
P. Sahitya Chetan
Division of Molecular Biology, Department of Zoology, S. V. Umversity, Tirupati, India
B. S. Ramakrishna
Department of Animal Sciences, University of Hyderabad, Hyderabad, India
P. Reddanna
Department of Animal Sciences, University of Hyderabad, Hyderabad, India
P. Sangeetha Lakshmi
Department of Pharmaceutical Sciences, S.P. Mahila Umversity, Tirupati, India
P. Murali Mohan
Department of Sericulture, S.P. Mahila University, Tirupati, India
W. Rajendra
Division of Molecular Biology, Department of Zoology, S. V. Umversity, Tirupati, India
International Journal of Pharmacology
Year: 2007 | Volume: 3 | Issue: 4 | Page No.: 341-346







ABSTRACT
In the present investigation, changes in Na+/K+-, Mg2+- and Ca2+-ATPase enzyme activities in different areas of rat brain were examined under the administration of the synthetic opioid analgesic drug tramadol in the absence of induction of pain. Male adult Wistar rats were used as the experimental animals with five groups of six rats each, housed in separate cages. Tramadol was injected subcutaneously into the rats and the changes in enzyme activities in different brain areas were followed at 3, 6, 12 and 24 h following drug administration. The enzyme activities showed a decrease in all areas of the brain following administration of single dose of tramadol and by 24 h they returned more or less to the respective control levels. In the brain of control rats, pons-medulla registered the highest Na+/K+-ATPase activity. Following tramadol administration, maximal decrease was recorded in cortex at 6 h. The Mg2+-ATPase activity in control rats was found to be highest in cortex. On tramadol dosing, maximal decrease was recorded in hypothalamus and cortex. Ca2+-ATPase activity was found to be highest in cortex in the brain of control rats. The activity of this enzyme showed a decrease in all areas of the brain following tramadol administration, with maximal decrease in hippocampus at 3 h. Minor deviations from the respective controls at this period were negligible and not statistically significant. The results indicate that the administration of tramadol during non-induction of pain would induce decrement in the activities of the ATPase enzymes, indicating its impact on energy metabolism and membrane transport functions.
PDF Abstract XML References
How to cite this article
P. Sahitya Chetan, B. S. Ramakrishna, P. Reddanna, P. Sangeetha Lakshmi, P. Murali Mohan and W. Rajendra, 2007. Tramadol Effects on the Activity Levels of ATPases in Mitochondrial Fractions of Rat Brain Areas During Non-Induction of Pain. International Journal of Pharmacology, 3: 341-346.
DOI: 10.3923/ijp.2007.341.346
URL: https://scialert.net/abstract/?doi=ijp.2007.341.346
DOI: 10.3923/ijp.2007.341.346
URL: https://scialert.net/abstract/?doi=ijp.2007.341.346
INTRODUCTION
The understanding of pathology of pain is uncertain and symptoms multiply and diversify until they become disease. Patients have such diverse and complex problems (Twycross and Fairfield, 1982; Turk and Flor, 1984; Banning et al., 1991a, b).
Pain perception is modulated by a variety of opioids through changes in neurotransmitters including nor-epinephrine and serotonin (Yaksh, 1988). In the clinical setting, analgesia may be induced by obtunding or interrupting the nociceptive process at one or more points between the peripheral nociceptor and the cerebral cortex (Kohn et al., 1997).
Different mechanisms of pain can be associated with different opioid receptor subtypes (Schmauss et al., 1983; Pasternak, 1993), so that variable responses to opioids might occur (Portenoy et al., 1990). Also, pain-related factors are involved in the response variability to analgesics, like poor responsiveness of neuropathic pain to opioids compared with nociceptive pain (Arner and Meyerson, 1988; Portenoy et al., 1990). Opioids constitute one of the largest, most widely used and most effective classes of drugs employed for the treatment of moderate to severe pain (Gutstein and Akhil, 2001).
Adenosine Triphosphatases (ATPases) play an important role as energy transducers by coupling ATP hydrolysis with energy processes (Kodama, 1985). The depolarization-induced release of a number of putative neurotransmitters including ATP and its derivatives from neural preparations is well documented and the release of ATP from synaptosomes is atleast partly calcium-dependent (Israel et al., 1976).
In numerous tissues, the activities of Na+/K+-ATPase may be influenced by different endogenous modulators (Rodrigez and Pena, 1995; Balzan et al., 2000; Ewart and Klip, 1995). This enzyme may be under the influence of various exogenous factors including certain divalent metals and organic compounds of toxicological interest (Horvat et al., 1997; Nikezićet al., 1998; Vasćet al., 1999; Vasićet al., 2002) as well as some drugs (Modi and Merchant, 2003).
Magnesium (Mg2+)-ATPase is the major ATPase (in quantity) of the plasma membrane (Vajreswari and Narayana Reddy 1992). Mg2+-ATPase is sensitive to membrane fatty acid composition (Zimmerman and Daleke, 1993), which is related to the activity of intracellular calcium-stimulated fatty acid synthases, suggesting a possible synergistic relationship between Mg2+-ATPase and Ca2+-ATPase.
In vitro effects of aspirin and salicylate on erythrocyte diameter and Na+/K+ ATPase activity were studied. The effects of the two compounds appear to be mediated by their differential effects on the Na+/K+ ATPase activity (Sarkar et al., 1989; Modi and Merchant, 2003). The role of Na+/K+-ATPase in morphine-Induced antinociception was examined by (Masocha et al., 2003). Na+/K+-ATPase (á-subunit) was found to be down-regulated in rat brain synaptosomes under chronic morphine treatment (Prokai et al., 2005).
Despite the logical assumption that the actions of analgesic drugs would be same in the presence or absence of pain, the biochemical actions on different areas of the brain during experimental induction of pain remain relatively unexplored and very little attention has been paid on the analgesic effects in animal models without experimental induction of pain. Research in this line is necessary as the consumption of analgesic drugs is seen even when there are no actual signs or feel for pain. The present study is expected to contribute to the existing information on the involvement of analgesia induced by the administration of an opioid drug during pain or no-pain conditions.
The present study has been taken up to examine the changes in Na+/K+-, Mg2+- and Ca2+-ATPase enzyme activities using healthy rats and without induction of pain. The present study intends to examine the change in activity of ATPases with non-induction of pain to the animal. Not to go into the detailed activity during pain and the changes in their activity levels following tramadol administration, this research is supposed to throw light on what happens when an analgesic drug is administered without reference to its symptomatic effect.
MATERIALS AND METHODS
Procurement and maintenance of experimental animals: The study was conducted in the Department of Zoology, S.V. University and the Department of Life Sciences, University of Hyderabad, with collaborators from S.P. Mahila University, India, during the early part of the year 2006. Male adult Wistar rats weighing 150±20 g were used as the experimental animals in the present investigation. The rats were purchased from the Indian Institute of Science, Bangalore, India, maintained in the animal house of the Department in polypropylene cages under laboratory conditions of 28±2°C temperature, LD 12:12 photoperiod and 75% relative humidity. The rats were fed with standard pellet diet and water ad libitum. The rats were maintained according to the ethical guidelines for animal protection and welfare bearing the CPCSEA 438/01/a/cpcsea/dt 17.07.2001 in its resolution No: 9/IAEC/svu/2001/dt 04.03.2002.
Selection of drug and dosage for administration: Tramadol, an opioid analgesic drug was selected for the present study. It was obtained as a commercial grade chemical from Apollo Pharmacy, Hyderabad, India. After the rats were acclimated to the laboratory conditions, they were divided into groups depending on the dosage and time for sacrificing the animals. Five groups of six each were housed in separate cages. The time periods chosen for experiments following administration of the drug were 3, 6, 12 and 24 h.
Tramadol dosage was administered according to ED50 value obtained in rats at 31 mg kg-1 in hot plate test conducted by (Giusti et al., 1997). All doses were given in the morning between 9 and 10 AM, keeping in view the altered activity of rats during the nights compared to the daytime. Controls were maintained individually for each group, p>0.05.
Isolation of tissues: The present study was carried out on different areas of the brain, viz. cerebral cortex (CC), cerebellum (CB), pons-medulla (PM), hippocampus (HI) and hypothalamus (HY). The animals were sacrificed at the chosen time periods mentioned above. The brain was isolated immediately and placed on a chilled glass plate. The brain areas were separated and frozen in liquid nitrogen (-180°C) and stored at -80°C until further use. At the time of analyses the tissues were thawed and used. Isolation of mitochondria was done using the methods of Cotman and Matthews (1971) and Dodd et al. (1981).
Assay of ATPase (ATP phosphohydrolase: E.C.3.6.1.3) activity: ATPase activity was estimated in mitochondrial fraction in different rat brain areas.
(A) Mg2+ and Na+, K+ ATPase activities were assayed by the method of Fritz and Hamrick (1966) as reported by Desaiah and Ho (1979) with slight modifications.
The reaction mixture in a volume of 3.0 mL contained 3 mM ATP, 3 mM MgCl2, 100 mM NaCl2, 20 mM KCl, 135 mM imidazole-hydrochloric acid buffer (pH 7.5) and 0.3ml of mitochondrial suspension as enzyme source. The reaction mixture was incubated at 37°C for 30 min and stopped by the addition of 0.1 mL of 50% TCA. Samples were then assayed for inorganic phosphate using the method of Lowry and Lopez (1946) as modified by Phillips and Hayes (1977). The color was read at 800 nm in a spectrophotometer. Mg2+ ATPase activity was measured in the presence of 1 mM ouabain, a specific inhibitor of Na+/K+ ATPase (McIlwain, 1963). Ouabain sensitive Na+/K+ ATPase activity was obtained by the difference between total ATPase activity and Mg2+ ATPase activity. The enzyme activity was expressed as μmoles of inorganic phosphate formed mg-1 protein h-1.
Ca2+ ATPase activity was determined by measuring the inorganic phosphate liberated during the hydrolysis of ATP. Ca2+ ATPase was estimated in the mitochondrial fraction. The reaction mixture in a volume of 3.0 mL contained 135 mM imidazole-hydrochloric acid buffer (pH 7.5), 5 mM MgCl2, 0.05 mM CaCl2, 4 mM ATP and 0.3 mL of mitochondrial suspension as enzyme source. The reaction mixture was incubated at 37°C for 30 min and stopped by the addition of 0.1 mL of 50% TCA. The inorganic phosphate formed was estimated by the using the method of Lowry and Lopez (1946) as modified by Phillips and Hayes (1977). The color was read at 800 nm in a spectrophotometer. Mg2+ ATPase activity was measured in the presence of 0.5 mM EGTA and this value was subtracted from total ATPase activity to get Ca2+ ATPase activity. Enzyme activity was expressed as μmoles of inorganic phosphate formed mg-1 protein h-1.
Statistical treatment of data: The enzyme assay was carried out with six separate replicates from each group. The mean and Standard Deviation (SD) were worked out using INSTAT statistical software and Analysis of Variance (ANOVA) was done using SPSS statistical software using Basic Programming techniques on IBM compatible personal computer for different parameters. Difference between control and experimental assays was considered as not significant above p<0.05.
RESULTS
The levels of parameters related to energy metabolism, viz. activity of the enzymes (Na+/K+-ATPase, Mg2+-ATPase and Ca2+-ATPase), were estimated in different regions of the brain with reference to injection of single dose of tramadol and following the changes in the parameters during the 24 h period that followed. The results are presented in Table 1-3.
| Table 1: | Changes in the Sodium, Potassium-ATPase (Na+/K+-ATPase) activity levels in different brain regions of rats at different time periods after administration of a single effective dose of tramadol |
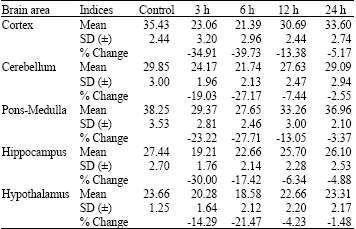 | |
| Each value, expressed in μmoles of inorganic phosphate formed mg-1 protein h-1, is the mean±Standard Deviation (SD) of observations from six separate experiments, Values are significant at least at p<0.05 in SNK test | |
| Table 2: | Changes in the Magnesium-ATPase (Mg2+-ATPase) activity levels in different brain regions of rats at different time periods after administration of a single effective dose of tramadol |
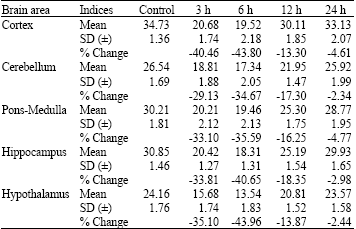 | |
| Each value, expressed in μmoles of inorganic phosphate formed mg-1 protein h-1, is the mean±Standard Deviation (SD) of observations from six separate experiments, Values are significant at least at p<0.05 in SNK test | |
| Table 3: | Changes in the Calcium-ATPase (Ca2+-ATPase) activity levels in different brain regions of rats at different time periods after administration of a single effective dose of tramadol |
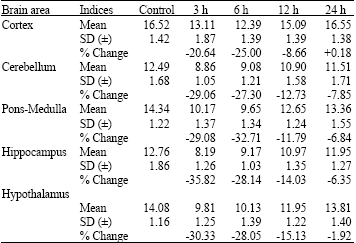 | |
| Each value, expressed in μmoles of inorganic phosphate formed mg-1 protein h-1, is the mean±Standard Deviation (SD) of observations from six separate experiments, Values are significant at least at p<0.05 in SNK test | |
Sodium and potassium ATPase: In the brain of control rats, PM registered the highest Na+/K+-ATPase enzyme activity and HY recorded the lowest level. Upon administration of single dose of tramadol, the Na+/K+-ATPase activity showed a decrease in all areas of the brain. Maximal decrease was in CC at 6 h, followed by HI at 3 h and PM CB and HY at 6 h.
Magnesium ATPase: The Mg2+-ATPase activity in the brain of control rats was found to be the highest in CC and lowest in HY.
Consequent on the administration of single dose of tramadol, the Mg2+-ATPase activity was found to decrease in all areas of the brain. Maximal decrease was in HY and CC, followed by HI, PM and CB. The maximal decreases in all areas were noticed at 6 h following tramadol administration.
Calcium ATPase: Ca2+-ATPase activity was found to be highest in CC in the brain of control rats. The enzyme activity was lowest in CB. Upon the administration of single dose of tramadol, the Ca2+-ATPase activity showed a decrease in all areas of the brain, with maximal decrease in HI at 3 h. CC showed a maximal decrease at 6 h following tramadol injection. As in the case of activities of Na+/K+ and Mg2+-ATPases, the Ca2+-ATPase activity showed recovery towards the control levels during the subsequent periods succeeding the periods of maximum activity. By 24 h after tramadol injection, the enzyme activity returned to near control levels and the minor deviations from the respective controls at this period were negligible and not statistically significant.
DISCUSSION
The present study was intended to examine the change in activity levels of the ATPases following administration of the opioid tramadol. In this study the changes in the enzyme activities connected with energy metabolism and membrane transport functions, viz. Na+/K+, Mg2+ and Ca2+ ATPase activities, were examined. The changes were recorded at different time periods following the injection of a single effective dose of tramadol during non-induction of pain.
The results obtained on ATPase activities indicate that the administration of tramadol without induction of pain would induce decrease in the activities of the ATPase enzymes towards lowered energy metabolism and alterations in membrane transport functions.
From inhibition of commercially available purified Na/K-ATPase isolated from porcine cerebral cortex in the presence of chosen drugs, as well as from the kinetic studies on synaptosomal plasma membranes, it was concluded that the drugs inhibit enzyme activity by acting in part directly on the enzyme protein (Horvat et al., 2006). The Na+/K+ ATPase activity of cells pre-treated with aspirin was lower than in controls (Modi and Merchant, 2003).
It has been reported that opioid drugs (Samsonova et al., 1979; Rekhtman et al., 1980; Brasc, 1990) may exert opposite effects on Na/K-ATPase activity, while some beta-blocking agents and cardiac glycosides inhibit the enzyme activity (Whikehart et al., 1991; Pelin, 1998; Clausen, 1998; Quadri and Ferrandi, 1998) in various tissues. The effect of drugs from the class of cardiac (methyldigoxin, verapamil, propranolol), sedative (diazepam) and antihistaminic (promethazine) drugs on Na/K-ATPase activity of plasma membranes in rat brain synaptosomes was studied (Horvat et al., 2006).
Several important synaptic plasma membrane proteins displayed significant regulation changes as a result of chronic morphine exposure in vivo. In particular, Na+/K+ ATPase, which is involved in regulation of the cell membrane potential by controlling sodium and potassium ion permeability was down-regulated in the neurons of morphine-exposed guinea-pigs (Prokai et al., 2005).
Further, as sodium and potassium ions are also important like calcium ions for the development and conduction of action potentials, the decrease in activities of Na+/K+ and Mg2+ ATPases may alter the rate of influx and efflux of the cations, namely Na+ and K+ respectively. Thus, the inhibition in the activity of Na+/K+-ATPase in specific regions of rat brain upon tramadol administration may be correlated with the altered membrane permeability properties. In the presence of the local anesthetic bupivacaine, Ca2+ concentrations remained significantly larger 60 sec after the initial peak. This also points to an inhibition of Ca2+ reuptake (Zink et al., 2002).
These reports, based results on a single-dose administration of tramadol on different brain areas, but not with other opioid drugs, make them relevant to the present observations of the present study as the administration of analgesics during the absence of pain or non-induction of pain would presumably down-regulate the activity of Ca2+ ATPase. The decrement in the activities of Na+/K+-, Mg2+- and Ca2+- dependent ATPases reflects the decreased turnover of ATP, presumably due to inhibition of the oxidoreductase system and uncoupling of the same from the electron transport system. These indicate that the administration of tramadol would induce decreases in the activities of the ATPase enzymes related to energy metabolism and membrane transport functions It was apparent that as the effects of the drug on different parameters get accentuated by 3 or 6 h after its administration. Following this, as the effect of the drug started to gradually vane. In the absence of pain, what an analgesic will do is a matter of curiosity. The work of Chetan (2006) indicated that even during non-induction or absence of pain-administration of tramadol causes changes in the activity levels of ATPases.
Thus, injection of a single dose of tramadol causes perceptible changes in the energy metabolism and the associated metabolic profiles, at least as part of its antinociceptive effect. These changes occur even during non-induction of pain and are indicative of some of the contrivances by which tramadol could normally exercise its analgesic effects in the presence of pain or during pre-emptive analgesia.
Despite the enormous work done in the line of the actions of analgesic drugs, the present research, we believe, is the first to be done in this area of pharmacology as the literature pertaining to this type of work is very scanty. The present investigation may look a wee bit nascent, but is expected to provide useful information on the biochemical changes with one-time administration of tramadol in different areas of brain during non-induction of pain or total absence of pain.
In conclusion, without reference to the pain-dependent models, the present study demonstrates that the activity levels of ATPases has discernible changes even during non-induction or total absence of pain in all the brain areas, presumably as a measure for regulating the level of the administered tramadol.
REFERENCES
- Balzan, S., G. D'Urso, S. Ghione, A. Martinelli and U. Montali, 2000. Selective inhibition of human erythrocyte Na+/K+ ATPase by cardiac glycoside and by mammalian digitalis like factor. Life Sci., 67: 1921-1928.
Direct Link - Banning, A., P. Sjogren and H. Henriksen, 1991. Pain causes in 200 patients referred to a multidisciplinary cancer pain. Clin. Pain, 45: 45-48.
Direct Link - Banning, A., P. Sjogren and H. Henriksen, 1991. Treatment outcome in a multidisciplinary cancer pain. Clin. Pain, 47: 129-134.
Direct Link - Brasc, D.A., 1990. Is intracellular sodium involved in the mechanism of tolerance to opioid drugs? Med. Hypotheses, 32: 161-167.
Direct Link - Clausen, T., 1998. Clinical and therapeutic significance of the Na+, K+ pump. Clin. Sci., 95: 3-17.
Direct Link - Dodd, P.R., J.A. Hardy, A.E. Oakley, J.A. Edwardson, E.K. Perry and J.P. Delaunoy, 1981. A rapid method for preparing synaptosomes: Comparison, with alternative procedures. Brain Res., 226: 107-118.
CrossRefDirect Link - Ewart, H.S. and A. Klip, 1995. Hormonal regulation of the Na+-K+-ATPase: Mechanisms underlying rapid and sustained changes in pump activity Am. J. Physiol.-Cell Physiol., 269: C295-C311.
Direct Link - Giusti, P., A. Buriani, L. Cima and M. Lipartiti, 1997. Effect of acute and chronic tramadol on [3H]-5-HT uptake in rat cortical synaptosomes. Br. J. Pharmacol., 122: 302-306.
Direct Link - Lowry, O.H. and J.A. Lopez, 1946. The determination of inorganic phosphate in the presence of labile phosphate esters. J. Biol. Chem., 162: 421-428.
PubMedDirect Link - Masocha, W., G. Horvath, A. Agil, M. Ocana and E. Del-Pozoet, 2003. Role of Na+,K+-ATPase in morphine-induced antinociception. J. Pharmacol. Exp. Ther., 306: 1122-1128.
Direct Link - Modi, D.N. and M.A. Merchant, 2003. In vitro effects of aspirin and salycilate on erythrocytes: Size and Na+/K+ ATPase activity. Indian J. Pharmacol., 35: 27-31.
Direct Link - Nikezi, G., A. Horvat, N. Nedeljkovi, S. Todorović and V. Nikolić et al., 1998. Influence of pyridine and urea on the rat brain ATPase activity. Gen. Physiol. Biophys., 17: 15-23.
Direct Link - Pasternak, G.W., 1993. Pharmacological mechanisms of opioid analgesics. Clin. Neuropharmacol., 16: 1-18.
Direct Link - Portenoy, R.K., K.M. Foley and C.E. Inturrisi, 1990. The nature of opioid responsiveness and its implications for neuropathic pain: New hypotheses derived from studies of opioid infusions. Pain, 43: 273-286.
Direct Link - Prokai, L., A.D. Zharikova and S.M. Jr. Stevens, 2005. Effect of chronic morphine exposure on the synaptic plasma-membrane sub proteome of rats: A quantitative protein profiling study based on isotope-coded affinity tags and liquid chromatography/mass spectrometry. J. Mass Spectrum, 40: 169-175.
Direct Link - Quadri, L. and M. Ferrandi, 1998. Involvement of Na+, K+-ATPase and its inhibitors in cardiovascular diseases. Exp. Opin. Ther. Pat., 8: 39-52.
Direct Link - Rodriguez de Lores Arnaiz, G. and C. Pena, 1995. Characterization of synaptosomal membrane Na+, K+-ATPase inhibitors. Neurochem. Int., 27: 319-327.
CrossRefDirect Link - Vajreswari, A. and K. Narayana Reddy, 1992. Effect of dietary fats on erythrocytes membrane lipid composition and membrane-bound enzyme activities. Metabolism, 41: 352-358.
Direct Link - Vasic, V., D. Jovanovic, D. Krstic, G. Nikezic, A. Horvat, L. Vujisic and N. Nedeljkovic, 1999. Prevention and recovery of CuSO4-induced inhibition of Na+/K+-ATPase and Mg2+-ATPase in rat brain synaptosomes by EDTA. Toxicol. Lett., 110: 95-103.
CrossRefDirect Link - Vasi, V., D. Jovanovi, A. Horvat, T. Momić and G. Nikeziće, 2002. Effect of Cd2+ and Hg2+ on the activity of Na+/K+-ATPase and Mg2+-ATPase adsorbed on polystyrene microtiter plates. Anal. Biochem., 300: 113-120.
Direct Link - Whikehart, D.R., B. Montgomery, D.H. Sorna and J.D. Wells, 1991. Beta-blocking agents inhibit Na+, K+-ATPase in cultured corneal endothelial and epithelial cells. J. Ocul. Pharmacol., 7: 195-200.
Direct Link - Zimmerman, M.L. and D.L. Daleke, 1993. Regulation of a candidate aminophospholipid-transporting ATPase by lipids. Biochemistry, 32: 12257-12263.
Direct Link - Zink, W., B.M. Graf, B. Sinner, E. Martin, R.H. Fink and G. Kunst,, 2002. Differential effects of bupivacaine on intracellular Ca2+ regulation-potential mechanisms of its myotoxicity. Anesthesiology, 97: 710-716.
Direct Link