
C. S. Nworu
Department of Pharmacology and Toxicology, Faculty ofPharmaceutical Sciences, University of Nigeria, Nsukka, 410001 Enugu State, Nigeria
P. A. Akah
Department of Pharmacology and Toxicology, Faculty ofPharmaceutical Sciences, University of Nigeria, Nsukka, 410001 Enugu State, Nigeria
C. O. Okoli
Department of Pharmacology and Toxicology, Faculty ofPharmaceutical Sciences, University of Nigeria, Nsukka, 410001 Enugu State, Nigeria
C. O. Esimone
Department of Pharmaceutics, Faculty of Pharmaceutical Sciences, University of Nigeria, Nsukka, 410001 Enugu State, Nigeria
F. B.C. Okoye
Department of Medicinal Chemistry, Faculty ofPharmaceutical Sciences, University of Nigeria, Nsukka, 410001 Enugu State, Nigeria
International Journal of Pharmacology
Year: 2007 | Volume: 3 | Issue: 4 | Page No.: 347-351







ABSTRACT
The modulatory activity of methanolic seed extracts of Garcinia kola (ME) on Delayed-Type Hypersensitivity (DTH) response, primary and secondary humoral responses and on in vivo leucocytes mobilisation were evaluated. Acute toxicity test of the extract was also carried out. The ME at 250 and 500 mg kg-1 body weight produced significant (p<0.05) inhibition of DTH response in mice by 67.40 and 53.29%, respectively. Primary and secondary sheep red blood cells-specific antibody titres were significantly elevated when compared to the control group. Agar-induced in vivo leucocytes mobilisation into the mice peritoneal fluid was significantly (p<0.05) increased by ME at 250 and 500 mg kg-1. The total leucocytes counts were higher in the extract-treated groups when compared to the control group. The mobilised white blood cells were predominantly polymorphonuclear neutrophils (PMNs). The ME administered (orally) at 5000 mg kg-1 did not caused lethality and signs of acute intoxication after 48 h observation period. The results of this study have established cellular and humoral immunomodulatory activities of G. kola extract and justify further investigations into the effects of specific constituents of the plant on immune system components.
PDF Abstract XML References
How to cite this article
C. S. Nworu, P. A. Akah, C. O. Okoli, C. O. Esimone and F. B.C. Okoye, 2007. The Effects of Methanolic Seed Extract of Garcinia kola on Some Specific and Non-Specific Immune Responses in Mice. International Journal of Pharmacology, 3: 347-351.
DOI: 10.3923/ijp.2007.347.351
URL: https://scialert.net/abstract/?doi=ijp.2007.347.351
DOI: 10.3923/ijp.2007.347.351
URL: https://scialert.net/abstract/?doi=ijp.2007.347.351
INTRODUCTION
The stress and pressure of modern society take a toll on immune system. Those with weakened immunity are more susceptible to infection and disease. The need to maintain or rebuild a healthy defense has led researchers to minerals, plants and fungi in search of natural substances with health-supporting properties. Compounds that are capable of interacting with the immune system to up regulate or down regulate specific aspects of the host response are classified as immunomodulators.
Those compounds which appear to stimulate the human immune response are being sought for the treatment of cancer, immunodeficiency diseases, or for generalised immunosuppression following drug treatment; for combination therapy with antibiotics and as adjuncts for vaccines (Jong et al., 1983; Jong and Birmingham, 1992). Those compounds that suppress immune reactions are potentially useful in the remedy of autoimmune (an abnormal immune response against self-antigens) or certain gastro-intestinal tract diseases (e.g., Crohns) (Badger, 1983). The human immune system is highly complex and consist of two categories of defense mechanisms-the innate (non-specific) and the adaptive (specific) systems (Atal et al., 1986; Guyton and Hall, 2006). These two mechanisms could be modified by substances to either enhance or suppress their ability to resist invasion by pathogens (William, 2001).
There has been a growing interest in identifying and characterising natural compounds with immunomodulatory activities (Wang et al., 1991; Sharma et al., 1994; Lee et al., 1995; Ganachari et al., 2004). Garcinia kola Heckel (Fam: Guttiferae) seed is a plant reputed for a number of therapeutic uses including, anti-inflammatory (Iwu and Igboko, 1982), antidiabetic (Iwu et al., 1990), hypolypidaemic (Adaramonye and Adeyemi, 2006), antioxidant (Farombi, 2002; Adaramonye et al., 2005), antimicrobial (Hussain, 1982; Iwu, 1999), antihepatotoxic (Iwu et al., 1987; Braide, 1991) and antiviral properties (Iwu, 1999). Most of these pharmacological activities are believed to be related to its antioxidant properties and to its ability to increase immunity in general (Okonji et al., 1999).
Garcinia kola is a medium sized tree found in moist forest and widely distributed throughout west and central Africa (Hutchinson and Dalziel, 1956). The plant is highly valued in these countries for its edible nuts. The seed, commonly known as bitter kola, is a masticatory agent and is a major kola substitute offered to guests at home and shared at social ceremonies. Although, there is no data on the effects of Garcinia kola on the immune system, some herbal products now have the plant extract included as immune-boosting agent (Meserole, 1999). The present study was conducted to determine the effects of the seed extracts of G. kola on the body defence mechanisms. In this study, we are presenting our initial findings on the effects of this extract on humoral and cell mediated immune responses.
MATERIALS AND METHODS
Preparation and extraction of plant material: Seeds of Garcinia kola Heckel (Guttiferae) were obtained commercially from a local market-Oba Nsukka, Enugu State, Nigeria in the month of June, 2006. The plant material was authenticated by Mr A. Ozioko, a plant taxonomist of Bioresources Development and Conservation Programme (BDCP) Center, Nsukka. The seeds were peeled, air-dried and pulverised. The powdered seed (500 g) was extracted with methanol in a soxhlet extractor for 24 h, concentrated in a rotary evaporator and yielded residue of 24.4% (w/w). The residue was subjected to phytochemical analysis using the procedures outlined by Harborne (1984), acute toxicity test (LD50) by Lorke method (1983) and its effects on some specific and non-specific immune responses in mice were determined.
Animals: Adult Swiss albino mice (19-25 g) of both sexes obtained from the Animal House of the Department of Pharmacology and Toxicology, University of Nigeria, Nsukka were used in the study. The animals were housed under standard conditions (25±2°C and 12 h light/dark cycle). They were fed with standard pellets (Guinea Feed Nigeria Ltd.) and had unrestricted access to clean drinking water.
Antigen: Fresh sheep blood was obtained from the animal farm of the Faculty of Veterinary Medicine, University of Nigeria, Nsukka. Sheep red blood cells (SRBCs) were washed three times in a copious volume of pyrogen-free sterile normal saline by centrifugation at 3000 x g for 10 min on each occasion. The washed SRBCs was adjusted to a concentration of 109 cells mL-1 for immunization and challenge.
Delayed Type Hypersensitivity (DTH) reaction: Delayed type hypersensitivity was induced in mice using Sheep Red Blood Cells (SRBC) as antigen. Animals were sensitized by subcutaneous injection of 0.02 mL of 109 cells mL-1 SRBC (day 0) in the plantar region of right hind foot paw and challenged on day 5 by subcutaneous injection of the same amount of antigen into the left hind pad. The oedema produced by antigenic challenge in the left hind paw was measured as the difference in the paw thickness before and 24 h after the challenge. The paw thickness was measured with a pocket-sized screw gauge. (Naved et al., 2005) Garcinia kola extract, ME (100, 250 and 500 mg kg-1) was administered 3 days prior to sensitization and continued till the challenge. (Naved et al., 2005; Shinde et al., 1999).
Humoral Antibody (HA) synthesis: Mice were immunized by an intraperitoneal injection (i.p.) of 0.1 mL of 109 SRBC mL-1 on day 0 and challenged by similar i.p. injection of the same amount on day 5. Primary antibody titre was determined on day 5 (before the challenge) and secondary titre on day 10 (Sharma et al., 1996) by the haemagglutination technique (Nelson and Mildenhall, 1967). The ME (100, 250 and 500 mg kg-1) was administered 3 days prior to immunization and continued daily for 5 days after the challenge. Blood samples were obtained by retro-orbital puncture in test tubes and allowed to clot. For each sample, a 25 μL serum was obtained after centrifugation and serially diluted two-fold in 96-U well microtitre plates using pyrogen-free sterile normal saline. The last well on each row contained sterile normal saline as control. The diluted sera were challenged with 25 μL of 1% (v/v) SRBC in the plates and then incubated at 37°C for 1 h. The highest dilution giving rise to visible haemagglutination was taken as antibody titre. Antibody titres were expressed in graded manner, the minimum dilution (1/2) being ranked as 1 (Calculated as-Log2 of the dilution factor). The mean ranks of different treatment groups were compared for statistical significance.
In vivo leucocytes mobilisation: The effect of the ME on in vivo leucocytes migration induced by inflammatory stimulus was investigated using the methods of Ribeiro et al. (1991). One hour after oral administration of the ME (100, 250 and 500 mg kg-1), each mice in the groups (n = 5) received intraperitoneal injections of 0.5 mL of 3% (w/v) agar suspension in normal saline. Four hours later, the mice were sacrificed and the peritoneum washed with 5 mL of a 5% solution of EDTA in Phosphate Buffered Saline (PBS). The peritoneal fluid was recovered and total and differential leucocytes counts (TLC and DLC) were performed on the perfusates.
Statistical analysis: Results were analysed using one way Analysis of Variance (ANOVA; Fischer LSD post hoc test) and expressed as mean±standard error of mean. Differences between means of treated and control groups were considered significant at p<0.05.
RESULTS
Phytochemical studies on ME show the presence of glycosides, sugars, flavonoids, tannins, saponins, sterols and triterpenoids. ME administered orally at 500 mg kg-1 did not cause lethality and signs of acute intoxication after 48 h observation period. The ME at 100, 250 and 500 mg kg-1 produced an inhibition DTH response in mice. This inhibition was produced in a dose-dependent manner and was significant (p<0.05) at 250 and 500 mg kg-1 (Table 1). The extract caused dose-related significant (p<0.05) elevation of primary and secondary SRBCs-specific antibody titre at 250 and 500 mg kg-1 compared with control (Table 2). The humoral antibody stimulation caused by ME at 250 and 500 mg kg-1 is comparable to that produced by the standard immunostimulant drug (levamisol, 2.5 mg kg-1).
The methanolic seed extracts of G. kola (500 mg kg-1) caused an increase in peritoneal leucocyte mobilization by 254.24% when compared to the control group (Table 3).
| Table 1: | The effect of ME on delayed type hypersensitivity reaction in mice |
 | |
| *: p<0.05; n = 5; negative sign shows stimulation of Delayed Type Hypersensitivity (DTH), ME = Methanolic Extract of G. kola | |
| Table 2: | The effects of ME on primary and secondary humoral immune response in mice |
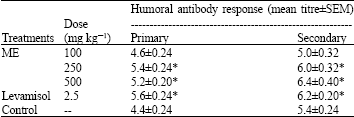 | |
| *: p<0.05; n = 5; ME = Methanolic Extract of G. kola | |
The proportion of neutrophils in the peritoneal perfusates was generally higher than lymphocytes in all the groups, but decreased with increase in dose of ME administered (Table 3).
DISCUSSION
High profile immune-destructive diseases such as AIDS and the concern about bioterrorism are leading consumers to seek natural ways to boost their immune systems. Essentially, the immune system is the body's means of surveillance, intended to protect the body from disease by searching and destroying any health-damaging agents. There are two aspects of immune protection-the innate response and the adaptive response (Atal et al., 1986; Guyton and Hall, 2006). Innate immunity is present at birth and provides the first barrier against microorganisms. Adaptive immunity is the second barrier against infection. It is acquired later in life and retains a memory of all the invaders it has faced. Interestingly, the innate and adaptive mechanisms could be modified by substances to either enhance or suppress their ability to resist invasion by pathogens (William, 2001).
Garcinia kola seed is a social masticatory agent used at ceremonies and presented to guests in several communities of south-eastern Nigeria. G. kola seed is rich in biflavonoids and has been speculated to stimulate the immune system because of its antioxidant and other related activities demonstrated by the seed extracts. (Okonji et al., 1999; Farombi, 2002; Adaramonye et al., 2005). Although there are no research reports supporting the immunomodulatory activities of G. kola seed, the seed is still chewed in the belief that it boosts the immune system. We investigated the effects of methanolic seed extracts of G. kola on cell-mediated and humoral immune responses in mice.
In this study, the manifestation of delayed type hypersensitivity reaction induced by sheep erythrocytes was inhibited by the extract in a dose-related manner. DTH is mediated by interferon-gamma (IFN-γ)-producing CD4+ (TH1) or CD8+ T cells (TC1) (Biedermann et al., 2001).
| Table 3: | The effect of ME on in vivo leucocyte mobilisatrion in mice |
 | |
| *: p<0.05, n = 5; TLC is Total Leucocyte Count; ME = Methanolic Extract of G. kola | |
It usually takes 24-72 h to develop and involves activation of T-cells, which results in the infiltration into the area of inflammation by monocytes and lymphocytes. DTH is known to be initiated by reaction between antigen-specific T cells and the antigen which results in the release of lymphokines that affect a variety of cell types, especially macrophages (Furr, 1998). From present results, the inhibition of DTH by G. kola extract is an indication of its ability to modulate cell-mediated immune response. This mechanism may be related to the anti-inflammatory properties of the plant which has been reported in an earlier study (Iwu and Igboko, 1982). This inhibition can occur by immune deviation which entails steering T-cells towards an IL-4 producing TH2 or TC2 phenotypes (Biedermann et al., 2001).
Administration of the extract equally caused an elevation of primary and secondary humoral immune response to sheep red cell antigen in mice. Antibody synthesis requires the co-operation of at least three major cell types, the macrophages, the B-lymphocytes and T-lymphocytes (Benecerral, 1978). The secondary titres are expectedly higher, since subsequent antigenic stimulation of priory-sensitized animals may result in high antibody production, as there is now an expanded clone of cells with memory of the original antigen available to proliferate into mature plasma cells (Furr, 1998). This property will enhanced humoral immune protection of the animal which is mediated through opsonization, direct neutralization of antigen, agglutination of antigen and activation of complement system to cause lyses and death of antigenic cells (Green and Harris, 1996).
It has been observed that the chemotactic movement of neutrophils towards the foreign body is the first and the most important step in phagocytosis (Ganachari et al., 2004). A significant increase in agar-induced leucocytes mobilisation into the peritoneum was recorded upon treatment with ME. This activity may help to increase the general resistance of the body against microbial infections. The polymorphonuclear neutrophils (PMNs), which engulf and eliminate invading microorganism, was the most mobilised leucocytes.
The results of this studies revealed that the methanolic seed extracts of G. kola has immunomodulatory effects on both the cell-mediated and humoral components of the immune system. This activity may partly explain the general acceptance and high success rates claimed in ethnomedicinal uses of G. kola seed in inflammatory and infective conditions. (Hussain et al., 1982; Iwu and Igboko, 1982; Iwu et al., 1987; Braide, 1991; Iwu, 1999). Although our preliminary phytochemical studies on ME showed the presence of glycosides, sugars, flavonoids, tannins, saponins, sterols and triterpenoids, we are yet to associate the observed activities with specific constituents of the seed.
CONCLUSION
The results of this study have established cellular and humoral immunomodulatory activities of G. kola extract and justify its inclusion in herbal tonics as immune-boosting agent.
REFERENCES
- Adaramonye, A.O., E.O. Farombi, E.O. Adeyemi and G.O. Emerole, 2005. Comparative study on antioxidant properties of flavonoids of Garcinia kola seeds. Pak. J. Med. Sci., 21: 331-339.
Direct Link - Adaramoye, O.A. and E.O. Adeyemi, 2006. Hypoglycaemic and hypolipidaemic effects of fractions from kolaviron, a biflavonoid complex from Garcinia kola in streptozotocin-induced diabetes mellitus rats. J. Pharm. Pharmacol., 58: 121-128.
PubMed - Biedermann, T.R., A. Mailhammer, C. Mai, A. Sander and F. Ogilivie et al., 2001. Reversal of established delayed-type hypersensitivity reactions following therapy with IL-4 or antigen specific Th2 cells. Eur. J. Immunol., 5: 1582-1591.
Direct Link - Braide, V.B., 1991. Inhibition of drug metabolism by flavonoid extract (Kolaviron) of Garcinia kola seeds in the rat. Phytother. Res., 5: 38-40.
Direct Link - Ganachari, M.S., S. Kumar and K.G. Bhat, 2004. Effect of Ziziphus jujuba leaves extract on phagocytosis by human neutrophils. J. Nat. Remedies, 4: 47-51.
Direct Link - Iwu, M.M., O.A. Igboko, C.O. Okunji and M.S. Tempesta, 1990. Antidiabetic and aldose reductase activities of biflavanones of Garcinia kola. J. Pharm. Pharmacol., 42: 290-292.
CrossRef - Jong, S.C. and J.M. Birmingham, 1992. Medicinal benefits of the mushroom Ganoderma. Adv. Applied Microbiol., 37: 101-134.
Direct Link - Lorke, D., 1983. A new approach to practical acute toxicity testing. Arch. Toxicol., 54: 275-287.
CrossRefPubMedDirect Link - Nelson, D.A. and P. Mildenhall, 1967. Studies on cytophilic antibodies l. The production by mice of macrophage cytophilic antibodies to sheep erythrocytes: Relationship to the production of other antibodies and development of delayed type hypersensitivity. Aust. J. Exp. Biol. Med. Sci., 45: 113-130.
CrossRefDirect Link - Ribeiro, R.A., C.A. Flores, F.Q. Cunha and S.H. Ferreira, 1991. IL-8 causes in vivo neutrophil migration by a cell dependent mechanism. Immunology, 73: 472-477.
PubMed - Sharma, M.L., C.S. Rao and P.L. Duda, 1994. Immunostimulatory activity of Picrorhiza kurroa leaf extract. J. Ethnopharmacol., 41: 185-192.
PubMed - Sharma, M.L., B. Singh, B.K. Chandan, A. Khajuria and A. Kaul et al., 1996. Actions of some flavonoids on specific and non-specific immune mechanisms. Phytomedicine, 3: 191-195.
CrossRefPubMedDirect Link - Shinde, U.A., A.S. Phadke, A.M. Nair, A.A. Mungantiwar, V.S. Dikshit and M.M. Saraf, 1999. Preliminary studies on the immunomodulatory activity of Cedrus deodara wood oil. Fitoterapia, 79: 333-339.
Direct Link - Wang, J.Z., X.J. Mao, H. Ito and K. Shimura, 1991. Immunomodulatory activity of polysaccharide from Acanthopanex obovatus roots. Planta Medica, 57: 335-336.
Direct Link - William, J.E., 2001. Review of antiviral and Immunomodulating properties of plants of the Peruvian rainforest. Alter. Med. Rev., 6: 567-579.
Direct Link
Njoku Reply
very good observations
Great economy of the language and god delivery of outcome.