
Lijun Zhu
Department of Basic Medical Sciences, Zhejiang University School of Medicine, 353 Yan an Road, Hangzhou, Zhejiang, 31003 1, Peoples Republic of China Department of Medical Oncology and Therapeutic Research, City of Hope National Medical Center, Duarte, CA, 91010, USA
Yun Yen
Department of Basic Medical Sciences, Zhejiang University School of Medicine, 353 Yan an Road, Hangzhou, Zhejiang, 31003 1, Peoples Republic of China Department of Medical Oncology and Therapeutic Research, City of Hope National Medical Center, Duarte, CA, 91010, USA
Jimin Shao
Department of Basic Medical Sciences, Zhejiang University School of Medicine, 353 Yan an Road, Hangzhou, Zhejiang, 31003 1, Peoples Republic of China Department of Medical Oncology and Therapeutic Research, City of Hope National Medical Center, Duarte, CA, 91010, USA
Christina Qi
Department of Medical Oncology and Therapeutic Research, City of Hope National Medical Center, Duarte, CA, 91010, USA
Christina Yen
Department of Medical Oncology and Therapeutic Research, City of Hope National Medical Center, Duarte, CA, 91010, USA
Jianhong Luo
Department of Basic Medical Sciences, Zhejiang University School of Medicine, 353 Yanan Road, Hangzhou, Zhejiang, 31003 1, Peoples Republic of China
Bingsen Zhou
Department of Medical Oncology and Therapeutic Research, City of Hope National Medical Center, Duarte, CA, 91010, USA
International Journal of Pharmacology
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 324-330







ABSTRACT
Overexpression of Vascular Endothelial Growth Factor (VEGF) has been associated with increased angiogenesis, tumor growth and metastasis in tumors. Fibroblast Growth Factor Receptor 3 (FGFR3) acts as an oncogene in several tumor types, including multiple myeloma, bladder cancer and cervical cancer. Our previous studies have shown that inhibition of FGFR3 caused down regulation of VEGF in multiple myeloma. In order to better understand the molecular mechanism of FGFR3 and VEGF, L6 cells lacking any endogenous FGFR were stably transfected with FGFR3 at the present study. The FGFR3 expression was validated by Northern blot and Western blot. Compared with parental L6 cell and L6V (transfected with vector only), L6 cells with stably transfected FGFR3 showed overexpression of VEGF. [3H] Thymidine uptake and CellTiter 96 Aqueous One Solution cell proliferation assay demonstrated that DNA synthesis and cell proliferation were increased in FGFR3 expression L6 cells. These observations indicate that FGFR3 might be associated with regulation of VEGF, suggesting that combinations of inhibition of VEGF and inhibition of FGFR3 may have an advantage for the treatment of FGFR3 overexpression neoplastic disease.
PDF Abstract XML References Citation
How to cite this article
Lijun Zhu, Yun Yen, Jimin Shao, Christina Qi, Christina Yen, Jianhong Luo and Bingsen Zhou, 2006. Fibroblast Growth Factor Receptor 3 Up-regulates Vascular Endothelial Growth Factor Expression in L6 Cells. International Journal of Pharmacology, 2: 324-330.
DOI: 10.3923/ijp.2006.324.330
URL: https://scialert.net/abstract/?doi=ijp.2006.324.330
DOI: 10.3923/ijp.2006.324.330
URL: https://scialert.net/abstract/?doi=ijp.2006.324.330
INTRODUCTION
Fibroblast Growth Factor Receptor 3 (FGFR3) is a membrane spanning tyrosine kinase that serves as a high affinity receptor for Fibroblast Growth Factors (FGFs) (Martin, 1998). FGFR3 contains three glycosylated extracellular immunoglobulin-like (Ig-like) domains, a Transmembrane (TM) domain and a split intracellular tyrosine kinase domain. FGFR3 is highly expressed in the cartilage growth plates, skin and central nervous system in a developing mouse. In adult tissues, it is detected in the brain, kidneys, testes and resting cartilages. Under ligand stimulation, FGFR3 undergoes dimerization and tyrosine autophosphorylation, resulting in cell proliferation, differentiation and migration in many different tissues. Depending on the cell context, this is achieved through different signal transduction pathways (Kanai et al., 1997; Chest et al., 2001).
The t (4;14) (p16;q32) translocation occurs in approximately 20% of Multiple Myeloma (MM) cells and tumors (Chesi et al., 1997). This translocation results in ectopic expression of functional FGFR3 and promotes myeloma cell proliferation and prevents apoptosis (Plowright et al., 2000). Cappellen et al. found expression of a constitutively activated FGFR3 in a large proportion of 2 common epithelial cancers, bladder and cervix (Cappellen et al., 1999). These evidences indicate an oncogenic role for FGFR3 in carcinomas. Our previous results showed that inhibition of FGFR3 will decrease (VEGF) expression when we compared the gene expression profiles of FGFR3 inhibited samples with those of control samples in OPM-2 MM cells using microarray analysis (Zhu et al., 2005). This phenomena also exist in the KMS-11 MM cell line (Qian et al., 2005).
VEGF is a key mediator of tumor angiogenesis (Folkman, 1995). VEGF mediates its activity mainly via 2 Receptor Tyrosine Kinases (RTKs): Flt-1 and KDR. These receptors are localized on the cell surface of various endothelial cell types. Over-expression of VEGF has been associated with increased angiogenesis, tumor growth and metastasis in solid tumors as well as in hematologic malignancies (Folkman, 1995; Holmgren et al., 1995; Hanahan et al., 1996; Fidler and Ellis, 1994; Kim et al., 1993; Dankbar et al., 2000; Bellamy et al., 1999; Podar et al., 2001; Tartaglia et al., 2001). In MM, studies showed that bone marrow angiogenesis increases with progression from Monoclonal Gammopathy of Undetermined Significance (MGUS) to nonactive MM to active MM (Vacca et al., 1999, 1994). Importantly the angiogenic factor VEGF is secreted by MM cells as well as bone marrow stromal cells. VEGF, in addition to stimulating bone marrow angiogenesis, also plays an important role in both autocrine and paracrine growth of MM cells and that correlates with disease progression and poor prognosis (Vacca et al., 1999; Ribatti et al., 1999).
In order to better understand the molecular mechanism of FGFR3 and VEGF involvement in the pathogenesis of oncogenesis, we analysis VEGF expression in L6 cell line, which has no endogenous FGFRs, by stable transfection with plasmid pcDNA3 containing full-length cDNA encoding FGFR3. Present results revealed FGFR3 expression increase cell proliferation and up-regulate VEGF expression.
MATERIALS AND METHODS
All experiment was conducted at City of Hope National Medical Center between June 2004 to March 2005
Expression vector construction: A vector containing full-length human FGFR3 cDNA was a gift from Dr. Podolsky’s lab. To construct FGFR3 expression vector, a Hind III/Kpn I fragment was released from the vector containing full-length FGFR3 cDNA to remove the 3’ untranslated region. This fragment was subsequently ligated into a Hind III/Kpn I-digested pcDNA3 plasmid (Invitrogen, San Diego, CA).
Cell culture and transfection: The L6 rat myoblast cell line devoid of endogenous FGFR was cultured in Dulbecco’s modified Eagle’s medium (Cellgro, Mediatech Inc., Herndon, VA) with 100 IU mL-1 penicillin, 100 μg mL-1 streptomycin and 10% fetal calf serum (Sigma). Cells were transfected using Lipofectin (Life Technologies, Inc.) with 1 μg of pcDNA3 plasmid containing full-length human cDNA encoding FGFR3 and pcDNA3 plasmid only. Stable transfectants were established by selection in medium supplemented with 800 μg mL-1 G418 (Life Technologies, Inc.) for 3 weeks. This clone was designated L6F. The clone containing the pcDNA3 vector only was designated L6V.
KMS-11 cell line characterized by the t (4;14) and FGFR3 overexpression was kindly provided by Dr. P. Lief Bergsagel. Cells were cultured in RPMI 1640 medium supplemented with 10% Fetal Bovine Serum (FBS), 1% (v/v) penicillin and streptomycin (100 μg mL-1) and maintained at 37°C in a 5% CO2 atmosphere.
Northern analyses: Total cellular RNA was extracted from cells and Northern blot analysis was performed. RNA was separated on a 1% agarose gel and then transferred to a Hybond-N membrane. The filter was then probed with 32P-end-labeled cDNA fragment of human FGFR3. The filter was exposed to X-ray film and relative amounts of mRNA were quantitated by densitometer tracing of the autoradiogram.
Antibodies and Western Blot analysis: Human anti-VEGF polycolonal antibody was purchased from ZYMED (San Francisco, CA) and anti-FGFR3 polyclonal antibody, which recognizes the C-terminal FGFR3, was purchased from Santa Cruz Biotechnology (Santa Cruz, CA).
Cells were harvested, washed with ice-cold PBS and lysed in RIPA buffer (1xPBS, 1% NP40, 0.5% sodium deoxycholate, 0.1% SDS) with freshly added inhibitors (100 ug mL-1 phenylmethylsulfonyl fluoride (PMSF), 1 mM sodium orthovanadate and 30 μl mL-1 aprotinin). The samples were separated on 7.5% sodium dodecyl sulfate-polyacrylamide gel by loading equivalent amounts of protein. After electrophoresis, the proteins were transferred to a PVDF membrane (Amersham Pharmacia Biotech). The membrane was incubated with antibodies. The immunoblots were detected by the Western-Light™ system (Applied Biosystems. Fpster City, CA). PVDF membranes were stripped and reprobed with α-tubulin as loading control. Values of the relative amounts of protein were determined by the densities of the bands using Image Quant version 5.2.
Immunohistochemical analysis: Cells were trypsinized and washed with PBS. The cells were then frozen in -70°C as a tissue block . In brief, frozen sections (5 μm) were cut, transferred onto poly-L-lysine-coated slides, air-dried and then washed in PBS, followed by quenching of endogeneous peroxidase activity with 0.3% hydrogen peroxide in methanol. After being further rinsed with PBS, the sections were incubated with normal goat serum for 20 min at room temperature to block nonspecific binding and then incubated with primary anti-VEGF antibody at a 1:75 dilution for 14 h at 4°C. After being washed in 0.2% Triton X-100 in PBS, the sections were further incubated with biotinylated anti-rabbit IgG for 30 min at room temperature and then washed in 0.2% Triton X-100 in PBS. After the addition of streptavidin-biotin-conjugated peroxidase and incubation for 30 min at room temperature, the sections were washed in 0.2% Triton X-100 in PBS and the localization of the proteins was visualized by incubating the sections with 3,3’- diaminobenzidine. The slides were counterstained with Mayer’s hematoxylin, dehydrated in a graded alcohol series, cleared in xylene and mounted.
DNA-synthesis assay: The effects of FGFR3 transfection on DNA syntheses were determined by measuring the incorporation of radiolabeled thymidine into the nucleic acids of cells. 1x105 cells/well were plated into 6-well plates and incubated at 37°C in a 5% CO2 atmosphere. After 24 h, the cells reached about 70% confluence, 0.1 μM [3H] thymidine (0.5 μCi/mL) were added and incubated for 1 h at 37°C, then washed with ice-cold PBS. The nucleic acids were precipitated with ice-cold 10% Trichloroacetic Acid (TCA) and collected on Whatman GF/C filters. The filters were then washed 3 times with 5 mL of ice-cold 10% TCA followed by 2 washes with 95% ethanol, dried and incubated with 0.25 mL of 4 N HCI at 65°C for 1 h. The samples were then neutralized by addition of 0.4 mL of 2.5 N NaOH. After addition of 5 mL of scintillation fluid, the radioactivity in each sample was quantified by scintillation spectrometry.
Cell proliferation assay: CellTiter 96 Aqueous One Solution Cell Proliferation Assay Kit (Promega, Madison, WI, USA) were used to further validate cell proliferation. Briefly, 2,000 cells/well were seeded in quadruplicate in 96-well plates. Day 1, 2, 3 and 4 after plating, 20 μL of CellTiter 96 Aqueous One Solution Reagent were added into each well followed by a 2 h incubation at 37°C in a humidified, 5% CO2 atmosphere. The absorbance at 490 nm was recorded using a microplate autoreader (Bio-Tek instruments).
Enzyme-linked immunosorbent assay: The cells were collected and washed three times with 2 mL PBS. Under sonication, the proteins were extracted from each sample and then analyzed by VEGF-enzyme-linked immunosorbent assays (ELISA) according to the manufacturer’s protocol (R and D Systems, Minneapolis, MN). Briefly, 96-well plates were coated with anti-human VEGF antibody, rinsed and blocked with phosphate-buffered saline (PBS), 1% bovine serum albumin (BSA), 5% sucrose and 0.05% NaN3 for 2 h. After washing, sample or standards were added to the wells and incubated for 2 h at Room Temperature (RT). The wells were rinsed again and biotinylated anti-human VEGF antibody was added for 2 h at RT. After a further washing step, the wells were incubated with streptavidin horseradish peroxidase for 20 min at RT and rinsed again. The reaction was started by the addition of H2O2 and tetramethylbenzidine for 30 min at RT. After stopping the reaction with 1 M H2SO4 the optical density of each well was detected by means of a microplate autoreader at 450 nm. Each well was analyzed in triplicate.
RESULTS
FGFR3 expression in L6 cells: The L6 rat myoblast cell line has been used in this study because this cell line does not express any endogenous FGFRs. L6 cells were stably transfected with plasmid pcDNA3.1 containing full-length cDNA encoding FGFR3. Stably transfected clones were selected after prolonged culture in selection medium and designated as L6F (L6 cells expressing FGFR3) or L6V (L6 cells expressing vector only). The expression of FGFR3 was confirmed by Northern blot and Western blot. As shown in Fig. 1A, parental L6 cells and L6V clone demonstrated no detectable level of FGFR3 mRNA. Whereas, L6F transfectant expressed higher levels of FGFR3, but was relatively lower than KMS-11 cells, which are characterized by the t(4;14) and FGFR3 overexpression (Chesi et al., 1997) and used as positive control in this study. The protein expression of FGFR3 from cell lysates using an antibody that recognizes the carboxyl terminus of FGFR3 demonstrated that FGFR3 protein was indeed present in L6F clones but not in parental L6 Cells and L6V clones (Fig. 1B). This result was consistent with Northern blot analysis.
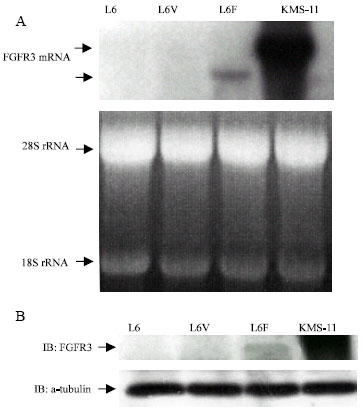 | |
| Fig. 1: | Expression of FGFR3 in L6F Clone. L6 cells, lacking endogenous FGFR3, were transfected using Lipofectin with pcDNA3.1 plasmid containing full-length human cDNA encoding FGFR3. (A) Northern blot analysis revealed the expression of the FGFR3 mRNA in the L6F clone. The arrows indicate mRNA from the endogenous FGFR3 (top) and the transfected FGFR3 (bottom). 28S rRNA and 18S rRNA were used as loading control. (B) Western blot analysis revealed the expression of FGFR3 protein in L6F clone. a-tubulin was used as loading control. KMS-11 cells were used as a positive control. |
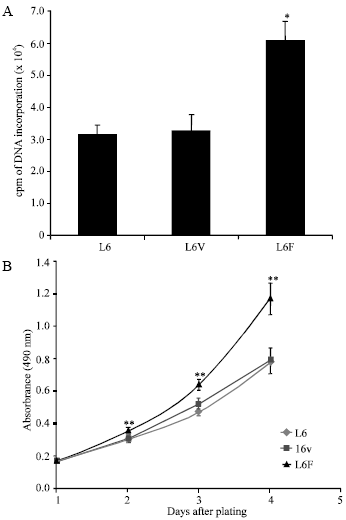 | |
| Fig. 2: | Effect of FGFR3 on DNA synthesis and cell proliferation. (A) [3H]-Thymidine incorporation was used to determine the effect of FGFR3 on DNA synthesis. Data represent means and SDs for triplicate experiments. (B) CellTiter 96 Aqueous One Solution reagent were used to test cell proliferation. Each point represents the mean±SD of quadruplicate. Statistical significance was determined by t-test: *p<0.05, **p<0.01 as compared with L6 control |
FGFR3 induces proliferation in L6F clone: To determine whether FGFR3 expression has effects on L6 cells, we then examined whether FGFR3 induced thymidine incorporation in DNA synthesis. As shown in Fig. 2A, L6F significant increased thymidine incorporation to 2 fold compared with parental L6 cells whereas [3H] thymidine uptake almost did not change in the L6V clone compared with parental L6 cells.
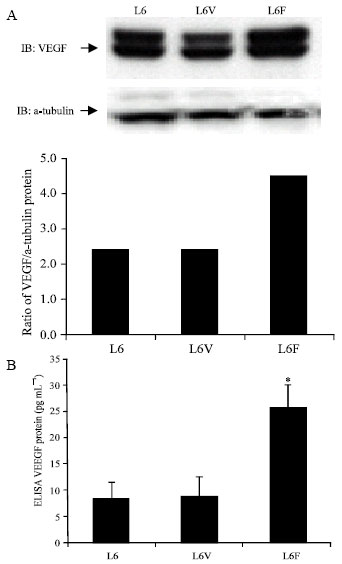 | |
| Fig. 3: | Expression of VEGF Protein in L6F Clone. (A) Western blot analysis and quantitative ratio were examined with anti-VEGF antibody and a-tubulin control as described in materials and methods. (B) The level of VEGF protein was measured by ELISA. Each experiment was performed in triplicate. Results are presented as (mean± SD). *p<0.05, t-test, compared with L6 control |
To further validate FGFR3 expression induced L6F cell proliferation, CellTiter 96 Aqueous One Solution Cell Proliferation Assay was conducted. As shown in Fig. 2B, L6F cells exhibited enhanced proliferation compared with L6 and L6V.
VEGF expression was increased in the L6F clone: To assess whether FGFR3 changes VEGF expression, parental L6 cell, L6V and L6F cells were harvested and VEGF protein expression were tested. As shown in Fig. 3A, VEGF expression was increased compared to L6 cells and L6V clones using Western blot. To further validate the increase of VEGF, we performed VEGF ELISA analyses to quantitate VEGF from cell lysates and found 2.5-fold increase in the L6F clone compared to parental L6 cells (Fig. 3B).
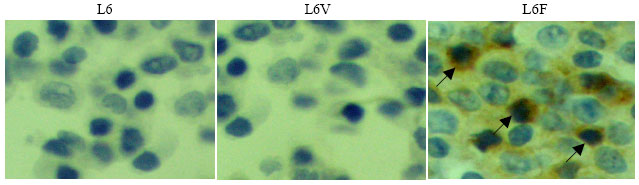 | |
| Fig. 4: | Immunohistochemical analysis of VEGF. Immunohistochemistry examined by antibody was performed as described in materials and methods. L6F showed significant increase of VEGF expression while there was no significant difference between L6 and L6V (magnification: 400x) |
Immunohistochemistry also showed that VEGF staining in the L6F clone was greater than in L6 or L6V (Fig. 4). These results clearly suggest that FGFR3 over-expression will up-regulate VEGF expression.
DISCUSSION
VEGF is a key mediator of tumor angiogenesis. Overexpression of VEGF has been associated with increased angiogenesis, tumor growth and metastasis in solid tumors and hematologic malignancies (Fidler et al., 1994; Kim et al., 1993; Dankbar et al., 2000; Bellamy et al., 1999; Podar et al., 2001). We previously found that inhibition of FGFR3 expression would decrease VEGF expression in MM cell lines (Zhu et al., 2005; Qian et al., 2005). This raised the question about the relationship between FGFR3 and VEGF. Therefore, the present study is aimed at investigating FGFR3 and VEGF. We stable transfected plasmid pcDNA3.1 containing full-length cDNA encoding FGFR3 into L6 cells, which has no endogenous FGFRs and subsequently detected overexpression of VEGF in L6F clones using Western blot, ELISA and immunohistochemistry. Present result clearly demonstrated that FGFR3 up-regulate VEGF expression. MM is an aggressive neoplastic disease characterized by the increased proliferation and extended life span of monoclonal plasma cells. Microvessel density of the bone marrow is increased in patients with active MM. Myeloma cells express and secrete VEGF (Dankbar et al., 2000; Bellamy et al., 1999; Vacca et al., 1999) and fibroblast growth factor (Bellamy et al., 1999; Vacca et al., 1999; Otsuki et al., 1999), both of which are considered potent angiogenic cytokines (Carmeliet and Jain, 2000) and are likely to contribute to the increased angiogenic potential of bone marrow plasma cells in progressive MM (Vacca et al., 1999). FGFR3 is normally not or very minimally expressed in plasma cells, but is overexpressed as a consequence of the chromosome translocation (Chesi et al., 1997, 2001; Otsuki et al., 1999). In some cases, activating mutations have also been found on the translocated allele (Ronchetti et al., 2001; Soverini et al., 2002). Activation of FGFR3 is a transforming event in haematopoietic cell allele (Chesi et al., 2001; Ronchetti et al., 2001; Li et al., 2001). Accumulated evidence demonstrated that FGFR3 play an oncogenic role in MM (Chesi et al., 2001; Plowright et al., 2000; Keats et al., 2003; Dvorak et al., 2003). In this study, we found that overexpression of FGFR3 into L6 cells caused increased DNA synthesis and proliferation.
In addition, many tumor cell lines, as well as freshly isolated hematological tumor cells, including MM cells, have been reported to produce and secrete VEGF (Dankbar et al., 2000; Bellamy et al., 1999; Podar et al., 2001; Fiedler et al., 1997; Dias et al., 1998), suggesting an autocrine VEGF loop in MM. Podar et al. (2001) reported that VEGF has direct effects on MM cells. VEGF stimulates proliferation and migration of MM cells by both autocrine and paracrine mechanisms. VEGF triggers tumor cell proliferation via a Raf-1-MEK-extracellular signal-regulated protein kinase pathway and migration via a PKC-dependent pathway (Podar et al., 2001). Preclinical studies demonstrated that inhibition of VEGF receptor blocks the growth and migration of MM cells in the bone marrow (Podar et al., 2004; Lin et al., 2002). Benvacizumab is a humanized form of a murine monoclonal antibody to VEGF. In a randomized clinical trial of bevacizumab in patients with metastatic renal cell cancer, the time to disease progression was prolonged but overall survival was not improved in patients (Yang et al., 2003).
From the published data we can conclude that both FGFR3 and VEGF play important roles in pathogenesis of MM or other cancers. But there is a lack of research that linked them together. In this study we demonstrated that FGFR3 can up-regulate VEGF expression. Though there is no direct evidence showing how FGFR3 up-regulate VEGF expression, some evidence demonstrated that FGFR3, through the MAPK and STAT3 pathway, enable cell proliferation and oncogenesis (Kanai et al., 1997; Chesi et al., 2001; Plowright et al., 2000). MAPK signaling cascade is essential for VEGF expression. Accumulated evidence showed that activating MAPK pathway would stimulate VEGF expression, while treatment with the MAPK inhibitor reduced expression of VEGF (Wu et al., 2003; Chelouvhe-Lev et al., 2004; Cassano et al., 2002). There was also direct evidence indicating that STAT3 protein binds to the VEGF promoter and STAT3 activity up-regulates VEGF expression using VEGF promoter mutagenesis and chromatic immunoprecipitation assays (Niu et al., 2002). Thus FGFR3 could, through MAPK and STAT3 pathway, up-regulate VEGF expression.
Our present study coupled with previous results indicated that FGFR3 might be associated with the regulation of VEGF. FGFR3 might mediate expression of VEGF and that may have a direct impact on the progression of myeloma or other cancers as well. Though complete elucidation of the role of FGFR3 in cancers requires further investigation, our observations suggest that the combination of inhibition of VEGF and inhibition of FGFR3 may be advantage for the treatment of FGFR3 overexpression cancer patients.
REFERENCES
- Bellamy, W.T., L. Richter, Y. Frutiger and T.M. Grogan, 1999. Expression of vascular endothelial growth factor and its receptors in hematopoietic malignancies. Cancer Res., 59: 728-733.
Direct Link - Cappellen, D., C. de Oliveira, D. Ricol, S. de Medina and J. Bourdin et al., 1999. Frequent activating mutations of FGFR3 in human bladder and cervix carcinomas. Nature Genet., 23: 18-20.
PubMedDirect Link - Carmeliet, P. and R.K. Jain, 2000. Angiogenesis in cancer and other diseases. Nature, 407: 249-257.
PubMedDirect Link - Cassano, A., C. Bagala, C. Battelli, S. Giovanni Q. Michela et al., 2002. Expression of vascular endothelial growth factor, mitogen-activated protein kinase and p53 in human colorectal cancer. Anticancer Res., 22: 2179-2184.
Direct Link - Chelouche-Lev, D., C.P. Miller, C. Tellez, M. Ruiz, M. Bar-Eli and J.E. Price, 2004. Different signalling pathways regulate VEGF and IL-8 expression in breast cancer: Implications for therapy. Eur. J. Cancer, 40: 2509-2518.
Direct Link - Chesi, M., E. Nardini, L.A. Brents, E. Schrock, T. Ried, W.M. Kuehl and P.L. Bergsagel, 1997. Frequent translocation t(4;14)(p16.3;q32.3) in multiple myeloma is associated with increased expression and activating mutations of fibroblast growth factor receptor 3. Nature Genet., 16: 260-264.
Direct Link - Chesi, M., L.A. Brents, S.A. Ely, C. Bais and D.F. Robbiani et al., 2001. Activated fibroblast growth factor receptor 3 is an oncogene that contributes to tumor progression in multiple myeloma. Blood, 97: 729-736.
PubMedDirect Link - Dankbar, B., T. Padro, R. Leo, B. Feldmann and M. Kropff et al., 2000. Vascular endothelial growth factor and interleukin-6 in paracrine tumor-stromal cell interactions in multiple myeloma. Blood, 95: 2630-2636.
PubMedDirect Link - Dias, S., R. Boyd and F. Balkwill, 1998. IL-12 regulates VEGF and MMPs in a murine breast cancer model. Int. J. Cancer, 78: 361-365.
Direct Link - Dvorak, P., D. Dvorakova, M. Doubek, J. Faitova, J. Pacholikova, A. Hampl and J. Mayer, 2003. Increased expression of fibroblast growth factor receptor 3 in CD34+ BCR-ABL+ cells from patients with chronic myeloid leukemia. Leukemia, 17: 2418-2425.
Direct Link - Fidler, I.J. and L.M. Ellis, 1994. The implications of angiogenesis for the biology and therapy of cancer metastasis. Cell, 79: 185-188.
PubMedDirect Link - Fiedler, W., U. Graeven, S. Ergun, S. Verago, N. Kilic, M. Stockschl�der and D.K. Hossfeld, 1997. Vascular endothelial growth factor, a possible paracrine growth factor in human acute myeloid leukemia. Blood, 89: 1870-1875.
PubMedDirect Link - Folkman, J., 1995. Angiogenesis in cancer, vascular, rheumatoid and other diseases. Nat. Med., 1: 27-30.
CrossRefDirect Link - Hanahan, D. and J. Folkman, 1996. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell, 86: 353-364.
PubMedDirect Link - Holmgren, L., M.S. O'Reilly and J. Folkman, 1995. Dormancy of micrometastases: Balanced proliferation and apoptosis in the presence of angiogenesis suppression. Nat. Med., 1: 149-153.
CrossRefDirect Link - Kanai, M., M. Goke, S. Tsunekawa and D.K. Podolsky, 1997. Signal transduction pathway of human fibroblast growth factor receptor 3. Identification of a novel 66-kDa phosphoprotein. J. Biol. Chem., 272: 6621-6628.
PubMedDirect Link - Keats, J.J., T. Reiman, C.A. Maxwell, B.J. Taylor and L.M. Larratt et al., 2003. In multiple myeloma, t(4;14)(p16;q32) is an adverse prognostic factor irrespective of FGFR3 expression. Blood, 101: 1520-1529.
PubMedDirect Link - Kim, K.J., B. Li, J. Winer, M. Armanini and N. Gillett et al., 1993. Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumour growth in vivo. Nature, 362: 841-844.
CrossRefDirect Link - Li, Z., Y.X. Zhu, E.E. Plowright, B.P. Leif and C. Marta et al., 2001. The myeloma-associated oncogene fibroblast growth factor receptor 3 is transforming in hematopoietic cells. Blood, 97: 2413-2419.
Direct Link - Lin, B., K. Podar, D. Gupta, Y.T. Tia and S. Li et al., 2002. The vascular endothelial growth factor receptor tyrosine kinase inhibitor PTK787/ZK222584 inhibits growth and migration of multiple myeloma cells in the bone marrow microenvironment. Cancer Res., 62: 5019-5026.
Direct Link - Martin, G.R., 1998. The roles of FGFs in the early development of vertebrate limbs. Genes Dev., 12: 1571-1586.
PubMedDirect Link - Niu, G., K.L. Wright, M. Huang, L. Song and E. Haura et al., 2002. Constitutive Stat3 activity up-regulates VEGF expression and tumor angiogenesis. Oncogene, 21: 2000-2008.
PubMedDirect Link - Otsuki, T., O. Yamada, K. Yata, H. Sakaguchi and J. Kurebayashi et al., 1999. Expression of fibroblast growth factor and FGF-receptor family genes in human myeloma cells, including lines possessing t(4;14)(q16.3;q32. 3) and FGFR3 translocation. Int. J. Oncol., 15: 1205-1212.
PubMedDirect Link - Plowright, E.E., Z. Li, P.L. Bergsagel, M. Chesi and D.L. Barber et al., 2000. Ectopic expression of fibroblast growth factor receptor 3 promotes myeloma cell proliferation and prevents apoptosis. Blood, 95: 992-998.
Direct Link - Podar, K., Y.T. Tai, F.E. Davies, S. Lentzsch and M. Sattler et al., 2001. Vascular endothelial growth factor triggers signaling cascades mediating multiple myeloma cell growth and migration. Blood, 98: 428-435.
PubMedDirect Link - Podar, K., L.P. Catley, Y.T. Tai, R. Shringarpure and P. Carvalho et al., 2004. GW654652, the pan-inhibitor of VEGF receptors, blocks the growth and migration of multiple myeloma cells in the bone marrow microenvironment. Blood, 103: 3474-3479.
PubMedDirect Link - Qian, S., G. Somlo and B. Zhou, L. Zou and S. Mi et al., 2005. Ribozyme cleavage leads to decreased expression of fibroblast growth factor receptor 3 in human multiple myeloma cells, which is associated with apoptosis and downregulation of vascular endothelial growth factor. Oligonucleotides, 15: 1-11.
PubMedDirect Link - Ribatti, D., A. Vacca, B. Nico, F. Quondamatteo and R. Ria et al., 1999. Bone marrow angiogenesis and mast cell density increase simultaneously with progression of human multiple myeloma. Br. J. Cancer, 79: 451-455.
Direct Link - Ronchetti, D., A. Greco, S. Compasso, G. Colombo and P. Dell-Era et al., 2001. Deregulated FGFR3 mutants in multiple myeloma cell lines with t(4;14): comparative analysis of Y373C, K650E and the novel G384D mutations. Oncogene, 20: 3553-3562.
Direct Link - Soverini, S., C. Terragna, N. Testoni, R. Deborah and T. Patrizia et al., 2002. Novel mutation and RNA splice variant of fibroblast growth factor receptor 3 in multiple myeloma patients at diagnosis. Haematologica, 87: 1036-1040.
Direct Link - Tartaglia, M., A. Fragale and P.A. Battaglia, 2001. A competitive PCR-based method to measure human fibroblast growth factor receptor 1-4 (FGFR1-4) gene expression. DNA Cell Biol., 20: 367-379.
Direct Link - Vacca, A., D. Ribatti, L. Roncali, G. Ranieri, G. Serio, F. Silvestris and F. Dammacco, 1994. Bone marrow angiogenesis and progression in multiple myeloma. Br. J. Haematol., 87: 503-508.
Direct Link - Vacca, A., D. Ribatti, M. Presta, M. Minischetti and M. Iurlaro et al., 1999. Bone marrow neovascularization, plasma cell angiogenic potential and matrix metalloproteinase-2 secretion parallel progression of human multiple myeloma. Blood, 93: 3064-3073.
PubMedDirect Link - Wu, J., S. Brandt and S.M. Hyder, 2005. Ligand- and cell-specific effects of signal transduction pathway inhibitors on progestin-induced vascular endothelial growth factor levels in human breast cancer cells. Mol. Endocrinol., 19: 312-326.
CrossRefDirect Link - Wu, Y., Q. Zhang, D.K. Ann, A. Akhondzadeh, H.S. Duong, D.V. Messadi and A.D. Le, 2003. Increased vascular endothelial growth factor may account for elevated level of plasminogen activator inhibitor-1 via activating ERK1/2 in keloid fibroblasts. Am. J. Physiol. Cell Physiol., 286: C905-912.
CrossRefDirect Link - Yang, J.C., L. Haworth, R.M. Sherry, P. Hwu and D.J. Schwartzentruber et al., 2003. A randomized trial of bevacizumab, an anti-vascular endothelial growth factor antibody, for metastatic renal cancer. N. Engl. J. Med., 349: 427-434.
Direct Link - Zhu, L., G. Somlo, B. Zhou, J. Shao and V. Bedell et al., 2005. Fibroblast growth factor receptor 3 inhibition by short hairpin RNAs leads to apoptosis in multiple myeloma. Mol. Cancer Ther., 4: 787-798.
PubMedDirect Link