
Z. A. Zakaria
School of Biotechnology and Life Sciences and Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
H. M. Khairi
Department of Biomedical Sciences, Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M. N. Somchit
Department of Biomedical Sciences, Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M. R. Sulaiman
Department of Biomedical Sciences, Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
A. M. Mat Jais
Department of Biomedical Sciences, Faculty of Medicine and Health Science, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
I. Reezal
Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
N. N. Mat Zaid
School of Biotechnology and Life Sciences and Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
S.N.Z. Abdul Wahab
School of Biotechnology and Life Sciences and 3Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
N. S. Fadzil
School of Biotechnology and Life Sciences and 3Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
M. Abdullah
School of Biotechnology and Life Sciences and 3Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
C. A. Fatimah
School of Biotechnology and Life Sciences and 3Medical Science Industry, Universiti Industri Selangor, Jalan Zirkon ATA, Seksyen 7, 40000 Shah Alam, Selangor, Malaysia
International Journal of Pharmacology
Year: 2006 | Volume: 2 | Issue: 2 | Page No.: 216-220







ABSTRACT
To determine on the antibacterial activity of the leaves and acute toxicity level of the leaves, tender leaves and rhizomes of Manihot esculenta var. Sri Pontian extracts. The chloroform (CME1) and ethanol (EME1) leaves extracts of M. esculenta (25, 50 and 100% concentrations) were tested against a selected groups of Gram positive and Gram negative bacteria using the disc diffusion method. On the other hand, the chloroform and ethanol leaves (CME2 and EME2), as well as the tender young leaves (CME3 and EME3) and rhizomes (CME4 and EME4), extracts of M. esculenta, (concentration ranging from 200 to 2600 ppm) were tested for their chronic toxicity level using the brine shrimp bioassay. CME1 was found to give positive antibacterial activity against L. monocytogenes, Vibrio cholerae, Shigella flexneri and Salmonella typhi while EME1 was effective against P. aeroginosa, C. diphtheria and V. cholerae. The results also showed that among the chloroform extracts, CME4 (LC50) = 413.9±51.6) possessed significantly (p<0.05) high toxicity followed by CME3 (LC50 = 496.2±33.1) and CME2 (LC50 = 532.9±22.9) while among the ethanol extracts, EME3 (LC50 = 344.7±33.9) was significantly (p<0.05) more toxic followed by EME2 (LC50 = 534.3±81.5) and EME4 (LC50 = 609.6±74.8). Overall, EME3 and CME4 were highly toxic than their counterpart (CME3 and EME4), respectively, while CME2 and EME2 did no show any discrepancy in their LD50 value. M. esculenta possess an antibacterial property and low toxicity level.
PDF Abstract XML References Citation
How to cite this article
Z. A. Zakaria, H. M. Khairi, M. N. Somchit, M. R. Sulaiman, A. M. Mat Jais, I. Reezal, N. N. Mat Zaid, S.N.Z. Abdul Wahab, N. S. Fadzil, M. Abdullah and C. A. Fatimah, 2006. The in vitro Antibacterial Activity and Brine Shrimp Toxicity of Manihot esculenta var. Sri Pontian (Euphorbiacea) Extracts. International Journal of Pharmacology, 2: 216-220.
DOI: 10.3923/ijp.2006.216.220
URL: https://scialert.net/abstract/?doi=ijp.2006.216.220
DOI: 10.3923/ijp.2006.216.220
URL: https://scialert.net/abstract/?doi=ijp.2006.216.220
INTRODUCTION
Manihot esculenta, commonly known as tapioca or cassava and native to South America (Olsen and Schaal, 1999) and widely distributed in the Southeast Asia region, is a plant belonging to the family Euphorbiacea. It is a tropical perennial shrub and is grown for its enlarged starch-filled tuberous roots (El-Sharkawy, 2004). It is classified under roots and tubers, which is the class of foods that basically provide energy in the human diet in the form of carbohydrates (Alfonso et al., 2004). Other than that, the young leaves are used as vegetable (Lancaster and Brooks, 1983) and contain a high amount of protein (Ravindran, 1993), as well as vitamin A and C.
There is a bitter, poisonous and a sweet, nonpoisonous-variety of M. esculenta (Essers, 1995). The poisoning effect of M. esculenta is attributed to the presence of hydrocyanic acid (HCN), a type of glycosides, found in all parts of the bitter, poisonous variety, but presence only in the skin of sweet, nonpoisonous variety of M. esculenta (Vetter, 2000). These glycosides can be removed or destroyed by peeling the skin of the sweet variety roots and boiling the bitter variety in water, respectively (Balagopalan and Rajalakshmy, 1998). The peeled roots of the sweet variety are usually eaten cooked or baked. The bitter ones are grated, mixed with water and pressed to extract the juice. The paste of cassava is baked into pancake-like bread while the extracted juice is fermented into strong liquor called kasiri. The juice can also be concentrated and sweetened until it becomes dark viscous syrup called kasripo (casareep). This syrup has antiseptic properties and is used for flavoring.
Traditionally, the medicinal folklore used of M. esculenta is attributed to its roots/rhizomes. In Suriname's traditional medicine, the Amerindians use the brown juice, obtained during processing, for burns. Bark decoction of the tuber of M. esculenta when drunk is said to be antihelmintic. According to Hartwell (1967-1971), M. esculenta is used in folk remedies for cancerous affections, condylomata, excrescences of the eye and tumors. Other than being reported to be antiseptic, cyanogenetic, demulcent and diuretic, M. esculenta is also a folk remedy for abscesses, boils, conjunctivitis, diarrhea, dysentery, flu, hernia, inflammation, marasmus, prostatitis, snakebite, sore, spasm and swellings (Duke and Wain, 1981). Furthermore, M. esculenta is indigenously known, to most Ugandans, to have more value as medicine for various diseases such as asthma, coughs, wounds, tuberculosis (TB), measles, backache, chest pain, skin rash, sweating, stomachache, headache and medicinal anesthesia. However, there are no scientifically proven studies reported on the medicinal properties of this plant.
Based on the folklore medicinal believes mentioned above and the fact that there is no well-documented medicinal uses for M. esculenta, the objectives of the current study are to determine for the presence of antibacterial activity in the leaves and to elucidate on the chronic toxicity level in the leaves, tender young leaves and roots/rhizomes of chloroform and ethanol extracts of M. esculenta.
MATERIALS AND METHODS
Materials: M. esculenta var. Sri Pontian leaves, tender young leaves and roots were supplied by Malaysian Agriculture Research and Development Institute Serdang, Selangor, Malaysia and a voucher specimen was deposited at the Herbarium of MARDI, Serdang, Selangor, Malaysia.
Microorganisms tested in this study were Streptococcus pneumoniae, Bacillus cereus, Listeria monocytogenes, Pseudomonas aeruginosa, Staphylococcus aureus, Corneybacterium diphtheria, Enterococcus faecalis, Klebsiella pneumoniae, Shigella flexneri, Proteus vulgaris, Vibrio cholerae, Escherichia coli, Vibrio parahemolyticus and Salmonella typhi.
Preparation of Manihot esculanta extracts: Fresh leaves of M. esculenta were oven-dried for 24 h at 40°C according to the methods described by Somchit et al. (2003) but with slight modifications. It was then ground into small pieces under sterilized condition and extracted separately with chloroform and ethanol in the ratio of 1:20 (w/v) for 24 h by using Soxhlet apparatus. The aqueous extract obtained was kept at -80°C for 48 h and then freeze-dried for 72 h while the resultant extraction of chloroform and ethanol was completely evaporated by using rotary evaporator machine (Somchit et al., 2003). The obtained dried crude extracts of chloroform (CEME) and ethanol (EEME), dissolved in dimethyl sulfoxide (DMSO) in the ratio of 1:10 (w/v) and considered as stock solutions with 100% concentration/strength, were then diluted using DMSO to the concentrations of 25 and 50% and used in the antibacterial studies. The same dried crude extracts of CEME and EEME, prepared on another occasion, were diluted using DMSO to the concentrations ranging from 200 to 2600 ppm and used in the toxicity studies.
Antibacterial study: The above-mentioned bacteria were incubated at 37°C±0.5 for 24 h after injection into nutrient broth. Mueller Hinton Agar (MHA) (Oxoid, UK), sterilized in a flask and cooled to 40-50°C, was poured (15 mL) into sterilized Petri dishes (diameter of 9 cm) and allowed to harden under room temperature. This is followed by homogenous distribution of 0.1 mL bacteria cultures (106 bacteria per mL) onto medium in Petri dishes.
Twenty microliter of the respective extract were then loaded into empty sterilized blank discs (6 mm diameter, Oxoid, UK) and dried under room temperature. Dried discs loaded with extracts were then positioned on the solid agar medium by pressing slightly (Sundar, 1996). Petri dishes were placed in incubator according to their respective growth temperature and condition for 18 to 24 h. At the end of the period, inhibition zones formed was measured in mm. The study was performed in triplicate and the formation of the inhibition zones were compared with those of antibiotic discs. In addition, commercial antibiotic discs (Chloramphenicol; 30 μg μL-1) were used for comparison.
Chronic toxicity study: The chronic toxicity level of leaves, tender young leaves and rhizomes of M. esculenta were determined using the brine shrimps (Artemia salina) bioassay described previously by Lieberman (1999). Briefly, brine shrimps were used at hatching stage (nauplii), after 46 h incubation of its eggs in a petri dish filled with sea salt water. Ten naupliis were placed in each vial containing the CEME or EEME extracts, at their respective concentration (ranging from 200 to 2600 ppm), an observed for 24 h. The brine shrimp are best seen against a dark background with good light from the side of the tube. Normally, most of the shrimp swim vigorously with their swimming rate, at room temperature, of several beats per second and show a phototaxis activity. Shrimp that lie motionless at the bottom of the tube are dead. The most common sub-lethal effect is impairment of swimming rate. For each tube, the fraction of shrimp that are dead was counted. LD50 values were obtained from the data according to the Reed-Muench method (Reed and Muench, 1938) by plotting data of the percentage of mortality rate against the respective dosage, plotted to a logarithmic scale, in each test tube. Each experiment was carried out with three replications, two sets of controls and was repeated five times. Statistical analysis was by Sigmastat 2.0 (Jandel Scientific Software, San Rafael, CA, USA; copyright 1995).
RESULTS
As can be seen from the Table 1, the CEMC was less effective when compared to AEMC and MEMC, in term of number of bacteria inhibited. At the concentration ranging from 10,000 to 50,000 ppm, the AEMC was effective against S. aureus and K. rhizophila while the CEMC was effective against the two bacteria, as well as B. cereus, P. vulgaris, S. flexneri and A. hydrophila. At the concentration ranging from 40,000 to 100,000 ppm, the AEMC was effective against C. diphtheriae, P. vulgaris, S. epidermidis and A. hydrophila; the MEMC was effective against C. diphtheriae and S. typhi and the CEMC was only effective against S. aureus. Interestingly, the MEMC was also found to be effective against E. coli, which is observed at the concentration of 70,000 ppm and above.
Our further studies to evaluate the level of chronic toxicity of both extracts of the leaves, young tender leaves and rhizomes of M. esculenta have demonstrated that all parts of M. esculenta possessed certain level of toxicity, which is considered very high when assessed using the brine shrimp bioassay (Table 2-4). This is indicated by the low level of concentration of each part of the plant that is needed to cause toxicity to half of the population of Artemia salina.
| Table 1: | The Antibacterial Activity of Aqueous, Methanol and Chloroform Extracts of Muntingia calabura Determined by Disc Diffusion Method |
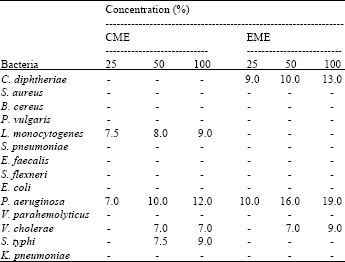 | |
| IZ = Inhibition zone (mm), Except for P. vulgaris (IZ = 9 mm), Chloramphenicol gave inhibition zone of ≤20 mm against all bacteria | |
| Table 2: | Acute LD50 of ethanol and chloroform extracts of M. esculenta leaves after 24 h exposure to Artemia salina assessed using the Reed-Muench method |
| Table 3: | Acute LD50 of ethanol and chloroform extracts of M. esculenta young tender leaves after 24 h exposure to Artemia salina assessed using the Reed-Muench method |
| a,bData with different asterisks differed significantly (p<0.05) | |
| Table 4: | Acute LD50 of ethanol and chloroform extracts of M. esculenta rhizomes after 24 h exposure to Artemia salina assessed using the Reed-Muench method |
| a,bData with different asterisks differed significantly (p<0.05) | |
In our studies, the chloroform and ethanol extracts of M. esculenta leaves (CME2 and EME2), as well as the tender leaves (CME3 and EME3) and rhizomes (CME4 and EME4), prepared in the concentration ranging from 200 to 2600 ppm, were found to possess different level of median lethal concentration (LC50) as described below. The results demonstrated that among the chloroform extracts, CME4 (median lethal dose (LC50) = 413.9±51.6) possessed significantly (p<0.05) higher toxicity level followed by CME3 (LC50 = 496.2±33.1) and CME2 (LC50 = 532.9±22.9) while among the ethanol extracts, EME3 (LC50 = 344.7± 33.9) was significantly (p<0.05) more toxic followed by EME2 (LC50 = 534.3±81.5) and EME4 (LC50 = 609.6±74.8). Overall, EME3 and CME4 were highly toxic than their counterpart (CME3 and EME4), respectively, while CME2 and EME2 did no show any discrepancy in their LC50 value.
DISCUSSION
M. esculenta var. Sri Pontian, locally known to the Malays as ‘ubi kayu’, is grown basically as an energy provider for human in the form of carbohydrates due to its enlarged starch-filled tuberous roots (El-Sharkawy, 2004). Other than that, its leaves and young tender leaves are used as vegetable in many part of the world (El-Sharkawy, 2004), including Malaysia. In Malaysia, particularly, M. esculenta is traditionally used in folklore medicine to treat various ailments such as headache, colds and fever and to treats gastric constipation.
However, there are many studies reported on its toxic effect, which is greatly attributed to the presence of free and bound cyanogenic glycosides, linamarin and lotaustralin (Frakes et al., 1985; Elias et al., 1997; Nambisan, 1999; Andersen et al., 2000). These compounds are converted to HCN in the presence of linamarase, a naturally occurring enzyme in M. esculenta (Nambisan, 1999). Linamarase acts on the glycosides when the cells are ruptured. All plant parts contain cyanogenic glycosides with the leaves having the highest concentrations. In the roots, the peel has a higher concentration than the interior. In cases of human malnutrition, where the diet lacks protein and iodine, under processed roots of high HCN cultivars may result in serious health problems (Tylleskar et al., 1992). According to World Health Organization (1993), the toxicity of cyanogenic plants depends primarily on the potential concentration of HCN that may be released upon consumption. If the cyanogenic plant is inadequately detoxified during processing or preparation of the food, the potential HCN concentration, which may be released, can still be high. Upon consumption of a cyanogenic plants, β-glycosidase will be released during digestion and remain active until deactivated by the low pH of the stomach (Thayer and Conn, 1981; Poulton, 1990). This enzyme will hydrolyze the cynogenic glycosides and release, at least, the potential HCN content of the plant (Thayer and Conn, 1981; Poulton, 1990) Cyanide exerts its toxic effects by binding to the ferric ion of cytochrome oxidase, an enzyme that accounts for about 90% of the total oxygen uptake in most cells via the electron transport chain (Friedman, 1980). Inhibition of cytochrome oxidase thus virtually completely disrupts cellular oxygen utilization resulting in cytotoxic hypoxia, disfunctioning and death (Friedman, 1980, Abu-Bakare et al., 1986; Gruhnert et al., 1994). This situation leads to the exposure of humans with high diets of cassava and cassava products and of human engaged in the cassava product industries to varying levels of cyanide poisoning resulting in chronic and acute cyanide toxicity (Tylleskar et al., 1992; Abu-Bakare et al., 1986; Akanji et al., 1990; Osuntokun, 1981; Bennet et al., 1987; Howlett et al., 1990).
The present study has demonstrated the potential used of M. esculenta as an antibacterial agent against the infection of some of the bacteria used. The chloroform and ethanol extracts of M. esculenta leaves were found to be effective in the treatment against gram positive (C. diphtheria, L. monocytogenes, P. aeroginosa) and gram negative (Vibrio cholerae, Shigella flexneri and Salmonella typhi) bacteria. Present study also demonstrated that both extracts were only effective against six out of fourteen bacteria used, which might help explained the fact that the Malays folklore medicinal used of M. esculenta did not include its used in treatment against bacteria infection. However, this does not exclude the fact that M. esculenta did possess antimicrobial activity. Although isolation and identification of the bioactive compounds responsible for the observed activity is not the main objective of this study, the activity might be attributed to the presence of bioactive compounds, such as tannins and flavonoids (Diaz et al., 1988), glycosides (Chukwurah and Ajali, 2000) and saponins (Pretorius et al., 2003). The presence of tannins, flavonoids and glycosides (Ogunleye and Ibitoye, 2003), which have been known to occur abundantly in leaves of all plants, was expected to play major role in the observed antibacterial activity of M. esculenta extracts.
Further study using the brine shrimps bioassay has demonstrated that the leaves, young tender leaves and rhizomes of M. esculenta possessed chronic toxicity effect that is different among them and did not depend on the type of solvents used in their extraction. The chloroform and ethanol extracts of M. esculenta leaves did not show any discrepancies in their LC50, while the ethanol extract of its tender young leaves and chloroform extract of its rhizomes exhibited high toxicity when compared to their counterparts, respectively. When compared among the chloroform extracts, the rhizomes was significantly more toxic followed by the tender young leaves and the leaves, while comparison among the ethanol extracts showed in sequence that the tender young leaves was much more toxic than the leaves and the rhizomes.
Based on the results obtained, it is plausible to suggest that M. esculenta leaves do possess certain level of antimicrobial activity, depending on the type of bacteria used. Furthermore, its leaves, tender young leaves and rhizomes do possess different level of toxicity, which is due to the presence of various types of glycosides that are converted to HCN. The toxicity level of each part of M. esculenta tested is considered higher since they gave the LD50 concentration below 1000 ppm, respectively.
ACKNOWLEDGMENT
The authors would like to thank Universiti Industri Selangor for the research grant (Project Code No. 03013; Vote No. 3090103013) and Universiti Putra Malaysia for the facilities.
REFERENCES
- Abu-Bakare, A., R. Taylor and K.G.M. Alberti, 1986. Tropical or malnutrition-related diabetes: A real syndrome. Lancet, 1: 1135-1138.
Direct Link - Akanji, A.O., I. Adeyefa, M. Charles-Davies and B.O. Osotimehin, 1990. Plasma glucose and thiocyanate response to different mixed cassava meals in non-diabetic Nigerians. Eur. J. Clin. Nutr., 44: 71-77.
PubMedDirect Link - Alfonso, L.C., Z. Fundora-Mayor, S. Pico and E. Salinas, 2004. The use of home Gardens as a component of the national strategy for the in situ conservation of plant genetic resources in cuba. PGR Newslett., 123: 9-18.
Direct Link - Andersen, M.D., P.K. Busk, I. Svendsen and B.L. Moller, 2000. Cytochromes P-450 from cassava (manihot esculenta crantz) catalyzing the first steps in the biosynthesis of the cyanogenic glucosides linamarin and lotaustralin. Cloning, functional expression in Pichia pastoris and substrate specificity of the isolated recombinant enzymes. J. Biol. Chem., 275: 1966-1975.
Direct Link - Balagopalan, C. and L. Rajalakshmy, 1998. Cyanogen accumulation in environment during processing of cassava (Manihot esculenta crantz) for starch and sago. Water Air Soil Pollut., 102: 407-413.
CrossRefDirect Link - Chukwurah, B.K.C. and U. Ajali, 2000. Antimicrobial investigation of the constituents of the methanol extract of the rhizomes of Anchomanes difformis, ENGL. Indian J. Pharm. Sci., 62: 296-299.
Direct Link - El-Sharkawy, M.A., 2004. Cassava biology and physiology. Plant Mol. Biol., 56: 481-501.
PubMedDirect Link - Elias, M., B. Nambisan and P.R. Sudhakaran, 1997. Catabolism of linamarin in cassava (Manihot esculenta Crantz). Plant Sci., 126: 155-162.
Direct Link - Frakes, R.A., R.P. Sharma and C.C. Willhite, 1985. Developmental toxicity of the cyanogenic glycoside linamarin in the golden hamster. Teratology, 31: 241-246.
PubMedDirect Link - Howlett, W.P., G.R. Brubaker, N. Mlingi and H. Rosling, 1990. Konzo an upper motor neurone disease studied in Tanzania. Brain, 113: 223-235.
Direct Link - Lancaster, P.A. and J.E. Brooks, 1983. Cassava leaves as human food. Econ. Bot., 37: 331-348.
CrossRefDirect Link - Lieberman, M., 1999. A brine shrimp bioassay for measuring toxicity and remediation of chemicals. J. Chem. Educ., 76: 1689-1690.
Direct Link - Nambisan, B., 1999. Cassava latex as a source of linamarase for determination of linamarin. J. Agric. Food. Chem., 47: 372-373.
CrossRefDirect Link - Ogunleye, D.S. and S.F. Ibitoye, 2003. Studies of antimicrobial activity and chemical constituents of Ximenia Americana. Trop. J. Pharmacol. Res., 2: 239-241.
Direct Link - Olsen, K.M. and B.A. Schaal, 1999. Evidence on the origin of cassava: Phylogeography of Manihot esculenta. Proc. Natl. Acad. Sci. USA., 96: 5586-5591.
Direct Link - Osuntokun, B.O., 1981. Cassava diet, chronic cyanide intoxication and neuropathy in the Nigerian Africans. World Rev. Nutr. Diet., 36: 141-173.
PubMedDirect Link - Ravindran, V., 1993. Cassava leaves as animal feed: Potential and limitations. J. Sci. Food Agric., 61: 141-150.
Direct Link - Somchit, M.N., I. Reezal, I.E. Nur and A.R. Mutalib, 2003. In vitro antibacterial activity of ethanol and water extract of Cassia alata. J. Ethnopharm., 84: 1-4.
CrossRef - Sundar, R.K., 1996. Antibacterial activity of some medicinal plants of Papua New Guinea. Int. J. Pharmacognosy, 34: 223-225.
Direct Link - Thayer, S. and E. Conn, 1981. Subcellular localization of dhurrin β-glucosidase and hydroxynitrile lyase in the mesophyll cells of sorghum leaf blades. Plant Physiol., 67: 617-622.
Direct Link - Tylleskar, T., M. Banea, N. Bikangi, R.D. Cooke, N.H. Poulter and H. Rosling, 1992. Cassava cyanogens and konzo, an upper motoneuron disease found in Africa. Lancet, 339: 208-211.
CrossRefPubMedDirect Link