
A.E. Samir Bashandy
Pharmacology Department, National Research Center, Egypt
International Journal of Pharmacology
Year: 2007 | Volume: 3 | Issue: 1 | Page No.: 27-33







ABSTRACT
High plasma level of cholesterol or triglycerides was associated with poor semen quality and direct adverse effects on testicular function that may lead to male infertility. The effect of fixed oil of Nigella sativa (N. sativa) on male fertility in normal and hyperlipidemic rats was studied. Induction of hyperlipidemia was done by feeding rats on a diet containing 1% cholesterol, 2% sheep fat and 0.5% cholic acid for 2 months. Oil of N. sativa was orally given (0.5mL/rat) for 2 months daily to normal and hyperlipidemic rats. Plasma lipid profile including cholesterol, triglycerides, Low Density Lipoprotein (LDL) and High Density Lipoprotein (HDL) was evaluated. Fertility index as determined by sex organ weights, plasma testosterone level and sperms evaluation (sperm motility, sperm abnormalities and sperm count) was investigated. In hyperlipidemic rats (control +ve), there was a marked increase in plasma cholesterol, triglycerides and LDL levels. On the other hand, there was a significant decrease in HDL and fertility index associated with increase in sperm abnormalities. Oral administration of oil of N. sativa to either normal or hyperlipidemic rats improved the fertility index as manifested by increase in seminal vesicle weight, testosterone level, sperm motility and sperm count and a decrease in sperm abnormalities. In conclusion, administration of N. sativa oil to hyperlipidemic rats improved their reproductive efficiency and produced additional protection against hyperlipidemia induced reduction in fertility.
PDF Abstract XML References
How to cite this article
A.E. Samir Bashandy, 2007. Effect of Fixed Oil of Nigella Sativa on Male Fertility in Normal and Hyperlipidemic Rats. International Journal of Pharmacology, 3: 27-33.
DOI: 10.3923/ijp.2007.27.33
URL: https://scialert.net/abstract/?doi=ijp.2007.27.33
DOI: 10.3923/ijp.2007.27.33
URL: https://scialert.net/abstract/?doi=ijp.2007.27.33
INTRODUCTION
Damage induced by endogenously generated reactive oxygen species has been proposed to be a major contributing factor in a variety of human diseases, including male infertility (Gagnon et al., 1991). Hypercholesterolemia was found to cause an increase in oxygen radicals production and lipid peroxidation levels in different tissues (Ohara et al., 1993). Lipid peroxidation is an important factor that may induce morphological changes in the spermatozoa (Sanchez et al., 2006). Feeding rats with diet supplemented with fat leads to increase in testes cholesterol level and degeneration of some gonadal cells (Chertok et al., 2001). The feeding of male rabbits on a diet rich in fat results in a functional disorder of hypothalamo-pituitary gonadal axis associated with short penis and damage of spermatogenesis (Zhu et al., 2005). It was found that the decrease in HDL and elevation of total cholesterol resulted in erectile dysfunction in men (Rao et al., 2006).
Nigella stativa (N. sativa) commonly known as black seed, belongs to family Ranunculaceae. It has been use in many Middle Eastern countries as a natural remedy (Swamy and Tan, 2000). Much of biological activity of the seeds has been shown to be due to thymoquinone, the major component of the essential oil, but it is also present in the fixed oil. The seeds or its oil have analgesic, antipyretic, antineoplastic and antihypertensive activities (Ali and Blunden, 2003). The fixed oil of N. sativa oil consists of 50% linoleic acid, 25% oleic acid, 12% palmitic acid, 2.84% stearic acid, 0.34% linolenic acid and 0.35% myristic acid (Cheikh-Rouhou et al., 2007). The fixed oil produced an increase in glutathione level of stomach (El-Dakhakhny et al., 2000a) and a decrease of serum total cholesterol in rats (El-Dakhakhny et al., 2000b). N. sativa oil has been reported to possess antioxidant activity (Bruits and Bucar, 2000). Thymoquinon has been shown to protect mice against benzo-pyrene induced chromosomal aberrations (Badary et al., 2007).
Supplementation of animal diets with oils rich in polyunsaturated fatty acids, such as linoleic acid has positively influenced reproductive functions (Fellner et al., 1995). Since N. sativa oil contains high percentage of unsaturated fatty acids and there are no reports concerning the effect of this oil on male reproductive system or sexual hormones, our objectives are investigation the effects of N. sativa oil on testicular endocrine, sperm quality and sperm count in normal and hyperlipidemic male rats.
MATERIALS AND METHODS
Materials: Sixty Four male (120-150 g, 3 months age) albino, Swiss strain rats were used in this study. The animals were purchased from the Ministry of public health, Helwan, Egypt. Cholesterol and cholic acid were obtained from El-Gomhorya Co., Egypt as pure powder. N. sativa oil was purchased from El-Kahira Co., Egypt.
Animals feeding: The rats were divided into two equal main groups, the first group was fed a basal diet throughout the experimental period and it is called normal group. The second main group was fed a basal diet supplemented with 1% cholesterol, 0.5% cholic acid (Balkan et al., 2004) and 2% sheep fat for 2 months and it is called hyperlipidemic group. Both the two main groups were subdivided into two equal subgroups, control and treated with N. sativa oil, each contains 16 rats. There were two types of control, contro –ve fed on the basal diet and control +ve Fed on the basal diet containing cholesterol. The animals were allowed free access to food and water. At 2 months postfeeding and before oil administration, the animals were fasted overnight and blood samples were collected from orbital plexus of male rats. Plasma cholesterol, triglycerides, LDL and HDL levels were measured colourimetrically with Kits (Bio Merieux, France) and these levels were considered the values of zero time.
Oil Administration and experimental: The oil of N. sativa was administered orally (0.5mL/rat) to both normal and hyperlipidemic rats daily for 2 months. Feeding of male rats on experimental diets extended throughout the experimental period.
Blood samples were collected from 8 animals in each group monthly (1 and 2 months post oil administration) and plasma was separated. Plasma cholesterol, triglycerides, LDL, HDL and testosterone levels were evaluated using kits from Bio Merieux, France. Testosterone level was assayed by enzyme immunoassay and other parameters were determined by using spectrophotometer. At the end of each month, 8 animals from each group were sacrificed, then testes, epididymis and seminal vesicles were removed and weighted. The epididymis was dissected in 20mL of normal saline (0.9% NaCl); incubated at 37°C and sperm motility was evaluated. Sperms were counted using a hemocytometer. Smears were prepared from the suspension stained with 1% eosin solution and examined for sperm abnormalities (Filler, 1993). Three hundreds spermatozoa per animal were evaluated for head and tail defects by light microscope. Classification of individual spermatozoa were (1) normal, (2) head abnormalities and (3) tail abnormalities. The percentage of total sperm abnormalities was calculated. The head abnormalities include small head,amorphous head,no hook and two heads. The tail abnormalities include folded tail, short tail and two tails.
The feeding of the rats and collection of blood samples were carried out in the animal house, National Resarch Center, Egypt. All the biochemical analysis and sperms evaluation were performed in Pharmacology Dept. NRC, Egypt.
Statistical Analysis: Statistical difference was calculated by using one Way analysis of variance and least significant difference range test (LSD). Data were expressed as means±SE.
RESULTS
Lipid profile: Feeding of male rats a diet containing 1% Cholesterol significantly (p<0.01) increased total cholesterol, triglycerides (Table 1) and LDL (Table 2) levels in plasma. On the other hand, HDL level (Table 2) decreased significantly (p<0.01) as compared to those fed the basal diet.
Oral administration of oil of Nigella sativa to normal rats caused a significant decrease (p<0.01) in Cholesterol, triglycerides and LDL levels as compared to control -ve, 2 months post administration and a significant increase (p<0.01) in HDL level.
The treatment of hyperlipidemic rats with the oil led to a significant decrease (p<0.01) of previous parameters except HDL which elevated significantly at 1 and 2 months post treatment as compared to control +ve
Weights of sex organs: The weights of testes and seminal vesicles of hyperlipidemic control rats decreased significantly (p<0.01) 2 months post treatment (Table 3) as compared to normal control.
Normal rats given oil of N. sativa showed a significant (p<0.01) increase in seminal vesicle weight at all time intervals, while the weights of testes and epididymis didn't change significantly. Hyperlipidemic rats treated with N. sativa oil showed increase in seminal vesicles (p<0.01) weight at all time intervals and in testes weight at 2 months post treatment as compared to control +ve.
Testosterone Level: Plasma testoterone level (Fig. 1) of hyperlipidemic control rats (control +ve) exhibited a significant decrease (p<0.01) at all time intervals as compared to normal control.
The hormone level elevated significantly (p<0.01) in normal or hyperlipidemic rats treated with oil of N. sativa as compared to corresponding control at any time interval.
Sperm Quality and Count:
Sperm motility: The values of sperm motility of control +ve are significantly (p<0.01) less than those of control –ve (Fig. 2).
| Table 1: | Effect of Nigella sativa oil on plasma cholesterol and triglycerides levels (mg/100 mL) in normal and hyperlipidemic rats |
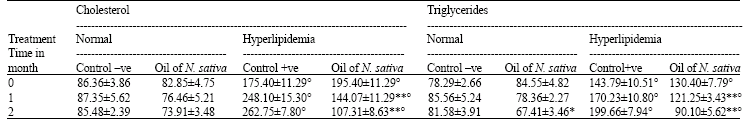 | |
| n = 8 per group, **p<0.01, *p<0.05 as compared with corresponding control, °p<0.01 as compared with corresponding normal group | |
| Table 2: | Effect of Nigella sativa oil on plasma LDL and HDL concentrations (mg/100 mL) in normal and hyperlipidemic rats |
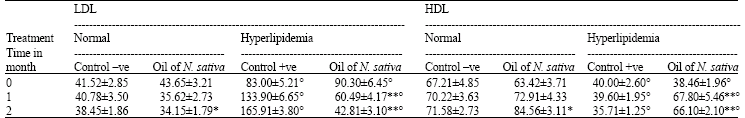 | |
| n = 8 rats per group, **p<0.01, *p<0.05 as compared with corresponding control, °p<0.01 as compared with corresponding normal group | |
| Table 3: | Effect of Nigella sativa oil on weights of some sex organs (g.) relative to body weight of normal and hyperlipidemic male rats |
 | |
| n = 8 rats per group **p<0.01 as compared with corresponding control. °p<0.01 as compared with corresponding normal control | |
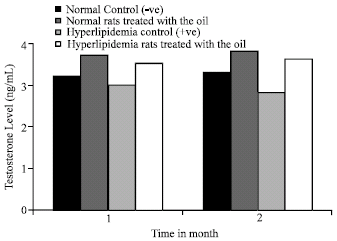 | |
| Fig. 1: | Effect of N. sativa oil on plasma testosterone level of normal and hyperlipidemic rats |
The motility of sperm elevated significantly (p<0.05) in normal rats given N. sativa oil after 2 months. Similar results were recorded in case of hyperlipidemic rats treated with oil of N. sativa at 1 (p<0.05) and 2 (p<0.01) months post-treatment as compared to +ve control.
Total sperm abnormality: Sperm analysis revealed a significant increase (p<0.01) in sperm abnormalities in +ve control at all time intervals (Fig. 3) as compared to –ve control.
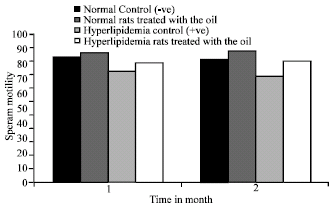 | |
| Fig. 2: | Effect of Nigella sativa oil on sperm motility (%) of normal and hyperlipidemic rats |
The treatment at normal or hyperlipidemic rats with N. sativa oil significantly (p<0.01) decreased sperm abnormalities at all time intervals.
Sperm count per epididymis: The results indicates that the sperm count of hyperlipidemic control rats Fig. 4 decreased Significantlly(p<0.01) 2 Months post treatment as compared to normal control.
Normal or hyperlipidemic rats treated with oil of N. sativa showed a significant increase (p<0.01) in sperm count as compared to their controls.
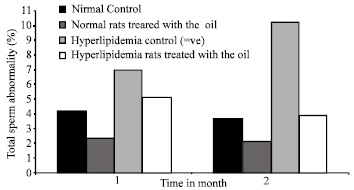 | |
| Fig. 3: | Effect of Nigella sativa oil on total sperm abnormality (%) of normal and hyperlipidemic rats |
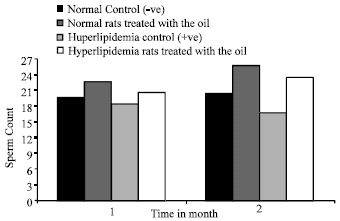 | |
| Fig. 4: | Effect of Nigella sativa oil on sperm count per epididymis (x 107) of normal and hyperlipidemic rats |
DISCUSSION
The cholesterol enriched diets is regarded as important factor in the development of abnormalities of male reproductive system (Shalaby et al., 2004), since it leads to the development of hyperlipidemia, atherosclerois and abnormal lipid metabolism.
In the present study, feeding of a diet containing cholesterol to male rats increased plasma total cholesterol, triglycerides and LDL, while it results in a decrease of HDL. Elevated triglycerides are now considered an independent risk factor for coronary heart disease and acute pancreatitis (Bersot et al., 2006). The anti atherogenic effect of HDL has been mainly attributed to its role in the reverse cholesterol transport (Fielding and Fielding, 1995).
Our results showed a significant decrease of plasma testosterone level in hyperlipidemic rats. This decrease may result from the increased number of degenerating Leydig cells (Purohit and Daradka, 1999) reduction of Leydig's cell nuclear diameter (Gupta and Dixi, 1988) or from decreased Leutinizing hormone level (tanaka et al., 2001) and testicular activity of 17 beta hydroxysteroid dehydrogenase (Gromadzka-Ostrowska et al., 2002). Moreover, serum testosterone response to testicular stimulating with human chorionic gonadotropin was significantly lower in hypercholesterolemic rabbits (Yamamoto et al., 1999).
The reproductive efficiency of hypercholesterolemic control rats of this study was low when compared to normal rats. This effect characterized by a significant decrease in fertility index, seminal vesicle weight, plasma testosterone level, sperm motility and sperm count associated with an increase in total sperm abnormalities. These effects may be due to direct effect of hyperlipidemia which result in a disorder of hypothalamo pituitary gonadal axis and damage of spermatogenesis (Zhu et al., 2005) or due to increase of oxidative stress, since both hypercholesterolemia (Ohara et al., 1993) and hypertriglyceridemia (Bae et al., 2001) caused an increase of oxygen radicals production and lipid peroxidation level in different tissues associated with decreased antioxidant effect of glutathione (De La Cruz et al., 2000). Reactive Oxygen Species (ROS) release was positively correleated with LDL concentration of hyperlipidemic patients and negatively correlated with HDL level leading to activation of oxidative stress (Araujo et al., 1995). Oxidative Stress (OS) arises as a consequence of excessive production of ROS and impaired antioxidant defense mechanism (Sikka, 2001). The generation of ROS has become a real concern because their potential toxic effects on sperm quality and function (Sikka, 1996). Spermatozoa are particularly susceptible to OS induced damage because their plasma membranes contain large quantities of fatty acids (Alvarez and Storey, 1995). OS mediated damage in the sperm plasma membranes may account for defective sperm function (Iwasaki and Gagnon, 1992). ROS induced lipid peroxidation is a causativa agent of morphological changes in sperms (Sanchez et al., 2006). OS induced DNA damage may accelerate the process of germ cell apoptosis (Agarwal et al., 2003) leading to a decline in sperm counts. It was reported that the significant reduction in sperm concentration and percentage of motile spermatozoa in rabbits with hypercholesterolemia could be attributed to defects in the secretory function of the Sertoli and Leydig cells resulting in impaired spermatogenesis and epididymal sperm maturation process. Moreover, epididymal dysfunction in hyper-cholesterolemic animals may result in both decreased sperm motility and increased sperm abnormalities (Yamamoto et al., 1999). It was Concluded that hyperlipidemia induced by cholesterol feeding to male rats altered testicular histology. The sperm motility and density were significantly reduced in cauda epididymis and testes of hyperlipidemic rats. The testicular cell population i.e. spermatocytes and spermatids were significantly reduced (Purohit and Daradka, 1999). The increased formation of ROS has been correlated with a reduction in sperm motility (Armstrong et al., 1999).
Administration of N. stativa oil to both normal or hyperlipidemic rats in the current study caused a significant decrease in cholesterol, triglycerides and LDL Concentrations and a significant increase in HDL Level. The reduction in lipid concentrations by N. sativa oil may result from the hypolipidemic effects of oleic (Allman-Farinelli et al., 2005) and linoleic acids (Wendel and Belury, 2006) major unsaturated fatty acids of the oil or from its effect on lipoprotein.
The testosterone level of normal and hyperlipidemic rats given oil of N. sativa orally elevated significantly. The dietary fatty acids affect the androgen metabolism. The highest plasma level of androgen was observed in male rats fed diets rich in unsaturated fatty acids, while lowest level was observed in the animals fed diets rich in saturated Fatty acids. The unsaturated fatty acids stimulate the activity of 17 β-hydoxysteroid dehydrogenase, the most important key enzyme in the testosterone synthesis pathway (Gromadzka-Ostrowska et al., 2002). The observed increase in seminal vesicles weight could be attributed to the increase in testosterone level, since the growth of seminal vesicles is highly dependent on androgen.
N. sativa oil is an effective free radical scavenger showing antioxidant activity and protection against the damage caused by free radicals. Therefore the oil is useful in hyperlipidemia induced free radicals (Badary et al., 2000). N. sativa oil protects some tissues from oxidative stress and lipid peroxidation (Ozugurlu et al., 2005). In the present study, oil or N. sativa administration led to increase in HDL level which act as antioxidant (Idzior and Walus et al., 2004). The present results revealed that oral administration of N. sativa oil to hypercholesterolemic rats significantly improved their reproductive efficiency; seminal vesicle weight, testosterone level, sperm motility and sperm quality. The antioxidant and hypolipidemic effects of N. sativa oil may be responsible for its beneficial action on improved the fertility index in hyperlipidemic rats. N. sativa oil contains unsaturated fatty acids which are effective for reproduction. It was concluded that the dietary unsaturated fatty acids are valuable to sustain the reproductive capacity of male turkeys especially when they are getting older (Blesbois et al., 2004). Polyunsaturated Fatty acids prevent the decrease in sperm output that normally occurs by 60 weeks of age in broiler chicken (Surai et al., 2000). Unsaturated fatty acids are important in sperm maturation. Dietary supplementation with shark liver oil containing unsaturated fatty acids significantly increased the percent motility of sperms and might improve the reproduction (Mitre et al., 2004).
It could be concluded that administration of N. sativa oil to hypercholesterolemic male rats improves the reduced male fertility induced by hypercholesterolemia.
REFERENCES
- Ali, B.H. and G. Blunden, 2003. Pharmacological and toxicological properties of Nigella sativa. Phytother. Res., 17: 299-305.
CrossRefPubMedDirect Link - Allman-Farinelli, M.A., K. Gomes, E.J. Favoloro and P. Petocz, 2005. A Diet Rich in high oleic-acid sunflower oil favorably alters low density lipoprotein cholesterol, triglycerides and factor VII coagulant activity. J. Am. Dietetic Assoc., 105: 1071-1079.
Direct Link - Alvarez, J.G. and B.T. Storey, 1995. Differential incorporation of fatty acids into and peroxidative loss of fatty acids from phospholipids of human spermatozoa. Mol. Reprod. Dev., 42: 334-346.
PubMedDirect Link - Armstrong, J.S., M. Rajasekaran, W. Chamulitra, P. Gatti, W.J. Hellstrom and S.C. Sikka, 1999. Characterization of reactive oxygen species induced effects on human spermatozoa movement and energy metabolism. Free Radic. Biol. Med., 26: 869-880.
CrossRefDirect Link - Badary, O.A., M.F. Abd-Ellah, M.A. El-Mahdy, S.A. Salama and F.A. Hamada, 2007. Anticlastogenic activity of Thymoquinone against benzo(a) pyrene in mice. Food Chem. Toxicol., 45: 88-92.
CrossRef - Bae, J., E. Bassenge, K. Kim, Y. Kim and K.S. Kim et al., 2001. Postprandial hypertriglyceridemia impairs endothelial function by enhanced oxidant stress. Atherosclerosis, 155: 517-523.
Direct Link - Balkan, J., S. Dogru-Abbasoglu, G. Aykac-Toker and M. Uysal, 2004. The effect of a high cholesterol diet on lipids and oxidative stress in plasma, liver and aorta of rabbits and rats. Nutr. Res., 24: 229-234.
Direct Link - Bersot, T., S. Haffner, W.S. Harris, K.A. Kellick and C.M. Morris, 2006. Hypertriglyceridemia: Management of atherogenic dyslipidemia. J. Fam. Pract., 55: 1-8.
Direct Link - Blesbois, E., V. Douward, M. Germain, P. Boniface and F. Pellet, 2004. Effects of n-3 polyunsaturated dietary supplementation on the reproductive capacity of male turkeys. Theriogenology, 61: 537-549.
Direct Link - Cheikh-Rouhou, S., S. Besbes, B. Hentati, C. Blecker, C. Deroanne and H. Attia, 2007. Nigella sativa L.: Chemical composition and physicochemical characteristics of lipid fraction. Food Chem., 101: 673-681.
CrossRefDirect Link - Chertok, V.M., T.A. Botvich, M.A. Khasina and O.A. Artyukova, 2001. Effect of Pollack liver oil on lipid metabolism in rat testes.Effect of Pollack liver oil on lipid metabolism in rat testes. Bullet Experim. Biol. Med., 131: 116-118.
Direct Link - De La Cruz, J.P., L. Quintro, M.A. Villalobos and F.S. De La Cuesta, 2000. Lipid peroxidation and glutathione system in hyperlipemic rabbits: Influence of olive oil administration. Biochem. Biophys. Acta, 1485: 36-44.
Direct Link - El-Dakhakhny, M., N.I. Mady and M.A. Halim, 2000. Nigella sativa L. oil protects against induced hepatotoxicity and improves serum lipid profile in rats. Arzneimittelforschung, 50: 832-836.
CrossRefPubMedDirect Link - Fellner, S.J., F.D. Sauer and J.G.K. Kramer, 1995. Steady-state rates of linoleic acid biohydrogenation by ruminal bacteria in continuous culture. J. Dairy Sci., 78: 1815-1815.
Direct Link - Fielding, C.J. and P.E. Fielding, 1995. Molecular physicology of reverse cholesterol transport. J. Lipid Res., 36: 211-228.
Direct Link - Gromadzka-Ostrowska, J., M. Przepiorka and K. Romanowicz, 2002. Influence of dietary fatty acids composition, level of dietary fat and feeding period in some parameters of androgen metabolism in male rats. Reprod. Biol., 2: 277-293.
Direct Link - Iwasaki, A. and C. Gagnon, 1992. Formation of reactive oxygen species in spermatozoa of infertile patients. Fertil. Steril., 57: 409-416.
PubMedDirect Link - Mitre, R., C. Cheminade, P. Allaume, P. Legrand and A.B. Legrand, 2004. Oral intake of shark liver oil modifies lipid composition and improves motility and velocity or boar sperm. Theriogenology, 62: 1557-1566.
Direct Link - Ozugurlu, F., S. Sahin, N. Idiz, O. Akyol, A. Iihan, R. Yigitoglu and B. Isik, 2005. The effect of Nigella sativa oil against experimental allergic encephalomyelitis via nitric oxide and other oxidative parameters. Cell. Mol. Biol., 51: 337-342.
Direct Link - Purohit, A. and H.M. Daradka, 1999. Effect of mild hyperlipidemia on testicular cell population dynamics in albino rats. Ind. J. Exp. Biol., 37: 396-398.
Direct Link - Sanchez, E.E.T., M.L. Marquette, D.B. Brown and N.H. Ansari, 2006. The effect of oxidative stress on human sperm morphology. Fertil., Steril., 86: S444-S444.
Direct Link - Shalaby, M.A., H.Y. El-Zorba and G.M. Kamel, 2004. Effect of alpha-tocopherol and simvastation on male fertility in hypercholesterolemic rats. Pharmacol. Res., 50: 137-142.
Direct Link - Sikka, S.C., 1996. Oxidative stress and role of antioxidants in normal and abnormal sperm function. Frontiers Biosci., 1: 78-86.
PubMed - Sikka, S.C., 2001. Relative impact of oxidative stress on male reproductive function. Curr. Med. Chem., 8: 851-862.
Direct Link - Surai, P.F., R.C. Noble, N.H. Sparks and B.K. Speake, 2000. Effect of long term supplementation with arachidonic or docosahexaenoic acids on sperm production in the broiler chicken. J. Reprod. Fertil., 120: 257-264.
Direct Link - Swamy, S.M.K. and B.K.H. Tan, 2000. Cytotoxic and immunopotentiating effects of ethanolic extract of Nigella sativa L. seeds. J. Ethnopharmacol., 70: 1-7.
CrossRefDirect Link - Tanaka, M., S. Nakaya, T. Kumai, M. Watanabe, N. Matsumoto and S. Kobayashi, 2001. Impaired testicular function in rats with diet induced hypercholesterolemia and/or streptozotocin-induced diabetes mellitus. Endocr. Res., 27: 109-117.
Direct Link - Wendel, A.A. and M.A. Belury, 2006. Effects of Conjugated linoleic acid and troglitazone on lipid accumulation and composition in lean and Zucker diabetic fatty rats. Lipids, 41: 241-247.
Direct Link - Yamamoto, Y., K. Shimamoto, N. Sofikitis and I. Miyagaula, 1999. Effects of hypercholesterolaemia on Leydig and Sertoli cell secretory function in the overall sperm fertilization capacity in the rabbit. Human Reprod., 14: 1516-1521.
Direct Link - Badary, O.A., A.B. Abdel-Naim, M.H. Abdel-Wahab and F.M. Hamada, 2000. The influence of thymoquinone on doxorubicin-induced hyperlipidemic nephropathy in rats. Toxicology, 143: 219-226.
CrossRefDirect Link