
S. M. Sherif
Biophysics Unit, Research Institute of Ophthalmology, 2 Al-Ahram Street: P.O. Box Giza, Egypt
A. A. Salwa
Biophysics Unit, Research Institute of Ophthalmology, 2 Al-Ahram Street: P.O. Box Giza, Egypt
International Journal of Pharmacology
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 298-302







ABSTRACT
Dexamethasone, one of the most potent glucocorticoids, is very effective anti-inflammatory drug. A side effect associated with long-term treatment of various diseases with steroids is a high incidence of posterior subcapsular cataracts. The literature did not indicate any reports discuss if the treatment with 0.1% dexamethasone has any effect(s) on the lens capsule or not. To answer this the lens capsule lipids was studied using different techniques. NewZealand albino rabbits were used in this study, where three groups were involved; control, 3DEXA and 5DEXA groups. Dexamethasone was instilled in both eyes four times a day for three weeks (3DEXA) and for five weeks (5DEXA). The measured parameters: total lipids, cholesterol, phospholipids and total protein as well as lipid spectroscopy (UV and FTIR) revealed significant changes in the capsular thickness as well as the physical characteristics of the capsular lipid bilayer.
PDF Abstract XML References Citation
How to cite this article
S. M. Sherif and A. A. Salwa, 2006. Long-Term Application of 0.1% Dexamethasone May Affect the Lens Capsular Lipids. International Journal of Pharmacology, 2: 298-302.
DOI: 10.3923/ijp.2006.298.302
URL: https://scialert.net/abstract/?doi=ijp.2006.298.302
DOI: 10.3923/ijp.2006.298.302
URL: https://scialert.net/abstract/?doi=ijp.2006.298.302
INTRODUCTION
Glucocorticoids (GC) play an important physiological role and have been widely used as valuable therapeutic agents for various diseases (Jobling et al., 2001). Endogenous GC are life-sustaining cholesterol derivatives produced in the zona fasciculate of the adrenal cortex and play a crucial role in maintaining cell homeostasis and viability of the organism and required to maintain normal carbohydrate, lipid and protein metabolism. GC are postulated to enhance wound healing, maintenance of cardiac vascular integrity and cardiac contractility and various other functions.
Externally applied GC are widely used principally to capitalize on their ability to inhibit inflammatory responses. In addition to the appearance of GC on tissues that have GC receptors, some of their adverse effects are observed in non target tissues for GC and are probably produced by changes in blood components, such as hyperglycemia and hyperlipidemia that result from the effects of GC on the main target tissue, liver (Watanabe et al., 2000). Steroid-induced cataracts were first documented by Black et al. (1960).
Cataract surgery generates short term inflammation, which is routinely controlled by applying anti-inflammatory drugs both before the surgery and for 2-6 weeks after surgery. Glucocorticoids are widely used for this purpose (Simone and Whitacre, 2001). The prolonged use of systemic (Black et al., 1960), topical (Laurell and Zetterstrom, 2002) or inhaled (Fraunfelder and Meyer, 1990) GC is known to be a high risk factor in the formation of posterior subcapsular cataracts.
Lens capsule is regarded as metabolically inert structure; contains the lens fiber and transmitting zonular tension in the unaccommodated lens. It is usually pictured as purly mechanical barrier to the passage of very large colloidal molecules or particles, but having no active effect on transport (Davson, 1980). There is no solid data are available regarding whether the long-term application of dexamethasone has a direct effect on lens capsule. Therefore in the present study, we attempted to clarify this, which may be a leading cause for enhancing and/or increasing the complications of the steroid-induced posterior subcapsular cataracts.
MATERIALS AND METHODS
Eyes were obtained from adult albino rabbits weighing 2.0-2.5 kg. The rabbits were selected from the animal house of the Research Institute of Ophthalmology and were classified into three groups (each with n = 8); normal (control) group and the other two groups were received topical instillation of 0.1% dexamethasone in both eyes for three weeks (3DEXA) and for five weeks (5DEXA), respectively. The anti-inflammatory corticosteroid, dexamethasone, was administered four times a day.
At the end of each period, the lenses were immediately dissected from the eyes and, using fine forceps, the lens capsules were separated from the fiber mass. The previously weighed lens capsules from all groups were homogenized separately in 1 mL bi-distilled water using cell homogenizer type Edmund Buhler 7400 Tübingen HO4. The resulted homogenate was centrifuged, then the supernatant was used to estimate the protein content following Lowry et al. (1951) procedure. The rest of the homogentaes were subjected to lipid extraction and purification according to Folch et al. (1957). All solvents used in lipid extraction were bubbled with nitrogen and during extraction nitrogen was frequently blown over the sample to prevent lipid oxidation.
The typical absorption spectra of the extracted lipids were recorded using spectrophotometer type Shimadzu uv-visible 240a (Japan). Total lipids, cholesterol and phospholipids were determined in an aliquotes of the extracted lipids after evaporating the solvent as described by Knight et al. (1972), Watson (1960) and Broekhyse (1968), respectively.
The infrared spectra were acquired with a Shimadzu Fourier transform infrared spectrometer model 8200 SNO (Japan), equipped with a standard detector. Measurements were made with IR cell (KBr windows). Interferograms were recorded, coadded and apodized with a Happ-Genzel function prior to fourier transform yielding an effective resolution of 2.0 cm-1 according to William et al. (1996). All the spectra were recorded at physiological temperature.
The resulted data were statistically evaluated using ANOVA procedure (SPSS package). The significance level was set at p<0.05.
RESULTS
The typical absorption spectra of lipids extracted from different lens capsules are given in Fig. 1. In the normal pattern, the maximum at 206-210 nm region corresponds to absorption of isolated double bonds of hydrocarbon chains and the shoulder at 274 nm with significant smaller amplitude corresponds to triene conjugates. This normal pattern mimic the normal pattern determined by Babizhayev (1989). In case of oxidized lipids, the uv-absorption pattern indicates an absorption maximum at 230 nm corresponds to primary lipid peroxidation products and the peak at 274 nm characterized by increased intensity relative to the normal one. According to this figure it is obviously clear that the extracted lipids from 3DEXA and 5DEXA groups were characterized by the same two peaks as the normal. In addition, the maximum at 230 nm is virtually absent and the intensity of the band at 274 nm mimic the normal one. This implies that there is no detectable lipid peroxidation products.
Table 1 shows that both total lipids and total protein were significantly increased (p<0.01) in 3DEXA and 5DEXA groups relative to the normal values. While, both cholesterol and phospholipids contents were significantly decreased (p<0.01) in both groups.
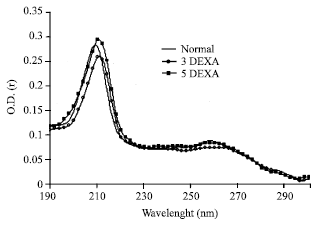 | |
| Fig. 1: | Typical absorption spectra of extracted lens capsule lipids from all studied groups, where an equal amounts were used |
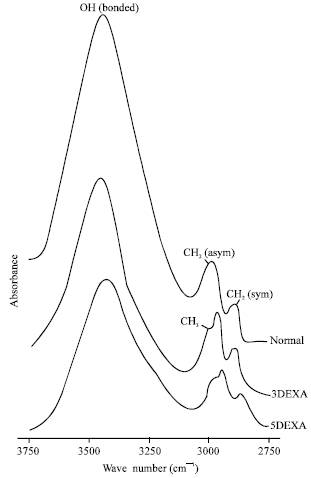 | |
| Fig. 2: | Infrared spectra of OH/CH2 stretching region (3500-2800 cm-1) of dry lipids from normal, 3DEXA and 5DEXA groups at physiological temperature |
The infrared stretching region at 3100-2800 cm-1 (Fig. 2) was used to investigate the lipid order of the fatty acyl chain. In the normal pattern two bands are discernable near 2930 and 2850 cm-1 correspond to the CH2 asymmetric and CH2 symmetric stretch, respectively.
| Table 1: | The estimated total lipids, cholesterol, phospholipids and total protein for the different studied groups. |
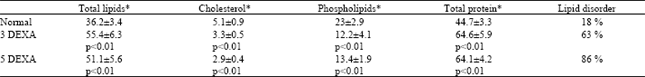 | |
| * Expressed as mg g-1 lens capsule, The last column shows the calculated percentage of lipid disorder as mentioned in the text. | |
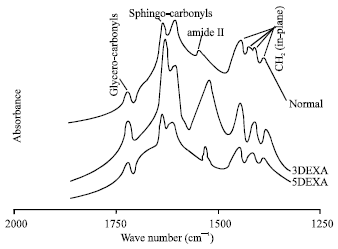 | |
| Fig. 3: | Infrared fingerprint region (1800-1300 cm-1) of dry lipids from normal, 3DEXA and 5DEXA groups at physiological temperature |
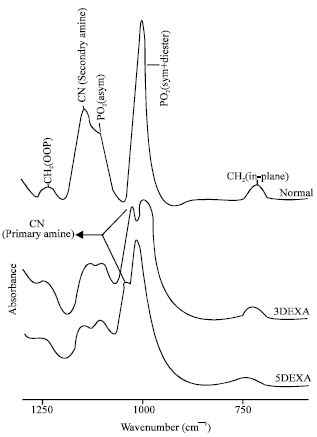 | |
| Fig. 4: | Infrared fingerprint region (1250-600 cm-1) of dry lipids from normal, 3DEXA and 5DEXA groups at physiological temperature |
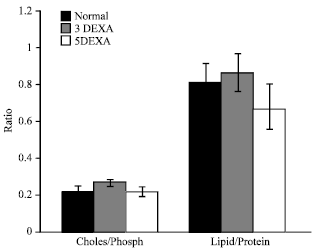 | |
| Fig. 5: | The calculated ratios for cholesterol/ phospholipids and total lipids/total protein in all studied groups |
In animals received topical dexamethasone for three weeks (3DEXA), this region shows three bands, where an additional band at 2980 cm-1 was appeared and assigned as CH3 symmetric terminal stretch. As the instillation period of 0.1% dexamethasone increased to five weeks (5DEXA group), these three bands are still present.
The frequency of the CH2 symmetric stretching band reflects the order of the lipid hydrocarbon chain (i.e., structural fluidity). Based upon mathematical analysis described by Borchman et al. (1993), Table 1 shows the percentage of lipid disorder (fluidity) of both 3DEXA and 5DEXA groups relative to the normal. It is clear that the lipid disorder increased as the instillation period of 0.1% dexamathasone increased; 63 and 86% for 3DEXA and 5DEXA groups, respectively.
The OH (H-bonded) group (3600-3100 cm-1) reflects the degree of lipid peroxidation and the amount of hydroxyl-containing lipids such as sphingolipids and cholesterol. Since we did not detect lipid peroxides in the uv-spectra, therefore this band arises from hydroxyl-containing lipids. The intensity of this band was decreased as the dexamethasone period increased relative to the normal one. This relative intensity indicate the relative abundance of this hydroxyl-containing lipids (Fig. 2).
The infrared spectra of the fingerprint region (1800-500 cm-1) provide us with a wealth of information. Figure 3 shows the 1800-1300 cm-1 region. The asymmetric band around 1740 cm-1 is due to lipids with glycerol backbone (glycero-carbonyls) as phosphatidylcholine or phosphatidylethanolamine. The intensity of this band increased in 3DEXA group only compared to the normal. The bands in infrared region between 1680 and 1620 cm-1 arise from the acyl-linked hydrocarbon chains of lipids with sphingosine backbone (sphingo-carbonyls). Two absorption bands are found in all studied groups around 1653 and 1629 cm-1. The intensity of the band at 1653 cm-1 was found to be increased as compared with the normal; the increase was dominant in 3DEXA group. While, the other band at 1629 cm-1 was found to be decreased in both dexamethasone treated groups, the decrease is also dominant in 3DEXA group.
For the IR band around 1550 cm-1 that results from the stretching mode of the sphingolipid amide II, it is clear that the intensity of this band increased in both dexamethasone treated groups, the increased intensity was obvious in the 3DEXA group.
The main absorption band around 1464 cm-1 results from the scissoring motion of the methylene CH2 segments of the hydrocarbon chains. The frequency, intensity and the number of components associated with this band reflects the form of lipid packing in the bilayer. Three absorption bands had detected in that region for both dexamethasone treated groups. The intensities decreased as the instillation period increased from 3 to 5 weeks. On the other hand, the normal pattern indicates four bands. Thus it is obvious that the in-plane (scissoring) motion of the CH2 segments in 3DEXA and 5DEXA groups had been decreased with reduction in the number of these segments.
Figure 4 shows the fingerprint region 1320-600 cm-1. The bending vibrational mode of CH2 bond within the fingerprint region can be monitored by observing the absorption bands around 1250 cm-1 (out of plane, OOP) and ≈731 cm-1 (rocking, inplane). The intensities of both bands were diminished by dexamethasone treatments. The structural changes within the phospholipids can be noticed by monitoring the absorption bands at 1190-1130 cm-1 (secondary amine), 1161 cm-1 (PO2- asymmetric stretch), 1090-1020 cm-1 (primary amine) and 1028 cm-1 (PO2¯ symmetric and diester stretch -C-O-P-O-C). In the normal lipid pattern only the primary amine band is present while, in 3DEXA and 5DEXA groups, both primary and secondary amine bands are present. The intensity of PO2¯ asymmetric stretch was increased as the time of instillation increased to 5 weeks. While, PO¯2 symmetric (and diester) decreased as compared with the normal pattern.
DISCUSSION
Glucocorticoids were used to suppress post-operation inflammation. High dose and long term therapy is well known to have adverse effects as glaucoma, cataract, change in corneal morphology and function and recently James et al. (2005) found that steroid treatment of lens epithelial cells is associated with significant changes in gene expression.
The typical absorption spectra of lens capsule lipids revealed no changes in both groups treated with dexamethasone, as well as no detectable lipid peroxidation products. The undetectable lipid peroxides is a good sign that the endogenous mechanism (biochemical event, oxidative degradation) for injury of the lipid bilayer not established all over the treatment periods, 5 weeks. The paradox association between the increased total lipids and the decreased cholesterol and phospholipids contents may resulted from another lipoidal material that incorporated within lens capsules. Biological membranes contain three kinds of major membrane lipids; phospholipids, glycolipids and sterols. In ocular membranes, phospholipids and cholesterol are essential components (Borchman et al., 1989), therefore we can attribute the increase in total lipids to an increased content of glycolipids.
The significant changes in the lens capsule constituents: total lipids, phospholipids, cholesterol and total protein will affect the thickness of the capsule, but as shown in Fig. 5 their relative ratios in all groups are oscillated around mean fixed value; i.e., non-significantly changed among different groups (0.78 for lipid/protein ratio and 0.2 for cholesterol/phospholipids one). This interesting finding shows that although lens capsules are suffering from variation in their thickness, they are biologically still able performing their normal function and the changes in the individual components is just to buffer the induced changes resulted from the treatment with 0.1% dexamethasone.
The decrease in cholesterol and phospholipids contents was also confirmed by the decreased intensity of the OH (H-bonded) group. Also, the parallel decrease in the IR band at ≈1028 cm-1 (PO¯2 and diester stretch) implies that phospholipids, particularly phospholipids containing amide groups are the most affected component of the lens capsule lipids due to 0.1% dexamethasone treatments. It is known that cholesterol adds stability to the phospholipid bilayer of membranes (Gimple et al., 1997) and phospholipids are responsible of membrane fluidity, thus decreasing both concentrations may be regarded as changes in the physical characteristics of the capsule; fluidity and stability due to the treatment with 0.1% dexamethasone.
In conclusion, it remains uncertain whether the reported changes resulted as a direct effect of treatment, so that we can suggest the presence of dexamathasone receptor(s) within the lens capsule, or secondary to the damage takes place in the epithelial cell layer (posterior subcapsular cataract). Regardless the origin of these changes, the lens capsules can buffer them by changing the physical characteristics of the capsular lipids with little biochemical contribution. Sato et al., (1996) show that protein did not change lipid structure even at protein/lipid weight ratio of two to one. There is an insisting need to know whether these changes persist such that play a role in the overall effects of dexamathasone or it is a transient effect of the treatment. So, the phospholipid-cholesterol rafts should be studied extensively taking into account the effect of protein.
REFERENCES
- Babizhayev, M.A., 1989. Accumulation of lipid peroxidation products in human cataracts. Acta Ophthalmol., 67: 281-287.
PubMedDirect Link - Black, R.L., R.B. Oglesby, J. von Sallmann and J.J. Bunim, 1960. Posterior subcapsular cataracts induced by corticosteroids in patients with rheumatoid artheitis. J. Am. Med. Assoc., 174: 166-171.
PubMedDirect Link - Borchman, D., O.P. Lamba and M.C. Yappert, 1993. Structural characterization of lipid membranes from clear and cataractous human lenses. Exp. Eye Res., 57: 199-208.
PubMedDirect Link - Folch, J., M. Lees and G.H.S. Stanley, 1957. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem., 226: 497-509.
CrossRefPubMedDirect Link - Fraunfelder, F.T. and S.M. Meyer, 1990. Posterior subcapsular cataracts associated with nasal or inhalation corticosteroids. Am. J. Ophthalmol., 109: 489-490.
PubMedDirect Link - Gimpl, G., K. Burger and F. Fahrenholz, 1997. Cholesterol as modulator of receptor function. Biochemistry, 36: 10959-10974.
PubMedDirect Link - James, E.R., V.M. Fresco and L.L. Robertson, 2005. Glucocorticoid-induced changes in the global gene expression of lens epithelial cells. J. Ocul. Pharmacol. Ther., 21: 11-27.
PubMedDirect Link - Jobling, A.I., A. Stevens and R.C. Augusteyn, 2001. Binding of dexamethasone by α-crystallin. Invest. Ophthalmol. Vis. Sci., 42: 1829-1832.
Direct Link - Knight, J.A., S. Anderson and J.M. Rawle, 1972. Chemical basis of the sulfo-phospho-vanillin reaction for estimating total serum lipids. Clin. Chem., 18: 199-202.
CrossRefPubMedDirect Link - Laurell, C.G. and C. Zetterstrom, 2002. Cancer induction and effect of Strobilanthes crispus leaf extract on liver histology and enzymes changes in rats. Br. J. Ophthalmol., 86: 1380-1384.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Simone, J.N. and M.M. Whitacre, 2001. Effects of anti-inflammatory drugs following cataract extraction. Curr. Opin. Ophthalmol., 12: 63-67.
PubMedDirect Link - Watanabe, H., H. Kosano and H. Nishigori, 2000. Steroid-induced short term diabetes in chick embryo: Reversible effects of insulin on metabolic changes and cataract formation. Invest. Ophthalmol. Vis. Sci., 41: 1846-1852.
PubMedDirect Link - William, L.D., A.D. Nicholas, B. Douglas and R.P. Ahuja, 1996. Studies on lipids and the activity of Na-K ATPase in lens fiber cells. Biochem. J., 314: 961-967.
Direct Link