lines. Food Chem. Toxicol., 46: 3443-3447.
PubMed - Teng, B.S., Y.H. Lu, Z.T. Wang, X.Y. Tao and D.Z. Wei, 2006. In vitro anti-tumor activity of isorhamnetin isolated from Hippophae rhamnoides L. against BEL-7402 cells. Pharmacol. Res., 54: 186-194.
PubMed - Wang, X.W., Q. Zhan, J.D. Coursen, M.A. Khan and H.U. Kontny et al., 1999. GADD45 induction of a G 2 M cell cycle checkpoint. Proc. Natl. Acad. Sci. USA., 96: 3706-3711.
PubMed - Wen, W., F. Li, N.Y. Ji, X.M. Li and C.M. Cui et al., 2009. A new cadinane sesquiterpene from the marine brown alga dictyopteris divaricata. Molecules, 14: 2273-2277.
PubMed - Yoon, W.J., Y.M. Ham, B.S. Yoo, J.Y. Moon, J. Koh and C.G. Hyun, 2009. Oenothera laciniata inhibits lipopolysaccharide induced production of nitric oxide prostaglandin E2 and proinflammatory cytokines in RAW264.7 macrophages. J. Biosci. Bioeng., 107: 429-438.
PubMed - Zhang, C. and S.K. Kim, 2009. Matrix metalloproteinase inhibitors (MMPIs) from marine natural products the current situation and future prospects. Mar. Drugs., 7: 71-84.
PubMed - Zhang, X.Y., J.Y. Tao, L. Zhao, Z.J. Huang and F.L. Xiong et al., 2007. In vitro anti-inflammatory effects of different solution fractions of ethanol extract from Melilotus suaveolens Ledeb. Chinese Med. J., 120: 1992-1998.
PubMed