
H.H. Azzaz
Department of Dairy Science, National Research Centre, 33 El-Buhouth Street, Dokki, Giza, Egypt
LiveDNA: 20.630
A.A. Aboamer
Department of Dairy Science, National Research Centre, 33 El-Buhouth Street, Dokki, Giza, Egypt
LiveDNA: 20.20856
Hoda Alzahar
Department of Dairy Science, National Research Centre, 33 El-Buhouth Street, Dokki, Giza, Egypt
Noha A. Hassaan
Department of Animal Production, National Research Centre, 33 El-Buhouth Street, Dokki, Giza, Egypt
H.A. Murad
Department of Dairy Science, National Research Centre, 33 El-Buhouth Street, Dokki, Giza, Egypt
International Journal of Dairy Science
Year: 2020 | Volume: 15 | Issue: 1 | Page No.: 48-53







ABSTRACT
Background and Objectives: The search for cheap-safe tools for promoting farm animal productivity is the main concern of the animal nutritionists’ nowadays. The main objective of this study was to evaluate impact of cellulase new formula for promoting performance of lactating Baladi goats. Materials and Methods: Eighteen early lactating Baladi goats were randomly divided into 3 groups; the 1st group was fed control ration (50% concentrate feed mixture (CFM), 25% berseem (clover) hay and 25% wheat straw), the 2nd fed control ration+LAB produced cellulase at 42.16 IU kg–1 DM (R1), while the 3rd fed control ration+42.16 IU kg–1 DM of Pan-Zyme® (R2). The goats were fed dry matter according to 4% of their body weight for 9 weeks. Results: Goats fed cellulases supplemented rations (R1 and R2) showed higher (p<0.05) nutrients digestibility coefficients, milk yield and milk fat, protein, lactose and total solids yields than those of control. Cellulases supplementation did not cause any change in all of blood parameters (ex; glucose, protein AST, ALT and urea concentrations), milk fatty acids and amino acids profiles. Conclusion: Inclusion of the produced cellulase in lactating goat’s rations improved their ability for feed utilization and milk production with no bad effects on liver and kidney functions, which reflecting the safety and efficacy of the new cellulase product.
PDF Abstract XML References Citation
Received: November 10, 2019;
Accepted: December 08, 2019;
Published: February 01, 2020
Copyright: © 2020. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
H.H. Azzaz, A.A. Aboamer, Hoda Alzahar, Noha A. Hassaan and H.A. Murad, 2020. Effect of Cellulases Supplementation on Milk Yield and Feed Utilization by Baladi Goats in Early Lactation. International Journal of Dairy Science, 15: 48-53.
DOI: 10.3923/ijds.2020.48.53
URL: https://scialert.net/abstract/?doi=ijds.2020.48.53
DOI: 10.3923/ijds.2020.48.53
URL: https://scialert.net/abstract/?doi=ijds.2020.48.53
INTRODUCTION
Growing consumer’s concern with use of growth promoters (ex: ionophores)1 in livestock feeding, has forced the nutritionists for searching for safe tools for promoting farm animal productivity2. The natural feed additives like probiotics3-5, medicinal plants6, essential oils7 and feed enzymes8-11 are considered as promising tools for animal’s performance enhancement. In recent years, feed enzymes have considerable attention as it considered as safe and cost-effective means for improving efficiency of feed utilization12. Most of feed enzymes are belonging to hydrolytic enzymes (ex: proteolytic, lipolytic and fibrolytic enzymes) which act for add water across a chemical bond13. The fibrolytic enzymes are large family of enzymes that act for hydrolysis of polysaccharides of plant cell walls (mainly cellulose, hemicellulose and pectin). Cellulase, hemicellulase (xylanase) and pectinase are the main representatives of these enzymes8-11. Cellulase as a multi enzyme system act collectively to break down of plant cellulose to simpler units of glucose14. The most common sources of cellulase are the microbial cellulase, as many of bacteria (ex: lactic acid bacteria) and fungi (ex: Aspergillus, Trichoderma and Penicillium genera) have the ability to produce it for the commercial usage15. The activity of the produced cellulase can be diverse depending on the microbial strain, the substrates and the cultivation conditions used16. Nowadays, cellulases utilization has been increased due to continuous increase of grains prices, which forced the farmers for depending more on forages and agricultural residues for feeding of their animals17. Positive effects of cellulases on digestibility of high fibrous diets in vitro and in vivo has been reported10,12,15,17. Exogenous cellulases accelerate the rate of rumen microbe’s colonization on feed particles and work in synergy with the endogenous rumen microbial enzymes18. Stable rumen environment is a key factor for achieving optimum milk production and a good animal’s health. In this concern, using cellulases in dairy animal’s feeding increase animal’s milk production by 5-25%19. Cellulases with their obvious importance as a feed additive are being imported for use in Egypt at a high cost. Therefore, the main objective of this work was to evaluate impact of cheap-locally produced cellulase compared with imported one on milk production and feed utilization by Baladi goats in early lactation.
MATERIALS AND METHODS
Goats and feed rations: Eighteen early lactating Baladi goats (29±0.5 kg) at 2nd to 3rd seasons of lactation were used in the present study. This research project was conducted from July, 2016 to June, 2019. The farm trials were extended for 9 weeks from March 5th to May 8th at Alstar farm for animal production, Khatatba city, Menoufia governorate, Egypt. The complete random design was used for dividing goats to 3 groups fed dry matter (DM) according to 4% of their body weight. The goats of first group was fed the control ration (50% concentrate feed mixture (CFM), 25% berseem (clover) hay and 25% wheat straw). According to results of previously conducted in vitro trials16, the second group was fed control ration+produced cellulase (Penicillium chrysogenum cellulase product contains 364772 units kg–1) at 42.16 (IU kg–1 of feed ration) (R1), while the third group was fed control ration+42.16 IU kg–1 DM of Pan-Zyme (commercial cellulase product contains 600000 units kg–1 and produced by VTR Bio-Tech Co, Ltd., China) (R2). The experimental ration was offered twice daily at 8.00 am and 4:00 pm. The enzymes were introduced once a day to each goat of second and third group at the morning feeding. Fresh water was available to the animals all the time. The feed ingredients and the chemical composition of the experimental control ration are shown in Table 1.
| Table 1: | Chemical composition of control ration’s ingredients (on DM basis) |
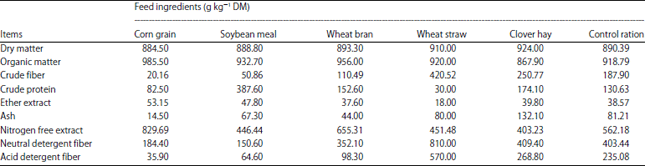 | |
TMR: Total mixed ration, control ration concentrate: roughage ratio was 1:1 on DM basis, the concentrate feed mixture (CFM) consisted of 60% corn, 22.6% soybean meal, 15% wheat bran,1% limestone, 0.4% minerals and 1% NaCl, the roughage part of the ration consisted of 50% Berseem hay and 50% wheat straw | |
Apparent digestibility: Nutrients digestion coefficients were determined by using silica as an internal marker. During the last 3 days of each month of the experimental period, fecal grab samples were collected in cloth bag connected to the animal back at 12 pm, from 3 animals of each group. The collected feces were dried in an oven at 60°C for 48 h. The dried feces from each animal were mixed and ground to pass a 1 mm sieve in a feed mill for chemical analysis. The digestibility coefficient of nutrient was calculated according to the following equation20:
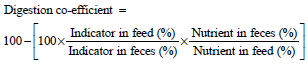
Feedstuffs and fecal samples were analyzed according to the AOAC21 methods to determine dry matter (DM), crude protein (CP), ether extract (EE) and ash contents. Organic matter (OM) contents were calculated by difference. The neutral detergent fiber (NDF) and acid detergent fiber (ADF) contents were determined using the methods described by Van Soest et al.22.
Sampling and analysis of blood serum: Blood samples were taken from jugular vein of 3 animals each group through the last 3 days of each month of the experimental period. At about 4 h after morning feeding the blood samples were collected in glass tubes and left to coagulate at room temperature. Serum was separated by centrifugation at 4000 Xg/20 min. and kept frozen at -20°C for later analysis. Serum urea, aspartate aminotransferase (AST), alanine aminotransferase (ALT), glucose, creatinine and cholesterol concentration were determined using specific kits (Stanbio Laboratory, Boerne, TX, USA) following manufacturer instructions.
Sampling and analysis of milk: Goats were milked by hand twice a day at 8:00 am and 8:00 pm by milking one teat while, the other one was left to the kid for suckling. Milk samples were taken after the end of adaptation period during the last 3 days of each week up to the end of experimental period. Samples of milk were collected immediately from each animal after morning and evening milking and milk yield was recorded. The sample of each animal represented a mixed sample of constant percentage of the evening and morning yield. Milk samples were analyzed for total solids, fat, true protein and lactose by infrared spectrophotometry (Milkotester LM2, Belovo, Bulgaria). Fat corrected milk (4% fat) was calculated by using the following equation11:
FCM = 0.4 M+15 F
where, M is the milk yield (g) and F is the fat yield (g).
Fatty acids profile of milk fat was determined as methylated fat according to Park et al.23 and separated by gas liquid chromatography. Amino acid profile of milk protein was quantitatively determined according to AOAC21 using high performance amino acid analyzer (Biochrom 30) with EZChrom Software for data collection and processing.
Statistical analysis: Data obtained from this study were statistically analyzed by IBM SPSS Statistics for Windows24 using the following general model procedure:
Yij = μ+Ti+eij
where, Yij is the parameter under analysis of the ij (goats) in digestibility and lactation trails, μ is the overall mean, Ti is the effect due to treatment on the parameter under analysis, eij is the experimental error for ij on the observation. Duncan's multiple range tests was used to test the significance among means at probability level of 0.05.
RESULTS
Effect of cellulases supplementation on nutrient’s digestibility and ration’s nutritive values: The results of Table 2 showed that goats fed enzymes supplemented rations (R1 and R2) showed significant increase (p<0.05) for most of nutrients digestibility coefficients and total digestible nutrients (TDN) than those fed the control ration. But, no significant differences were found between all goats’ groups in CP, EE digestibility and digestible crude protein (DCP) values.
| Table 2: | Effect of cellulases supplementation on nutrient digestibility and nutritive values of the experimental rations |
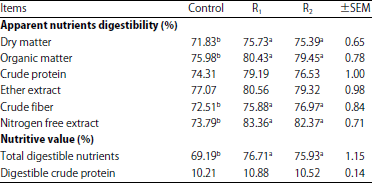 | |
Means with different letter (a, b) in the same row are significantly different at p<0.05 | |
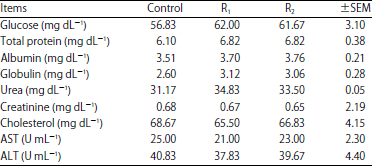
| Table 3: | Effect of cellulases supplementation on goat’s blood parameters |
 | |
| Table 4: | Effect of cellulases supplementation on goat’s milk yield and milk composition |
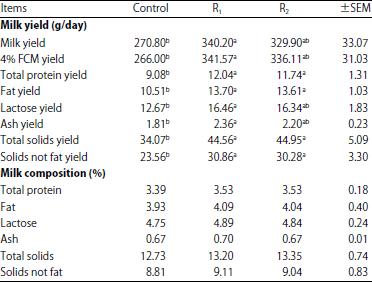 | |
Means with different letter (a, b) in the same row are significantly different at p<0.05 | |
Effect of cellulases supplementation on goat’s blood parameters: Cellulases supplemented goats (R1 and R2) had numerical but not significant higher serum glucose, protein, albumin and globulin concentration than those of the control (Table 3). No significant changes were detected between all goats’ groups in creatinine, urea, cholesterol, aspartate aminotransferase (AST) and alanine aminotransferase (ALT) values.
Effect of cellulases supplementation on goat’s milk yield and milk composition: Milk composition of supplemented goats with cellulases was not affected significantly compared to goats of the control (Table 4). But goats fed the produced cellulase supplemented ration (R1) had higher (p<0.05) milk, 4% fat corrected milk (FCM) and all milk components yields than those fed the control. The goats fed ration supplemented with the Pan-Zyme (commercial feed enzyme) (R2) showed numerical (but not significant) increase in milk, 4% fat corrected milk (FCM) and lactose yields compared with those of the control.
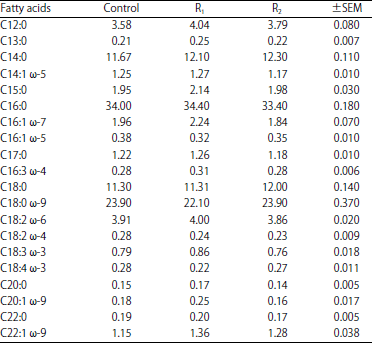
| Table 5: | Effect of cellulases supplementation on goat’s milk fatty acids profile |
 | |
| Table 6: | Effect of cellulases supplementation on goat’s milk amino acids profile |
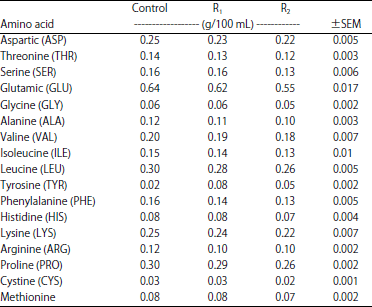 | |
Also, there were no significant differences between goats fed enzymes supplemented rations (R1 and R2) in milk and its component's yields. Although, there was enhancement in milk fat yield of the enzymes treated goats, cellulases supplementation has no effect on milk fatty acids profile (Table 5). Also, data of Table 6 showed that cellulases supplementation has no effect on milk amino acids profile.
DISCUSSION
Higher DM, OM, CF, NFE and TDN digestibility for goats fed enzymes treated rations (R1 and R2) than control (Table 2) is may be due to break down of anti-nutritional factors (cell wall fibers) of the supplemented rations and liberation of more soluble carbohydrate for the action of rumen microflora. Absence of the enzymes effect on the CP digestibility is may be due to low degradability of corn protein (zein) in the rumen. Similar positive results were reported by Azzaz et al.12,17 who found that addition of cellulolytic enzymes to goats and sheep’s diets improve their ability for digestion of banana wastes and date seeds respectively. On the contrary, Wahyuni et al.25 revealed that the enzyme addition to TMR containing oil palm frond silage did not affect the nutrients digestibilities by male goat.
Absence of the change in the urea, ALT and AST concentrations for cellulases treated goats indicated that cellulases addition has no harmful effect on kidney and liver functions. Higher DM, OM, CF and NFE digestibility (Table 2) may be the reason for the numerical higher blood glucose and protein concentrations. These findings are supported by many of studies12,17,26 on sheep and goats, which reflecting the safety of cellulases utilization in animal feeding.
The marked increase in milk and its component’s yields for goats fed cellulases supplemented rations (R1 and R2) is probably due to feed utilization improvement as a result of higher efficacy of nutrients digestion, higher production of ruminal propionate and microbial protein synthesis by enzyme's treated goats. These results are in good agreement with those obtained by Azzaz et al.12,17,26, who reported that lactating goats treated with fibrolytic enzymes showed higher milk and fat corrected milk production with no effect on milk composition. On the other hand, Titi and Lubbadeh27 recorded significant increased in milk total solids, protein and fat percentages due to fibrolytic enzymes supplementation. On the contrary, Flores et al.28 noticed no effect of fibrolytic enzyme addition on lactational performance of the treated ewes. Although, there was enhancement in milk fat yield of the enzymes treated goats, cellulases supplementation has no effect on milk fatty acids profile. In contrast, Abd El-Aziz et al.29 reported that feeding lactating buffaloes on fibrolytic enzymes supplemented diets caused an increase in milk long chain fatty acids, mono and poly-unsaturated fatty acids with a decrease in conjugated linoleic acid (CLA) content. As in the case of milk fatty acids profile, cellulases supplementation has no effect on milk amino acids profile. This may be due to lack cellulases supplementation for making positive change in goat's milk protein and fat content (percentages).
Finally, it was obvious from this study that locally produced cellulase show superiority over the commercial cellulase source for improving goat's diets digestion and milk productivity. It is known that higher cost of production is perhaps the major constraint in commercialization of new sources of enzymes. In current study, cellulase was produced on cheap carbon sources (mainly agricultural by products), which make their production process more economic. On the other hand, the stage of lactation may be has potent impact on goats performance, therefore, it is recommended to conduct this study on goats in late lactation for data confirmation.
CONCLUSION
It could be concluded that inclusion of cellulases in lactating goat’s rations have no deleterious effects on the treated goat’s health, but improved their feed utilization and milk production with no positive effect on milk fatty and amino acids profiles.
SIGNIFICANCE STATEMENT
This study discover the potential use of new product of cellulase in feeding of lactating goats in early lactation. This study can be beneficial for breeders of small ruminants, as cellulase (feed additives) can improve the productive performance of farm animals in economic way. Also this study will help the researcher to uncover the critical areas of use feed enzymes effectively to maximize dairy animal's productivity and increase the quality of the resultant milk.
REFERENCES
- Azzaz, H.H., H.A. Murad and T.A. Morsy, 2015. Utility of Ionophores for ruminant animals: A review. Asian J. Anim. Sci., 9: 254-265.
CrossRefDirect Link - Ismail, S.A., A.M. Abdel-Fattah, M.A. Emran, H.H. Azzaz, M.S. El-Gamal and A.M. Hashem, 2018. Effect of partial substitution of ration's soybean meal by biologically treated feathers on rumen fermentation characteristics (in vitro). Pak. J. Biol. Sci., 21: 110-118.
CrossRefDirect Link - Azzaz, H.H., H.M. Ebeid, T.A. Morsy and S.M. Kholif, 2015. Impact of feeding yeast culture or yeast culture and propionibacteria 169 on the productive performance of lactating buffaloes. Int. J. Dairy Sci., 10: 107-116.
CrossRefDirect Link - Azzaz, H.H., T.A. Morsy and H.A. Murad, 2016. Microbial feed supplements for ruminant's performance enhancement. Asian J. Agric. Res., 10: 1-14.
CrossRefDirect Link - Azzaz, H.H., H.A. Aziz, H. Alzahar and H.A. Murad, 2018. Yeast and Trichoderma viride don't synergistically work to improve olive trees by products digestibility and lactating Barki ewe's productivity. J. Biol. Sci., 18: 270-279.
CrossRefDirect Link - Azzaz, H.H., E.S.A. Farahat, T.A. Morsy, H.A. Aziz, F.I. Hadhoud and M.S. Abd-Alla, 2016. Moringa oleifera and Echinacea purpurea as supplements for rhamani lactating Ewe's diets and their effect on rumen characteristics, nutrients digestibility, blood parameters, milk production, composition and its fatty acid profile. Asian J. Anim. Vet. Adv., 11: 684-692.
CrossRefDirect Link - Kholif, A.E., A.Y. Kassab, H.H. Azzaz, O.H. Matloup, H.A. Hamdon, O.A. Olafadehan and T.A. Morsy, 2018. Essential oils blend with a newly developed enzyme cocktail works synergistically to enhance feed utilization and milk production of Farafra ewes in the subtropics. Small Rumin. Res., 161: 43-50.
CrossRefDirect Link - Murad, H.A. and H.H. Azzaz, 2010. Cellulase and dairy animal feeding. Biotechnology, 9: 238-256.
CrossRefDirect Link - Murad, H.A. and H.H. Azzaz, 2011. Microbial pectinases and ruminant nutrition. Res. J. Microbiol., 6: 246-269.
CrossRefDirect Link - Azzaz, H.H., H.A. Murad, A.M. Kholif, T.A. Morsy, A.M. Mansour and H.M. El-Sayed, 2013. Increasing nutrients bioavailability by using fibrolytic enzymes in dairy buffaloes feeding. J. Biol. Sci., 13: 234-241.
CrossRefDirect Link - Azzaz, H.H., A.A. Aboamer, H. Alzahar, M.M. Abdo and H.A. Murad, 2019. Effect of xylanase and phytase supplementation on goat's performance in early lactation. Pak. J. Biol. Sci., 22: 265-272.
CrossRefDirect Link - Azzaz, H.H., A.M. Kholif, H.A. Murad, M.A. Hanfy and M.H. Abdel Gawad, 2012. Utilization of cellulolytic enzymes to improve the nutritive value of banana wastes and performance of lactating goats. Asian J. Anim. Vet. Adv., 7: 664-673.
CrossRefDirect Link - Abd El Tawab, A.M., H.A. Murad, M.S.A. Khattab and H.H. Azzaz, 2019. Optimizing production of tannase and in vitro evaluation on ruminal fermentation, degradability and gas production. Int. J. Dairy Sci., 14: 53-60.
CrossRefDirect Link - Azzaz, H.H., H.A. Murad, A.M. Kholif, M.A. Hanfy and M.H. Abdel Gawad, 2012. Optimization of culture conditions affecting fungal cellulase production. Res. J. Microbiol., 7: 23-31.
CrossRefDirect Link - Aboul-Fotouh, G.E., G.M. El-Garhy, H.H. Azzaz, A.M. Abd El-Mola and G.A. Mousa, 2016. Fungal cellulase production optimization and its utilization in goat’s rations degradation. Asian J. Anim. Vet. Adv., 11: 824-831.
CrossRefDirect Link - Khattab, M.S.A., H.H. Azzaz, A.M. Abd El Tawab and H.A. Murad, 2019. Production optimization of fungal cellulase and its impact on ruminal degradability and fermentation of diet. Int. J. Dairy Sci., 14: 61-68.
CrossRefDirect Link - Azzaz, H.H., E.S.A. Farahat and H.M. Ebeid, 2017. Effect of partial replacement of corn grains by date seeds on rahmani ram’s nutrients digestibility and Nubian goat’s milk production. Int. J. Dairy Sci., 12: 266-274.
CrossRefDirect Link - Khattab, M.S.A., E.A. El-Bltagy, A.M. Abd El Tawab, O.H. Matloup, T.A. Morsy, H.H. Azzaz and M.M. Abdou, 2019. Productive performance of lactating buffaloes fed ration containing date seed and fibrolytic enzymes. J. Applied Sci., 19: 241-246.
CrossRefDirect Link - Gado, H.M., A.Z.M. Salem, P.H. Robinson and M. Hassan, 2009. Influence of exogenous enzymes on nutrient digestibility, extent of ruminal fermentation as well as milk production and composition in dairy cows. Anim. Feed Sci. Technol., 154: 36-46.
CrossRefDirect Link - Ferret, A., J. Plaixats, G. Caja, J. Gasa and P. Prio, 1999. Using markers to estimate apparent dry matter digestibility, faecal output and dry matter intake in dairy ewes fed Italian ryegrass hay or alfalfa hay. Small Rumin. Res., 33: 145-152.
CrossRefDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Park, S.J., C.W. Park, S.J. Kim, J.K. Kim and Y.R. Kim et al., 2002. Methylation methods for the quantitative analysis of Conjugated Linoleic Acid (CLA) isomers in various lipid samples. J. Agric. Food Chem., 50: 989-996.
CrossRefDirect Link - Wahyuni, R.D., W. Ngampongsai, C. Wattanachant, W. Visessanguan and S. Boonpayung, 2012. Effects of enzyme levels in total mixed ration containing oil palm frond silage on intake, rumen fermentation and growth performance of male goat. Songklanakarin J. Sci. Technol., 34: 353-360.
Direct Link - Azzaz, H.H., H.A. Aziz, E.S.A. Farahat and H.A. Murad, 2015. Impact of microbial feed supplements on the productive performance of lactating nubian goats. Global Vet., 14: 567-575.
CrossRefDirect Link - Titi, H.H. and W.F. Lubbadeh, 2004. Effect of feeding cellulase enzyme on productive responses of pregnant and lactating ewes and goats. Small Rumin. Res., 52: 137-143.
CrossRefDirect Link - Flores, C., G. Caja, R. Casals, E. Albanell and X. Such, 2008. Performance of dairy ewes fed diets with a fibrolytic enzyme product included in the concentrate during the suckling period. Animal, 2: 962-968.
CrossRefDirect Link - Abd El-Aziz, M., S.M. Kholif and T.A. Morsy, 2012. Buffalo's milk composition and its fat properties as affected by feeding diet supplemented with flaxseed or fibrolytic enzymes in early lactation. J. Life Sci., 4: 19-25.
Direct Link