
Sheyda Hoseini
Laboratory of Toxicology, Faculty of Pharmacy, and Pharmaceutical Sciences Research Centre, Tehran University of Medical Sciences, Tehran, Iran
Hadi Esmaily
Laboratory of Toxicology, Faculty of Pharmacy, and Pharmaceutical Sciences Research Centre, Tehran University of Medical Sciences, Tehran, Iran
Azadeh Mohammadirad
Laboratory of Toxicology, Faculty of Pharmacy, and Pharmaceutical Sciences Research Centre, Tehran University of Medical Sciences, Tehran, Iran
Mohammad Abdollahi
Laboratory of Toxicology, Faculty of Pharmacy, and Pharmaceutical Sciences Research Centre, Tehran University of Medical Sciences, Tehran, Iran
International Journal of Pharmacology
Year: 2006 | Volume: 2 | Issue: 3 | Page No.: 280-285







ABSTRACT
In the present study, the effects of short-term administration of sildenafil a selective PDE5 inhibitor on serum glucose and hepatic glycogenolysis and gluconeogenesis were examined in rats in vivo. Sildenafil was administered intraperitoneally at doses of 0.5, 1.0 and 5.0 mg kg-1. Two hours post treatment, liver was perfused, removed and homogenized. The activities of the key enzymes of glycogen phosphorylase (GP) and phosphoenolpyruvate carboxykinase (PEPCK) were analyzed in the homogenate. The high dose of sildenafil (5 mg kg-1) caused a significant reduction in serum glucose levels in comparison to control group (94.6±4.7 vs. 121.16±6.05 mg dL-1). The activity of PEPCK remained unchanged when animals were treated with various doses of sildenafil. Interestingly, sildenafil reduced hepatic GP at all doses administered (20.31±1.42, 17.18±1.2, 15.69±1.1 vs. 25.8±1.8 U g-1 liver protein for control group). It is concluded that administration of sildenafil markedly reduces liver glycogenolysis which in turn lowers blood glucose concentration at higher doses. This effect of sildenafil seems to be in relation with its NO mimicking potential and antioxidant properties. The underlying mechanisms involved seem complicated since parallel increase in the activities of GP and PEPCK was not observed.
PDF Abstract XML References Citation
How to cite this article
Sheyda Hoseini, Hadi Esmaily, Azadeh Mohammadirad and Mohammad Abdollahi, 2006. Effects of Sildenafil a Phosphodiesterase 5 Inhibitor on Rat Liver Cell Key Enzymes of Gluconeogenesis and Glycogenolysis. International Journal of Pharmacology, 2: 280-285.
DOI: 10.3923/ijp.2006.280.285
URL: https://scialert.net/abstract/?doi=ijp.2006.280.285
DOI: 10.3923/ijp.2006.280.285
URL: https://scialert.net/abstract/?doi=ijp.2006.280.285
INTRODUCTION
The phosphodiesterases phosphodiesterase (PDEs) are a superfamily of enzymes which catalyze the hydrolysis of the cyclic nucleotides cAMP, cGMP to their corresponding inactive 5-monophosphate counterparts. The cyclic nucleotides play a prominent role in the regulation of cellular functions and PDE inhibition can therefore elicit a variety of effects (Dal-Piaz and Giovannoni, 2000). There are 11 families of PDE enzymes (PDE1-PDE11) which are recognized by substrate specificity, kinetic characteristics and amino acid sequence. Sildenafil is a potent and selective inhibitor of PDE5 that decreases the breakdown of endogenously produced cGMP in smooth muscle cells and nerves (Ballard et al., 1998) and is currently used for treatment of male erectile dysfunction (Souness et al., 2000). The pharmacologic actions of phosphodiesterase inhibitors (PDEIs) are due to their potential to increase intracellular cAMP and cGMP as intracellular signal messengers that modulate both the intensity and the nature of immediate and delayed cellular responses (Soderling and Beavo, 2000).
The recent studies in animals (Milani et al., 2005) and human (Rahimi et al., 2005; Aydin et al., 2001) have demonstrated an impact of PDE5 inhibitors upon glucose metabolism but the reports are controversial. In some studies, cGMP accumulation in the liver cells has been shown to have little effect on glycogenolysis (Atefi et al., 2004; Pointer et al., 1976) while on the other hand, the inhibition of hepatocyte protein synthesis and gluconeogenesis caused by cytokines and nitric oxide (NO) has been attributed to an increase in cGMP (Parker et al., 1997) level. Also, T-1032, a new cGMP PDE-5 inhibitor has shown potent inhibitory influence on the action of physiologic insulin in capillary recruitment and glucose uptake (Mahajan et al., 2003). There is evidence that liver is one of the organs that well responds to PDE inhibitors by influencing glucose output (Abdollahi et al., 2003a). The liver plays a major role in blood glucose homeostasis by maintaining a balance between the uptake and storage of glucose via glycogenesis and the release of glucose via glycogenolysis and gluconeogenesis. If the liver is overloaded with glucose, a hepatic steatosis can develop together with high glycogen content. Conversely, if glucose availability in the diet is reduced, glucose-utilizing pathways are inhibited and glucose-producing pathways are activated. During fasting, glucose is produced by the liver glycogenolysis. Then, when the glycogen stores are depleted, glucose is produced de novo from precursors such as lactate, alanine or fructose through a pathway called gluconeogenesis. The regulation of these metabolic pathways involves the rapid modulation of the activity of specific proteins (enzymes, transporters) and modulating their transcription rate or post-transcriptional steps such as mRNA half-life and translation efficiency (Abdollahi et al., 2003b; Foufelle and Ferre, 2002; Nordlie et al., 1999). Regarding above-mentioned reports, the aim of this study was to examine whether effect of administration of sildenafil (selective PDE5 inhibitor) might affect activities of hepatic key enzymes of gluconeogenesis and glycogenolysis including phosphoenolpyruvate carboxykinase (PEPCK) and glycogen phosphorylase (GP) in rat.
MATERIALS AND METHODS
Materials: All chemicals were obtained from Sigma-Aldrich (Dorset, England).
Animals: Adult male Wistar rats, supplied by the animal house of Faculty of Pharmacy, Tehran University of Medical Sciences, weighing 200-250 g were used in this study. All animals were housed under standard laboratory conditions and fed normal laboratory rat chow and water.
Treatment: The animals were housed under controlled environmental conditions of temperature (22±2°C) and a 12 h light/dark cycle. Animals were allowed to acclimatize for 7 days. They were fed with standard rat chow and had free access to food and water. The protocol of study was approved by TUMS/PSRC ethic committee.
Sampling: Sildenafil was administered intraperitoneally at doses of 0.5, 1.0 and 5.0 mg kg-1. Two hours later, the liver was removed under anesthesia by transverse abdominal incision and perfusion with cold 0.9% saline. The weight of liver was recorded and kept frozen at -70°C until homogenized. The activities of the key enzymes of phosphoenolpyruvate carboxykinase (PEPCK) and glycogen phosphorylase (GP) were determined in liver homogenate to identify changes in glycogenolysis and gluconeogenesis respectively, as described previously (Saadat et al., 2004; Abdollahi et al., 2004a). Blood sample was taken under anesthesia by cardiac puncture and serum was separated and kept at -20°C for glucose assay.
Glucose assay: Serum glucose level was measured in the presence of glucose oxidase and peroxidase using o-dianisidine-HCl as a chromogen. The amount of glucose formed is related to amount of o-dianisidine oxidation products that measured spectrophotometrically at 436 nm (Bergmeyer et al., 1974).
Determination of PEPCK activity in liver: The liver homogenate was mixed with 10 volumes of cold 0.2 M phosphate buffer, pH 7.4, containing 2 mM mercaptoethanol and 2 nM EDTA and was then mixed and centrifuged for 10 min at 60000xg at 0°C and the supernatant was used for enzyme activity assay. Enzymatic activity was assayed in the reverse direction, carboxylation of phosphoenolpyruvate to form oxaloacetic acid in the presence of NADH as previously described (Chang and Lane, 1966). PEPCK activity is expressed as unit per gram of liver protein.
Determination of GP activity in liver: The liver homogenate was centrifuged at 30000xg for 30 min and the supernatant was used for enzyme activity assay. Enzymatic activity was assayed in the direction of glycogen breakdown by measuring the reduction of NADP (Lowry et al., 1967). GP activity is expressed as unit per gram of liver protein.
Determination of liver protein concentration: The liver homogenate was centrifuged as described above and the total protein concentration was measured in the supernatant using bovine serum albumin as standard by Lowry procedure (Lowry et al., 1951).
Statistical analysis: The results were analyzed for statistical significance by one-way ANOVA test and Tukey’s post hoc multi-comparison. All data were expressed as means±SEM of six animals in each group. A p<0.05 was considered statistically significant.
RESULTS
Administration of sildenafil at doses of 0.5 and 1.0 mg kg-1 did not alter serum glucose concentration but the dose of 5.0 mg kg-1 caused a significant reduction (22.41%, p<0.01) in comparison to the control group (94.6±4.7 vs. 121.16±6.05 mg dL-1, Fig. 1).
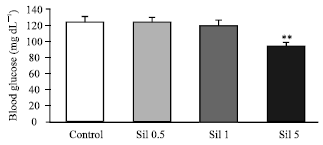 | |
| Fig. 1: | Effects of sildenafil on rat blood glucose concentration. Sildenafil (Sil) was administered in different doses of 0.5, 1 and 5 mg kg-1. Values are expressed as mean±SE of 6 animals in each group. ** Significantly different from control at p<0.01 |
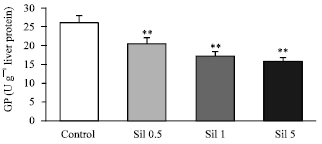
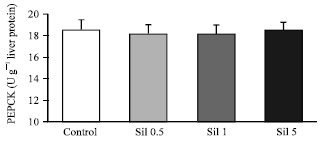
Administration of sildenafil at doses of 0.5, 1.0 and 5.0 mg kg-1 reduced the hepatic GP activity in comparison to control group (20.31±1.42, 17.18±1.2, 15.69±1.1 vs. 25.8±1.8 U g-1 liver protein, Fig. 2) with a mean percentage of decreases of 21, 33 and 39%, respectively. The activity of PEPCK was not changed after administration of sildenafil (Fig. 3).
DISCUSSION
The results of this study indicated that in vivo treatment of rats by sildenafil dose-dependently reduces hepatic GP activity without affecting PEPCK. Interestingly, only high dose of sildenafil affected blood glucose and reduced its level. GP is the key enzyme of glycogen breakdown that catalyzes the degradative phosphorolysis of glycogen to glucose-1-phosphate. This enzyme is promoted by substrate as well as allosteric effectors, including adenosine monophosphate (AMP) and phosphorylation (Barford and Johnson, 1989). There is evidence that glucagon and other cAMP-increasing agents exert a glycogenolytic effect by maintaining GP in a phosphorylated active state (Bollen et al., 1998). cAMP or Ca2+ dependent glycogenolytic agents cause glycogen degradation through specific protein kinases which may activate the GP. Considering the ability of sildenafil to reduce hepatic GP activity, one mechanism would be that sildenafil inhibits cAMP-dependent processes. To explain this, it should be noted that increasing cGMP is a fundamental mechanism of action of sildenafil. Our recent study on rat hepatocytes in vitro indicated that sildenafil increases hepatocytes cGMP levels but does not significantly alter glycogenolysis and gluconeogenesis (Atefi et al., 2004; Abdollahi et al., 2003c; Abdollahi and Simaiee, 2003). That is partly in agreement with the present findings when considering that blood glucose does not significantly change with normal doses of sildenafil. Sildenafil, through the inhibition of PDE5 and augmentation on the action of NO-cGMP axis shows similar effects to NO in many organs. Recently, NO has been suggested as a second messenger molecule for the stimulatory effect of insulin in carbohydrate metabolism (Khan et al., 2000; Khan and Sinha, 1990). Incubation of various tissues, including heart, liver, kidney, muscle and intestine from mice and erythrocytes or their membrane fractions from humans, with physiologic concentration of insulin results in the activation of a membrane-bound NOS and synthesis of No. Furthermore, NO has shown an insulin-like effect in stimulating glucose transport and glucose oxidation in muscle, a major site for insulin action that is completely blocked in the presence of NOS inhibitor. Also, injection of NO in alloxan-induced diabetic mice mimics the effect of insulin in the control of hyperglycemia by lowering plasma glucose concentration (Kahn et al., 2000; Czech, 1990; Khan and Sinha, 1990). There is also evidence that a combination of tumor necrosis factor a, interferon γ and interleukin 1β plus lipopolysaccharide in hepatic cultures result in an induction of NOS and concomitant inhibition of hepatic gluconeogenesis and glycogenolysis (Ceppi and Titheradge, 1998; Casado et al., 1996). Similarly, the hypoglycemia that is seen in septic shock, is the result of reduced liver glucose output while the peripheral glucose utilisation is enhanced (Ceppi et al., 1989). In addition, it has been reported that culturing hepatic parenchymal cells in the presence of insulin results in an NO-dependent inhibition of the glucagon-stimulated rate of glucose production via gluconeogenesis plus glycogenolysis (Stadler et al., 1995). NO can inhibit both the basal and the glucagon-and cyclic AMP-mediated mobilization of glycogen (Ceppi and Titheradge, 1998). Also NO is shown to stimulate glycolysis through a glycogenolysis-independent mechanism and a concomitant increase of glycogen possibly through inhibition of glycogenolysis (Cidad et al., 2004). These mentioned mechanisms all explain the inhibitory effect of sildenafil on glycogenolysis and also its blood glucose lowering effect.
On the other hand, a role for reactive oxygen intermediates in the regulation of hepatic glucose production has been reported (Ceppi and Titheradge, 1998). Troglitazone inhibits expression of the PEPCK gene in isolated hepatocytes by an antioxidant property due to existence of the alpha-tocopherol (vitamin E) moiety in its chemical structure (Davies et al., 2001). The antidiabetic drug metformin which acts through inhibition of hepatic gluconeogenesis produces concurrent antioxidant effect that is beneficial in the treatment of diabetes (Cosic et al., 2001). Recent studies indicated a protective role for cGMP from induction of oxidative stress inside cells (Aghababaeian et al., 2005; Radfar et al., 2005; Astaneie et al., 2005; Abdollahi et al., 2004b; Jahanshahi et al., 2004; Abdollahi et al., 2003c). It has been reported that sildenafil, through the inhibition of PDE5 and augmentation of inhibitory action of the NO-cGMP axis on NADPH oxidase expression and activity inhibits superoxide formation in corpus cavernosal smooth muscle cells (Koupparis et al., 2005; Shukla et al., 2005). Thus antioxidant potential of sildenafil might be another mechanism for its glucose lowering effect and inhibition of glycogenolysis.
Taken collectively, it is concluded that sildenafil reduces glycogenolysis which in turn lowers blood glucose concentration especially at higher doses. This effect of sildenafil seems to be consistene with its NO mimicking potential and antioxidant properties. Additional studies are required to determine whether the same effects could be seen in humans.
ACKNOWLEDGMENTS
This study was supported by a grant form Toxicology Pivot, Tehran University of Medical Sciences.
REFERENCES
- Abdollahi, M. and B. Simaiee, 2003. Stimulation by theophylline and sildenafil of rat submandibular secretion of protein, epidermal growth factor and flow rate. Pharmacol. Res., 48: 445-449.
CrossRefDirect Link - Abdollahi, M., T.S. Chan, V. Subrahmanyam and P.J. O'Brien, 2003. Effects of phosphodiesterase 3,4,5 inhibitors on hepatocyte cAMP levels, glycogenolysis, gluconeogenesis and susceptibility to a mitochondrial toxin. Mol. Cell Biochem., 252: 205-211.
PubMedDirect Link - Abdollahi, M., M. Donyavi, S. Pournourmohammadi and M. Saadat, 2004. Hyperglycemia associated with increased hepatic glycogen phosphorylase and phosphoenolpyruvate carboxykinase in rats following subchronic exposure to malathion. Comp. Biochem. Physiol. Part C, 137: 343-347.
Direct Link - Abdollahi, M., A. Ranjbar, S. Shadnia, S. Nikfar and A. Rezaie, 2004. Pesticides and oxidative stress: A review. Med. Sci. Monit., 10: RA141-RA147.
PubMedDirect Link - Aghababaeian, R., M. Ghazi-Khansari, K. Abdi, F. Taghadosinejad and M. Abdollahi, 2005. Protective effects of sildenafil and dipyridamol from lead-induced lipid peroxidation in perfused rat liver. Int. J. Pharmacol., 1: 157-160.
CrossRefDirect Link - Aydin, A., H. Orhan, A. Sayal, M. Ozata, D. Sahin and A. Isimer, 2001. Oxidative stress and nitric oxide related parameters in type II diabetes mellitus: Effects of glycemic control. Clin. Biochem., 34: 65-70.
Direct Link - Ballard, S.A., C.J. Gingell, K. Tang, L.A. Turner, M.E. Price and A.M. Naylor, 1998. Effects of sildenafil on the relaxation of human corpus cavernosum tissue in vitro and on the activities of cyclic nucleotide phosphodiesterase isozymes. J. Urol., 159: 2164-2171.
Direct Link - Barford, D. and L.N. Johnson, 1989. The allosteric transition of glycogen phosphorylase. Nature, 340: 609-616.
CrossRefDirect Link - Bollen, M., S. Keppens and W. Stalmans, 1998. Specific features of glycogen metabolism in the liver. Biochem. J., 336: 19-31.
Direct Link - Casado, M., M.J. Diaz-Guerra, L. Bosca and P. Martin-Sanz, 1997. Differential regulation of nitric oxide synthase mRNA xpression by lipopolysaccharide and pro-inflammatory cytokines in fetal hepatocytes treated with cycloheximide. Biochem. J., 327: 819-823.
PubMedDirect Link - Ceppi, E.D. and M.A. Titheradge, 1998. The importance of nitric oxide in the cytokine-induced inhibition of glucose formation by cultured hepatocytes incubated with insulin, dexamethasone and glucagon. Arch. Biochem. Biophys., 349: 167-174.
PubMedDirect Link - Chang, H.C. and M.D. Lane, 1966. The enzymatic carboxylation of phosphoenolpyruvate. II. Purification and properties of liver mitochondrial phosphoenolpyruvate carboxykinase. J. Biol. Chem., 241: 2413-2420.
PubMedDirect Link - Cidad, P., A. Almeida and J.P. Bolanos, 2004. Inhibition of mitochondrial respiration by nitric oxide rapidly stimulates cytoprotective GLUT3-mediated glucose uptake through 5'-AMP-activated protein kinase. Biochem. J., 384: 629-636.
PubMedDirect Link - Cosic, V., S. Antic, M. Pesic, O. Jovanovic, S. Kundalic and V.B. Djordjevic, 2001. Monotherapy with metformin: Does it improve hypoxia in type 2 diabetic patients? Clin. Chem. Lab. Med., 39: 818-821.
PubMedDirect Link - Dal-Piaz, V. and M.P. Giovannoni, 2000. Phosphodiesterase 4 inhibitors, structurally unrelated to rolipram, as promising agents for the treatment of asthma and other pathologies. Eur. J. Med. Chem., 35: 463-480.
PubMedDirect Link - Davies, G.F., R.L. Khandelwal, L. Wu, B.H. Juurlink and W.J. Roesler, 2001. Inhibition of phosphoenolpyruvate carboxykinase (PEPCK) gene expression by troglitazone: A peroxisome proliferator-activated receptor-gamma (PPARgamma)-independent, antioxidant-related mechanism. Biochem. Pharmacol., 62: 1071-1079.
PubMedDirect Link - Foufelle, F. And P. Ferre, 2002. New perspectives in the regulation of hepatic glycolytic and lipogenic genes by insulin and glucose: A role for the transcription factor sterol regulatory element binding protein-1c. Biochem. J., 366: 377-391.
Direct Link - Jahanshahi, G., V. Motavasel, A. Rezaie, A.A. Hashtroudi, N.E. Daryani and M. Abdollahi, 2004. Alterations in antioxidant power and levels of epidermal growth factor and nitric oxide in saliva of patients with inflammatory bowel diseases. Dig. Dis. Sci., 49: 1752-1757.
CrossRefPubMedDirect Link - Kahn, N.N., K. Acharya, S. Bhattacharya, R. Acharya, S. Mazumder, W.A. Bauman and A.K. Sinha, 2000. Nitric oxide: The second messenger of insulin. IUBMB Life, 49: 441-450.
Direct Link - Khan, N.N. and A.K. Sinha, 1990. Stimulation of prostaglandin E1 binding to human platelet membrane by insulin and the activation of adenylate cyclase. J. Biol. Chem., 265: 4976-4981.
PubMedDirect Link - Koupparis, A.J., J.Y. Jeremy, S. Muzaffar, R. Persad and N. Shukla, 2005. Sildenafil inhibits the formation of superoxide and the expression of gp47 NAD(P)H oxidase induced by the thromboxane A2 mimetic, U46619, in corpus cavernosal smooth muscle cells. BJU Int., 96: 423-427.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Milani, E., S. Nikfar, R. Khorasani, M.J. Zamani and M. Abdollahi, 2005. Reduction of diabetes-induced oxidative stress by phosphodiesterase inhibitors in rats. Comp. Biochem. Physiol. C. Toxicol. Pharmacol., 140: 251-255.
PubMedDirect Link - Nordlie, R.C., J.D. Foster and A.J. Lange, 1999. Regulation of glucose production by the liver. Ann. Rev. Nutr., 19: 379-406.
PubMed - Parker, J.C., M.A. van Volkenburg, N.A. Nardone, D.M. Hargrove and K.M. Andrews, 1997. Modulation of insulin secretion and glycemia by selective inhibition of cyclic AMP phosphodiesterase III. Biochem. Biophys. Res. Commun., 236: 665-669.
CrossRefDirect Link - Pointer, R.H., F.R. Butcher and J.N. Fain, 1976. Studies on the role of cyclic guanosine 3':5'-monophosphate and extracellular Ca2+ in the regulation of glycogenolysis in rat liver cells. J. Biol. Chem., 251: 2987-2992.
Direct Link - Radfar, M., B. Larijani, M. Hadjibabaie, B. Rajabipour, A. Mojtahedi and M. Abdollahi, 2005. Effects of pentoxifylline on oxidative stress and levels of EGF and NO in blood of diabetic type-2 patients: A randomized, double-blind placebo-controlled clinical trial. Biomed. Pharmacother., 59: 302-306.
Direct Link - Rahimi, R., S. Nikfar, B. Larijani and M. Abdollahi, 2005. A review on the role of antioxidants in the management of diabetes and its complications. Biomed. Pharmacother., 59: 365-373.
PubMedDirect Link - Saadat, M., S. Pournourmohammadi, M. Donyavi, R. Khorasani, G. Amin, A.N. Salehnia and M. Abdollahi, 2004. Alteration of rat hepatic glycogen phosphorylase and phosphoenolpyruvate carboxykinase activities by Satureja khuzestanica Jamzad essential oil. J. Pharm. Pharm. Sci., 7: 310-314.
Direct Link - Soderling, S.H. and J.A. Beavo, 2000. Regulation of cAMP and cGMP signaling: New phosphodiesterases and new functions. Curr. Opin. Cell Biol. Rev., 12: 174-179.
Direct Link - Souness, J.E., D. Aldous and C. Sargent, 2000. Immunosuppressive and anti-inflammatory effects of cyclic AMP phosphodiesterase (PDE) type 4 inhibitors. Immunopharmacol. Rev., 47: 127-162.
CrossRefDirect Link - Stadler, J., D. Barton, H. Beil-Moeller, S. Diekmann, C. Hierholzer, W. Erhard and C.D. Heidecke, 1995. Hepatocyte nitric oxide biosynthesis inhibits glucose output and competes with urea synthesis for L-arginine. Am. J. Physiol., 268: G183-188.
Direct Link - Astaneie, F., M. Afshari, A. Mojtahedi, S. Mostafalou, M.J. Zamani, B. Larijani and M. Abdollahi, 2005. Total antioxidant capacity and levels of epidermal growth factor and nitric oxide in blood and saliva of insulin-dependent diabetic patients. Arch. Med. Res., 36: 376-381.
CrossRefPubMedDirect Link - Abdollahi, M., F. Fooladian, B. Emami, K. Zafari and A. Bahreini‐Moghadam, 2003. Protection by sildenafil and theophylline of lead acetate‐induced oxidative stress in rat submandibular gland and saliva. Hum. Exp. Toxicol., 22: 587-592.
CrossRefDirect Link