
Zeinab I. Sadek
Department of Dairy Science, National Research Center, P.O. Box 12622, Dokki, Cairo, Egypt
B.M. Refaat
Department of Dairy Science, National Research Center, P.O. Box 12622, Dokki, Cairo, Egypt
E.H. Abd El-Shakour
Department of Botany and Microbiology, Faculty of Science, Al-Azhar University, Cairo, Egypt
Nayra Sh. Mehanna
Department of Dairy Science, National Research Center, P.O. Box 12622, Dokki, Cairo, Egypt
LiveDNA: 20.4398
M.S. Hassan
Department of Dairy Science, National Research Center, P.O. Box 12622, Dokki, Cairo, Egypt
International Journal of Dairy Science
Year: 2017 | Volume: 12 | Issue: 2 | Page No.: 93-104







ABSTRACT
Background: Processed cheese could be estimated as microbiologically safe, nevertheless microorganisms may gain entrance during all stages of processing especially spore forming bacteria, one way to limit the growth of undesirable microorganisms is incorporation of probiotics which show high antimicrobial activity against these pathogens. Materials and Methods: The antibacterial activity of six different lactic acid bacteria against Bacillus cereus and Clostridium perfringes were studied. Results: Lactobacillus curvatus and Lactobacillus acidophilus were the most effective cultures against B. cereus , while Leuconostoc mesenteroides against C. perfringes . Supplementation of processed cheese with L. curvatus and the mixed culture of the three other probiotics effectively suppressed the growth and survival of B. cereus and it could not be recovered after 14 days of storage and room temperature. While, supplementation of the cheese by metabolites of L. curvatus, L. acidophilus and mixed culture of L. curvatus, L. acidophilus and L. mesenteroides reduce the viable counts of B. cereus and the reduction reached 100% at 14 days using metabolites of L. curvatus and the mixed culture of L. curvatus, L. acidophilus and L. mesenteroides and at 21 days using L. acidophilus when the cheese stored at 25°C. But, cold storage delayed the reduction of B. cereus . Also, a pronounced reduction in viability of C. perfringes was observed with L. mesenterials culture, as C. perfinges could not be recovered after 30 and 21 days at room and refrigerator temperature, respectively. Conclusion: Supplementation of probiotic metabolites of L. mesenteroides and the mixed culture (L. curvatus , L. acidophilus and L. mesenteroides ) led to decrease the viability and survival of C. perfringens as it could not be recovered after 30 days at refrigerator temperature for 45 and 30 days respectively, at room temperature.
PDF Abstract XML References Citation
Received: October 06, 2016;
Accepted: November 02, 2016;
Published: February 25, 2019
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Zeinab I. Sadek, B.M. Refaat, E.H. Abd El-Shakour, Nayra Sh. Mehanna and M.S. Hassan, 2017. Biocontrol of Processed Cheese by Incorporation of Probiotic
Bacteria and its Metabolites. International Journal of Dairy Science, 12: 93-104.
DOI: 10.3923/ijds.2017.93.104
URL: https://scialert.net/abstract/?doi=ijds.2017.93.104
DOI: 10.3923/ijds.2017.93.104
URL: https://scialert.net/abstract/?doi=ijds.2017.93.104
INTRODUCTION
Processed cheese is made by heating a mixture of natural cheeses of different types and different ages with emulsifying salts and various ingredients under lower pressure and constant stirring until a homogenous mass of desired properties is formed1-3.
The growing demand for high-value health promoting foods such as dairy products has encouraged the industries and the research field, not only to explore new emerging food processes and formulations in which they are able to assure functional but also safe products4-6. Probiotics are recently defined by Krishna7 as a live microbial food ingredient. When probiotics are ingested in sufficient quantities, extra health benefits to the consumer will be added such as enhanced immune response, alleviation of symptoms of lactose intolerance, treatment of diarrhea, reduction of serum cholesterol, vitamin synthesis and anti-carcinogenic and anti-microbial activities8-12. There are different types of Lactic Acid Bacteria (LAB) they are used as probiotic which include Lactobacillus and Bifidobacterium13. Highly undesirable microbial contaminates of processed cheese are rod-shaped endospore-forming bacteria of the genera Bacillus and Clostridium, formation of the spores allows it to be resistance of heat, chemicals and other adverse environments that undergoes during processing preparation of the product. Bacillus cereus might be the most common cause of food borne illnesses worldwide, but barely reported to health agencies because its symptoms tend to be short-lived and self-limited14.
Moreover, Clostridium perfringens causes a toxin mediated disease represents as the 3rd most commonly reported food-borne illness15. Some C. perfringens strains produce important toxin named C. perfringens enterotoxin (CPE), which is responsible for several human gastrointestinal diseases16. In addition, B. ceraus reported to be capable of producing 2 types of enterotoxin, first diarrheal disease which occurs when ingested food contains 105 cells, second enteric disease which occurs when food contains preformed toxin17.
Lactic Acid Bacteria (LAB) are a group of Gram-positive bacteria including the genera Lactobacillus, Lactococcus, Leuconostoc and Streptococcus. It produces a wide range of antimicrobial metabolite comprising organic acids, carbon dioxide, ethanol, hydrogen peroxide, diacetyl and bacteriocins18. Bacteriocins of lactic acid bacteria have attracted the attention of many investigators especially in the recent years because of their use as a natural food preservative19. So, this study aimed to validate the effectiveness of antibacterial activity of some LAB against these pathogens for supplementation of processed cheese with suitable strains to suppress the growth of B. cereus and C. perfringens and subsequently to increase the safety and quality of the cheese.
MATERIALS AND METHODS
Bacterial strains
Lactic acid bacterial strains: Leuconostoc mesenteroides B-118 and Lactobacillus acidophilus were provided by Chr. Hansen’s Laboratory, Denmark. Streptococcus thermophilus and Lactococcus lactis were obtained from the Dairy Department, National Research Center (NRC). Lactobacillus curvatus B-4562 and Lactobacillus rhamnosus B-445 were supplied by the Northern Regional Research Laboratory, IIIinois, USA (NRRL). Leuconostoc mesenteroides B-118, Lactobacilluc acidophilus, Lactobacillus curvatus B-4562 and Lactobacillus rhamnosus B-445 were previously studied in (NRC) and proved to be probiotics20,21 by testing for probiotic characteristics (acid tolerance, survivability in different bile salt concentration and antibiotic susceptibility as well as antimicrobial activity to some pathogens according to FAO. and WHO.22.
Pathogenic strains: Three strains of B. cereus (tested as mixed strain) and also, three strains of C. perfringens which were isolated and identified previously23 were used for inoculation of the resultant cheese.
Media: The used media are tryptone soya broth (Oxoid) for antagonism, MRS broth media for culture of lactic acid. Bacteria strains, polymyxin-pyruvate-eggyolk-mannitol-bromothymol blue agar (PEMBA) (Oxoid) for B. cereus, perfringens agar (TSC) (Oxoid) for C. perfringens, M17 was used for Streptococcus thermophilus and Lactococcus lactis, (MRS agar medium) (Oxoid, UK) for Lactobacillus rhamnosus, Leuconostoc Medium (MRS with sucrose and sodium azide)24 for Leuconostoc mesenteroides, MRS medium with replacing glucose with trehalose and addition of bile salt for Lactobacillus acidophilus and MRS media with addition of sodium chloride for Lactobacillus curvatus.
Media used throughout the current investigation were in a dehydrated form and prepared according to the manufacturer's instructions.
Experimental procedures
Growth and preservation LAB strains: Each lactic acid bacteria culture was maintained individually in 11% (w/v) sterile Reconstituted Skim Milk (RSM) powder at 37°C for 24 h, except L. lactis, L. mesenteroides incubated at 30°C for 18 h and stored at 4°C between transfers, each culture was sub cultured twice in MRS medium (Oxoid) before use.
Screening for antibacterial activity of the tested LAB metabolites towards B. cereus and C. perfringens
Cell-free supernatant: The cell-free supernatant fluids were obtained by centrifuging the overnight cultures of the tested LAB strains (grown at 37°C for 24 h in MRS broth media) at 4000 rpm for 15 min at 4°C. Supernatant was filter-sterilized through 0.22 μm Millex-GV membranes (Millipore) and pH was adjusted to 6 to exclude the organic acid effects.
Antibacterial activity: The activity of the resulting metabolites of the tested LAB strains were tested against B. cereus and C. perfringens using the well diffusion agar assay as described by Lyon and Glatz25. Aerobically for B. cereus and anaerobically for C. perfringens in gas pack anaerobic jar using kits of anaerobic Gas Generating Kits (Oxoid). Each strain was tested in triplicate per plate and plates were incubated for 24 h at 37°C. The ability of each probiotic strain metabolite to inhibit the growth of B. cereus and C. perfringens was determined by measuring the diameter in millimeter of the clear inhibition zone formed around discs.
Antagonism activity of the tested LAB strains against pathogenic strains: Bacillus cereus and C. perfringens strains were transferred into tryptone soya broth (Oxoid) and each tested LAB strains were cultured in MRS broth media and incubated at 37°C for 24 h. The cultures individually were diluted in saline solutions to an appropriate inoculum size (105 CFU mL–1 for B. cereus and C. perfringens and 107 CFU mL–1 for lacic acid bacteria tested strains) and were ready for inoculation together into 100 mL tryptone soya broth. Each of six LAB strains in tryptone soya broth was inoculated with Bacillus cereus and C. perfringens with two controls for pathogenic. All incubated at 37°C with gas pack anaerobic jar using anaerobic kits.
The counts of B. cereus and C. perfringens in the mixed culture were measured after incubation at intervals (0, 24 and 48 h).
| Table 1: | Ingredients used in the preparation of blends for processed cheese |
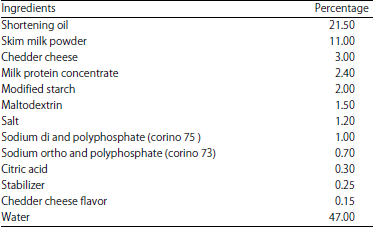 | |
Manufacturing of processed cheese (triangles): Processed cheese was manufactured as described by Mayer26 and Schar and Bosset27. The composition and percentage of ingredients used are given in Table 1. The mixture of ingredients was heated in a batch cooker with constant agitation, until a homogeneous mass is obtained. The melting temperature of processed cheese was 85±1°C and the total melting time was 15 min from the beginning of heating to the beginning of discharge. The resultant was manually filled into sterilized glass jar and covered with aluminum foil and their covers.
All the used ingredients were tested and were found free from aerobic and anaerobic spore forming bacteria. The batches were performed in the laboratories of Faculty of Agriculture, Ain Shams University.
The first batch was divided into five equal portions and were inoculated with LAB strains, B. cereus and C. perfringens strains at 45°C and divided according to antimicrobial results as following:
| • | Inoculation with 2% L. curvatus |
| • | Inoculation with 2% L. acidophilus |
| • | Inoculation with 2% mixed culture (L. curvatus, L. acidophilus and L. mesenteroides) |
| • | Inoculation with 2% L. mesenteroides |
| • | As control without inoculated with LAB strains |
All these treatments were obtained with addition of citric acid to adjust pH. The first and second treatments were inoculated separately with 105 CFU mL–1 of B. cereus strain.
Whereas, third treatment divided into two equal portions and inoculated one portion with 105 CFU mL–1 of B. cereus strain and second portion with 105 CFU mL–1 of C. perfringens strain. The fourth treatment was inoculated with 105 CFU mL–1 of C. perfringens strain.
Also, the fifth treatment divided into two equal portions and inoculated one portion with 105 CFU mL–1 of B. cereus strain and second portion with 105 CFU mL–1 of C. perfringens strain as control without inoculated with LAB strains.
The second batch was divided into five equal portions as mentioned previously for the first batch, but used the metabolites of LAB with percent 2% which prepared as mentioned before but the strains grown in sterilized reconstituted skim milk were incubated at 37°C for 24 h and then manufacture processed cheese from these cultures without addition of citric acid. The control made from reconstituted skim milk powder without inoculated with metabolites of LAB.
All the cheese treatments were filled into sterilized glass jars and covered with aluminum foil and their covers. Half of these containers from each treatment were stored at 25°C and the other half at 7±2°C.
All samples including the main control, the contaminated controls and the treated contaminated cheeses were subjected for bacteriological analysis to investigate the behavior of the tested pathogens at zero time and after 1, 2, 3, 7, 14, 21, 30, 45 and 60 days of storage at refrigerator and room temperature. Each pathogenic was recovered on its selective agar media.
Bacteriological analysis: To all cheese samples (25 g) were homogenized for 1 min in 225 mL of sterile solution (2% w/v) of sodium citrate. Analysis was carried out using the following procedures.
Bacillus cereus counts: Bacillus cereus counts were carried out by spreading 0.1 mL of the appropriate dilution onto PEMBA medium (Oxoid). The incubation temperature was 37°C for 18-24 h.
Clostridium perfringens counts: Clostridium perfringens counts were carried out on Perfringens agar (TSC) (Oxoid) according to Varadaraj28.
RESULTS AND DISCUSSION
Antagonistic activity of some lactic acid bacteria against aerobic spore former (B. cereus) and anaerobic spore former (C. perfringens): Bacteriocin and bacteriocin like producing species have now been identified among all the genera that comprise the Lactic Acid Bacteria (LAB) including Lactococcus, Streptococcus, Lactobacillus, Leuconostoc and Pedicoccus29.
These bacteriocins are extremely important for preventing the growth of spoilage and pathogenic bacteria.
Antibacterial activity of some lactic acid bacteria against B. cereus and C. perfringens: Using the well diffusion agar assay, the antibacterial activity of six different lactic acid bacteria against B. cereus and C. perfringens strains is presented in Fig. 1 and 2. The recorded results revealed that all the tested LAB have variable antibacterial activity since supernatants of the six tested LAB strains inhibited the growth of B. cereus and this resulted different inhibition zone diameter (9-13 mm).
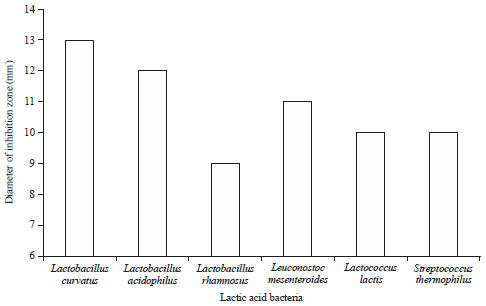 | |
| Fig. 1: | Antibacterial activity of some lactic acid bacteria against Bacillus cereus |
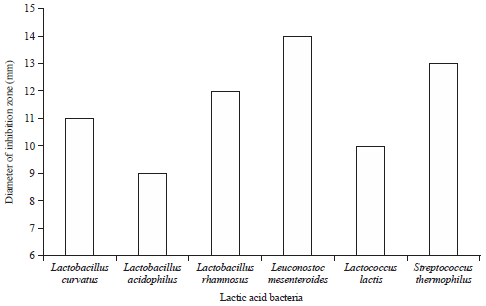 | |
| Fig. 2: | Antibacterial activity of some lactic acid bacteria against Clostridium perfringens |
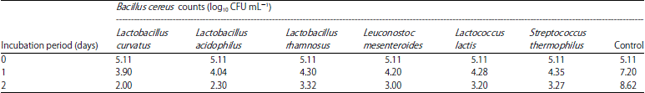
| Table 2: | Antagonism activity of some lactic acid bacteria against B. cereus in mixed cultures |
 | |
| Table 3: | Antagonism activity of some lactic acid bacteria against C. perfringens in mixed culture |
 | |
Lactobacillus curvatus was the most effective culture among all the tested cultures and formed the highest inhibition zone (13 mm diameter). While, Lactobacillus rhamnosus exhibited the smallest inhibition zone (9 mm). Regarding the antibacterial activity of the supernatants of LAB tested strain against C. perfringens, the largest inhibition activity were deserved by supernatants of Leuconostoc mesenteroides followed by Streptococcus thermophilus and Lactobacillus rhamnosus which inhibitd the growth of C. perfringens and this appeared by inhibition zones; 14, 13 and 12 mm, respectively (Fig. 2). Additionally, L. curvatus proved to have suppressive effect toward C. perfringens with inhibition zone diameter 11 mm, while, L. acidophilus show the lowest suppressive effect.
Ahmadova et al.30 reported that the cell free supernatant of culture of L. curvatus A6 which exhibited the presence of curvacin A encoding gene inhibited the growth of Listeria monocytogenes and B. cereus strains.
Saroj31 reported that the cell-free supernatant bacteriocins produced by strain of L. acidophilus showed inhibition zone against Bacillus spp.
Morever, the ability of LAB to produce bacteriocins, some of which have a broad spectrum of activity were reported by Gillor et al.32.
Recently, the scientist proved that, leucocin produced by Leuconostoc showed antibacterial activity againist Gram-positive bacteria33.
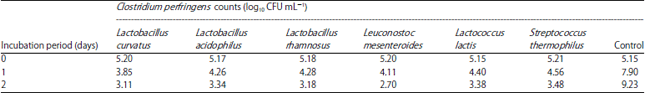
Antagonistic activity of some lactic acid bacteria against B. cereus and C. perfringens: The antibacterial activity of six LAB strains toward B. cereus and C. perfringens in tryptone soya broth was evaluated (Table 2, 3).
It is obvious that, B. cereus and C. perfringens proliferated well in tryptone soya broth and increasing in counts by 3.51 and 4.08 log cycle respectively, in control after 48 h. While, in mixed culture with the tested LAB bacteria, a pronounced reduction in viability of B. cereus and C. perfringens was observed. Bacillus cereus showed rapid decline in mixed culture with L. curvatus, as its numbers were decrease by 3.11 log cycle after 48 h of incubation. Lactobacillus acidophilus and L. mesenteroides also showed strong antibacterial activity against the pathogen and reduce survival of B. cereus by 2.81, 2.11 log cycle, respectively after 48 h of incubation. While, L. rhamnosus, L. lactis and S. thermophilus show slightly weaker inhibitory activity as numbers of B. cereus decreased by 1.79, 1.91 and 1.84 log cycles, respectively after 48 h of incubation (Table 1). These results indicated that tested strains of LAB could restrict the activity of B. cereus (growth or survival).
Among the tested probiotics L. mesenteroides presented a high inhibitory effect against C. perfringens resulting of decreasing the counts from 5.20-4.11 log10 CFU mL–1 after 24 h of inoculation with reduction percent (21.3%) reaching 2.70 log10 CFU mL–1 with reduction percent (48.7%) after 48 h of inoculation. Also, L. curvatus and L. rhamnosus were effectively inhibited the growth of C. perfringens leading to reduce counts by 2.09 and 2 log cycle, respectively after 48 h of incubation. Whereas, L. acidophilus, L. lactis and S. thermophilus were able to inhibit growth and decrease counts by (1.83, 1.77 and 1.73 log cycle) after 48 h.
The inhibition of B. cereus was due to the antimicrobial metabolites e.g., bacteriocin, organic acids and hydrogen peroxide by the tested strains of Lactobacillus and Lactococcus34.
Mkrtchyan et al.35 and Nazzaro et al.36 reported that L. acidophilus exhibited broad spectrum activity against L. monocytogenes and B. cereus.
Hereupon, L. mesenteroides gained the highest anticlostridial activity than the other tested strains and appeared to be promising culture.
According to the results of antibacterial and mixed culture assay, three strains of LAB which was previously proved to be probiotics by Mabrouk20 and Ibrahim21 (L. curvatus, L. acidophilus and L. mesenteroides) which showed the highest antimicrobial effect toward B. cereus and C. perfringens were selected for further study.
Biocontrol of processed cheese by incorporation of probiotic bacteria and its metabolites: In recent years, several reports were dealing with antibacterial activity of LAB especially probiotics and their role for liming the growth of undesirable microorganisms.
The processed cheese was manufactured using the chosen probiotic bacteria through the results of antibacterial and antagonistic activity of some LAB towards B. cereus and C. perfringens in the previous section (L. curvatus, L. acidophilus and L. mesenteroides) or their metabolites with inoculation of B. cereus and C. perfringens in the processed cheese.
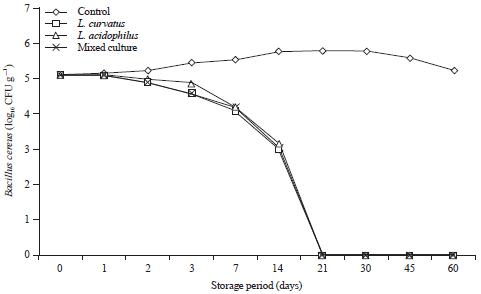
Behavior of B. cereus and C. perfringens and probiotics during storage of processed cheese at 25°C and at 7±2°C for 60 days: The pattern of selected probiotics and B. cereus growth in inoculated processed cheese during storage at 25 and 7°C for 60 days are shown in Fig. 3 and 4.
Generally, B. cereus showed a rapid decline in its culturable numbers in cheese manufactured by inoculation with the selected probiotics and B. cereus during storage at 25°C as well as 7°C.
It is obvious that B. cereus can survive in control cheese till the end of storage period (60 days) reaching maximum counts after 30 days of storage period (Fig. 3). Such counts were nearly the estimated count to be sufficient to produce toxin as detectable enterotoxin concentration considered with colonization37 level >106. However, in compromised consumers a much smaller dose of 1.2×103 mL–1 may cause illness38.
In contrast, supplementation of processed cheese with L. curvatus effectively suppressed the growth and survival of B. cereus as it showed higher inhibition effect than inoculation with L. acidophilus and the pathogen could not be recovered after 14 days of storage with decreasing of its counts by 1.04 log cycle and the roduction of count was 1.52 log cycle as compared to the control cheese after 7 days of storage at room temperature (Fig. 3), but at cold storage, the pathogen can survive for more than 14 days and no viable cells were detected at 21 days of storage (Fig. 4).
Also, supplementation of cheese with the probiotic L. acidophilus led to reduce the viability and survival of B. cereus as it could not be recovered after 21 days of storage at room temperature (25°C) as well as at cold storage 7±2°C (Fig. 3, 4).
It could not be recover after 21 days with supplementation of both probiotic strains (L. curvatus, L. acidophilus and L. mesenteroides) at refrigerator temperature (Fig. 4).
Concerning mixed culture, cheese (L. curavatus, L. acidophilus and L. mesenteroides) were found to suppress the survival of B. cereus and could not be recovered after 14 days at room temperature (Fig. 3) and 21 days at refrigerator temperature, respectively (Fig. 4).
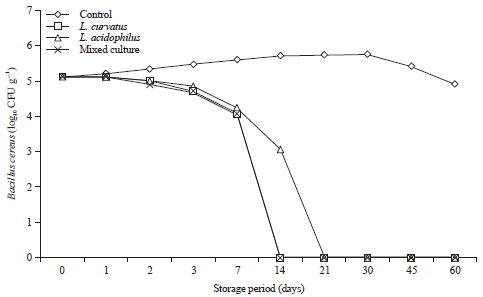 | |
| Fig. 3: | Behavior of Bacillus cereus during storage of processed cheese supplement with probiotic strains at 25°C for 60 days, Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
 | |
| Fig. 4: | Behavior of Bacillus cereus during storage of processed cheese supplemented with probiotics at 7 ± 2°C for 60 days. Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
The reduction of viable counts of B. cereus could be mainly attributed to the production of antimicrobial substance produced by the used probiotic cultures.
All probiotics were proliferated with gradual increase in counts during the first weak and then the increase was slight. These results are in line with those reported by Rukure and Bester39.
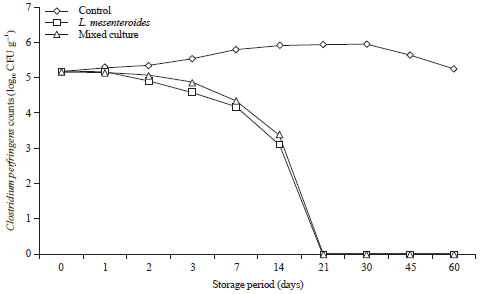
Figure 5 and 6 show that C. perfringens can be proliferated in control processed cheese without probiotics reached the maximum growth rate at 14 days of the storage period at room temperature. Also, despite storage under refrigerated conditions (7±2°C) there are opportunity for survival and proliferation of the pathogen, as reached the maximum rate in control cheese after 30 days. While, in processed cheese with the selected probiotic bacteria, a pronounced reduction in viability of C. perfringens were observed. When processed cheese supplemented with L. mesenteroides, C. perfringens could not be recovered after 30 and 21 days at room and refrigerator temperature, respectively (Fig. 5 and 6).
Additionally, C. perfringens counts were continually decrease and could not be recovered after 21 days in processed cheese made with mixed culture (L. curvatus, L. acidophilus and L. mesenteroides) which appears to have more inhibitory activity than L. mesenteroides alone at room and refrigerator temperature.
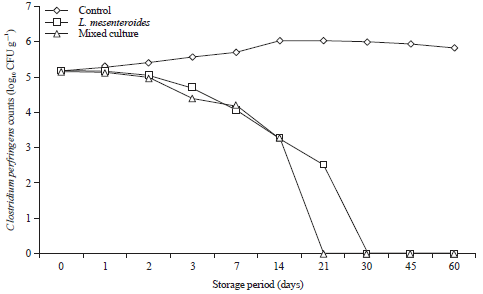 | |
| Fig. 5: | Behavior of Clostridium perfringens during storage of processed cheese supplemented with probiotics at 25°C for 60 days. Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
 | |
| Fig. 6: | Behavior of Clostridium perfringens during storage of processed cheese supplemented with probiotics at 7±2°C for 60 days. Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
In this respect De Vuyst and Vandamme40 reported that the bacteriocins e.g., nisin, pediocin and leuconocin have a relatively broad spectrum of activity against Gram-positive bacteria.
The antimicrobial effects of Leuconostoc against pathogenic and spoilage microorganisms have been reported early due to the production of organic acid, peroxide as well as bacteriocin41.
Behavior of B. cereus and C. perfringens during storage of processed cheese supplemented with probiotic metabolites at 25°C and at 7±2°C for 60 days: The LAB are considered generally recognized as safe (GRAS) by the US Food and Drug Administration42. Thus, the use of their metabolites as biological preservatives has been discussed43.
Growth and survival of B. cereus in processed cheese with and without the chosen probiotics metabolites during storage for 60 days at 25°C and at 7±2°C are shown in Fig. 7 and 8.
Results show that, B. cereus grew well in control cheese and reached the maximum growth rate at 30 days of the storage period either at room temperature or refrigerator temperature (Fig. 7 and 8).
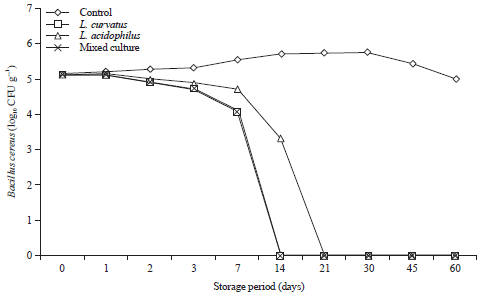 | |
| Fig. 7: | Behavior of Bacillus cereus during storage of processed cheese supplemented with probiotic metabolites at 25°C for 60 days. Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
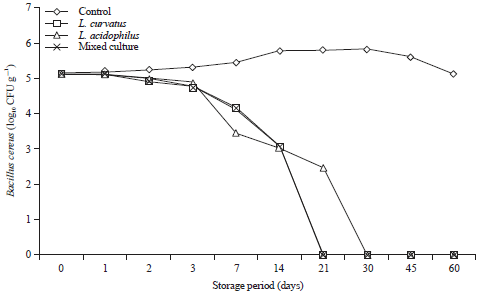 | |
| Fig. 8: | Behavior of Bacillus cereus during storage of processed cheese supplemented with probiotic metabolites at 7±2°C for 60 days. Mixed culture: L. curavatu, L. acidophilus and L. mesenteroides |
On the other hand, supplementation of the cheese by metabolites of L. curvatus, L. acidophilus and mixed culture of L. curvatus, L. acidophilus and L. mesenteroides reduce the viable counts of B. cereus and the reduction reached 100% after 14 days by using metabolites of L. curavatus and the mixed culture of (L. curvatus, L. acidophilus and L. mesenteroides) metabolites and after 21 days by using L. acidophilus when the cheese stored at 25°C (Fig. 7). But, cold storage (7±2°C) of the supplemented cheese delayed the reduction of B. cereus (Fig. 7).
These results clearly indicate that L. curvatus showed strong antibacterial activity against B. cereus, weather with supplementation as culture or its metabolites in the processed cheese. In this respect, Kumar and Arumugam44 reported inhibition zone (9 mm) with curvacin A (produced by L. curvatus) towards B. cereus.
Recently, Ahmadova30 found that bacteriocin produced by L. curvatus strain was heat stable, resistant to physiological concentrations of bile salts and active in a broad pH range and bacteriostatic against Bacillus cereus.
On the other hand, the results were pointed out that supplementation of probiotic metabolites of L. mesenteroides and the mixed culture (L. curvatus, L. acidophilus and L. mesenteroides) showed similar results leading to decrease the viability and survival of C. perfringens as it could not be recovered after 30 days. While, the counts of the pathogen in the control cheese reached the maximum (5.93 log10 CFU g–1) at the same time at refrigerator temperature (Fig. 9).
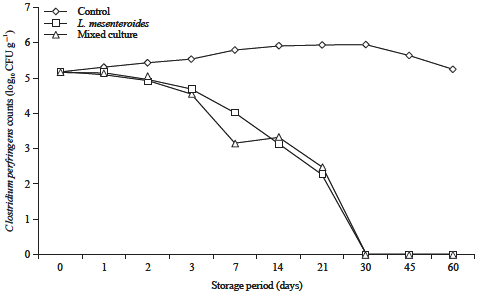 | |
| Fig. 9: | Behavior of Clostridium perfringens during storage of processed cheese supplemented with probiotic metabolites at 7±2°C for 60 days. Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
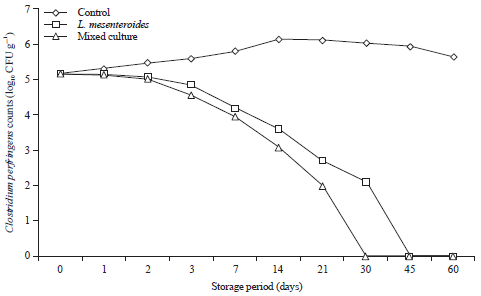 | |
| Fig. 10: | Behavior of Clostridium perfringens during storage of processed cheese supplemented with probiotic metabolites at 25°C for 60 days. Mixed culture: L. curvatus, L. acidophilus and L. mesenteroides |
On the contrary, survival of C. perfringens could not be recovered after 45 and 30 days for supplementation of L. mesenteroides and mixed culture (L. curvatus, L. acidophilus and L. mesenteroides) metabolites when the cheese stored for 60 days at room temperature (Fig. 10).
Ennahar et al.45 found that, the bactericidal activity of leuconocin seems to be targeting primarily Listeria strains. In addition Clostridium and Bacillus have been reported to be sensitive to this bacteriocin.
More recent, Devi and Halami46 recorded that some pathogenic bacteria being sensitive to the leucocin include Clostridium.
CONCLUSION
It is recommended to use improvement of product quality and safety could be applying of Good Hygienic practices (GHP) and Hazard Analysis and Critical Control Point (HACCP) system. The other strategy for reducing or controlling these pathogens can be achieved through incorporation of some probiotics or its metabolites, which suppress or arrest these microorganisms and so to improve the hygienic quality and safety of the product.
REFERENCES
- Bunka, F., O. Kriz, A. Velickova, L. Bunkova and S. Kracmar, 2009. Effect of acid hydrolysis time on amino acid determination in casein and processed cheeses with different fat content. J. Food Compos. Anal., 22: 224-232.
CrossRefDirect Link - Bubelova, Z., B. Tremlova, L. Bunkova, M. Pospiech, E. Vitova and F. Bunka, 2015. The effect of long-term storage on the quality of sterilized processed cheese. J. Food Sci. Technol., 52: 4985-4993.
CrossRefDirect Link - Rahaie, S., S.M.T. Gharibzahedi, S.H. Razavi and S.M. Jafari, 2014. Recent developments on new formulations based on nutrient-dense ingredients for the production of healthy-functional bread: A review. J. Food Sci. Technol., 51: 2896-2906.
CrossRefDirect Link - Moghari, A.A., S.H. Razavi, M.R. Ehsani, M. Mousavi and T.H. Nia, 2014. Chemical, proteolysis and sensory attributes and probiotic microorganisms viability of Iranian ultrafiltered-Feta cheese as a function of inulin concentration and storage temperature. Qual. Assurance Saf. Crops Foods, 7: 217-224.
CrossRefDirect Link - Ladjevardi, Z.S., S.M.T. Gharibzahedi and M. Mousavi, 2015. Development of a stable low-fat yogurt gel using functionality of psyllium (Plantago ovata Forsk) husk gum. Carbohydr. Polym., 125: 272-280.
CrossRefDirect Link - Krishna, K.V., S. Arunkumar, J.S. Arunkumar and S.V. Koti, 2014. Probiotics for health and disease-myth or truth???? Int. J. Biomed. Adv. Res., 5: 406-408.
CrossRefDirect Link - Shah, N.P., 2007. Functional cultures and health benefits. Int. Dairy J., 17: 1262-1277.
CrossRefDirect Link - Salari, M., S.H. Razavi and S.M.T. Gharibzahedi, 2014. Characterising the synbiotic beverages based on barley and malt flours fermented by Lactobacillus delbrueckii and paracasei strains. Qual. Assurance Saf. Crops Foods, 7: 355-361.
CrossRefDirect Link - Shokoohi, M., S.H. Razavi, M. Labbafi, A. Vahidinia and S.T. Gharibzahedi, 2015. Wheat sprout flour as an attractive substrate for the producing probiotic fermented beverages: Process development and product characterisation. Qual. Assur. Saf. Crops Foods, 7: 469-475.
CrossRefDirect Link - Torkamani, M.G., S.H. Razavi and S.M.T. Gharibzahedi, 2015. Critical quality attributes of Iranian Taftoon breads as affected by the addition of rice bran sourdough with different lactobacilli. Qual. Assur. Saf. Crops Foods, 7: 305-311.
CrossRefDirect Link - Ehsannia, S. and M.R. Sanjabi, 2016. Physicochemical, microbiological and spoilage analysis of probiotic processed cheese analogues with reduced emulsifying salts during refrigerated storage. J. Food Sci. Technol., 53: 996-1003.
CrossRefDirect Link - Macfarlane, S., E. Furrie, J.H. Cummings and G.T. Macfarlane, 2004. Chemotaxonomic analysis of bacterial populations colonizing the rectal mucosa in patients with ulcerative colitis. Clin. Infect. Dis., 38: 1690-1699.
CrossRefDirect Link - Abu Elnaga, A.S.M., R.H. Hedia, N.S. Ata and M.S. Zaki, 2014. Bacterial aspect of food poisoning. Life Sci. J., 11: 290-298.
Direct Link - Grass, J.E., L.H. Gould and B.E. Mahon, 2013. Epidemiology of foodborne disease outbreaks caused by Clostridium perfringens, United States, 1998-2010. Foodborne Pathog. Dis., 10: 131-136.
CrossRefDirect Link - Lahti, P., A. Heikinheimo, T. Johansson and H. Korkeala, 2008. Clostridium perfringens type A strains carrying a plasmid-borne enterotoxin gene (genotype IS1151-cpe or IS1470-like-cpe) as a common cause of food poisoning. J. Clin. Microbiol., 46: 371-373.
CrossRefDirect Link - Alfonzo, A., G. Ventimiglia, O. Corona, R. Di Gerlando and R. Gaglio et al., 2013. Diversity and technological potential of lactic acid bacteria of wheat flours. Food Microbiol., 36: 343-354.
CrossRefDirect Link - Enan, G., A.R. Al-Mohammadi, G. El-Didamony, M.E.F. Abdel-Haliem and A. Zakaria, 2014. Antimicrobial activity of Enterococcus faecium NM2 isolated from urine: Purification, characterization and bactericidal action of enterocin NM2. Asian J. Applied Sci., 7: 621-634.
CrossRefDirect Link - Sadek, Z.I., B.M. Refaat, E.H. Abd El-Shakour, N.S.H. Mehanna and M.S. Hassan, 2015. Potential sources of aerobic and anaerobic spore former bacteria in processed cheese. Res. J. Pharmaceut. Biol. Chem. Sci., 6: 757-769.
Direct Link - Lyon, W.J. and B.A. Glatz, 1993. Isolation and purification of propionicin PLG-1, a bacteriocin produced by a strain of Propionibacterium thoenii. Applied Environ. Microbiol., 59: 83-88.
Direct Link - Mayer, H.K., 2001. Bitterness in processed cheese caused by an overdose of a specific emulsifying agent? Int. Dairy J., 11: 533-542.
CrossRefDirect Link - Schar, W. and J.O. Bosset, 2002. Chemical and physico-chemical changes in processed cheese and ready-made fondue during storage: A review. LWT-Food Sci. Technol., 35: 15-20.
CrossRefDirect Link - Varadaraj, M.C., 1993. Methods for detection and enumeration of foodborne bacterial pathogens: A critical evaluation. J. Food Sci. Technol., 30: 1-13.
Direct Link - Jack, R.W., J.R. Tagg and B. Ray, 1995. Bacteriocins of gram-positive bacteria. Microbiol. Rev., 59: 171-200.
Direct Link - Ahmadova, A., S.D. Todorov, I. Hadji-Sfaxi, Y. Choiset and H. Rabesona et al., 2013. Antimicrobial and antifungal activities of Lactobacillus curvatus strain isolated from homemade Azerbaijani cheese. Anaerobe, 20: 42-49.
CrossRefDirect Link - Gillor, O., A. Etzion and M.A. Riley, 2008. The dual role of bacteriocins as anti- and probiotics. Applied Microbiol. Biotechnol., 81: 591-606.
CrossRefDirect Link - Zacharof, M.P. and R.W. Lovitt, 2012. Bacteriocins produced by lactic acid bacteria a review article. APCBEE Procedia, 2: 50-56.
CrossRefDirect Link - Rossland, E., T. Langsrud, P.E. Granum and T. Sorhaug, 2005. Production of antimicrobial metabolites by strains of Lactobacillus or Lactococcus co-cultured with Bacillus cereus in milk. Int. J. Food Microbiol., 98: 193-200.
CrossRefDirect Link - Mkrtchyan, H., S. Gibbons, M. Heidelberger, M. Zloh and H.K. Limaki, 2010. Purification, characterisation and identification of acidocin LCHV, an antimicrobial peptide produced by Lactobacillus acidophilus n.v. Er 317/402 strain Narine. Int. J. Antimocrob. Agents, 35: 255-260.
CrossRefDirect Link - Nazzaro, F., S. Pepe, T. Granese, A. Cozzolino, F. Cardinale and F. Fratianni, 2014. Presence of rocket salad (Eruca sativa) in the medium might affect the growth of Lactobacillus acidophilus and its biological properties. J. Prob. Health, Vol. 2, No. 2.
Direct Link - Van Netten, P., A. van de Moosdijk, P. van Hoensel, D.A.A. Mossel and I. Perales, 1990. Psychrotrophic strains of Bacillus cereus producing enterotoxin. J. Applied Bacteriol., 69: 73-79.
CrossRefDirect Link - Giannella, R.A. and L.A. Brasile, 1979. A hospital food-borne outbreak of diarrhea caused by Bacillus cereus: Clinical, epidemiologic and microbiologic studies. J. Infect. Dis., 139: 366-370.
CrossRefDirect Link - Rukure, G. and B.H. Bester, 2001. Survival and growth of Bacillus cereus during Gouda cheese manufacturing. Food Control, 12: 31-36.
CrossRefDirect Link - De Vuyst, L. and E.J. Vandamme, 1994. Nisin, a Lantibiotic Produced by Lactococcus lactis Subsp. lactis: Properties, Biosynthesis, Fermentation and Applications. In: Bacteriocins of Lactic Acid Bacteria: Microbiology, Genetics and Applications, De Vuyst, L. and E.J. Vandamme (Eds.). Blackie Academic and Professional, London, England, pp: 151-221.
- Trias, R., L. Baneras, E. Badosa and E. Montesinos, 2008. Bioprotection of golden delicious apples and iceberg lettuce against foodborne bacterial pathogens by lactic acid bacteria. Int. J. Food Microbiol., 123: 50-60.
CrossRefDirect Link - Calo-Mata, P., S. Arlindo, K. Boehme, T. de Miguel, A. Pascoal and J. Barros-Velazquez, 2008. Current applications and future trends of lactic acid bacteria and their bacteriocins for the biopreservation of aquatic food products. Food Bioprocess. Technol., 1: 43-63.
CrossRefDirect Link - Pirttijarvi, T.S.M., M.A. Andersson and M.S. Salkinoja-Salonen, 2000. Properties of Bacillus cereus and other bacilli contaminating biomaterial-based industrial processes. Int. J. Food Microbiol., 60: 231-239.
CrossRefDirect Link - Kumar, A.M. and S. Arumugam, 2012. Impact of culture conditions on the production of curvacin a by Lactobacillus curvatus LC05. Int. J. Pharma Bio Sci., 3: 427-432.
Direct Link - Ennahar, S., T. Sashihara, K. Sonomoto and A. Ishizaki, 2000. Class IIa bacteriocins: Biosynthesis, structure and activity. FEMS Microbiol. Rev., 24: 85-106.
CrossRefDirect Link - Devi, S.M. and P.M. Halami, 2013. Conjugal transfer of bacteriocin plasmids from different genera of lactic acid bacteria into Enterococcus faecalis JH2-2. Ann. Microbiol., 63: 1611-1617.
CrossRefDirect Link