
Jhy- Ming Li
Department of Animal Science, National Chiayi University, Chiayi, Taiwan
Shu- Yu Hsieh
Department of Animal Science, National Chiayi University, Chiayi, Taiwan
Yan- Ming Horng
Department of Animal Science, National Chiayi University, Chiayi, Taiwan
Chean- Ping Wu
Department of Animal Science, National Chiayi University, Chiayi, Taiwan
International Journal of Dairy Science
Year: 2017 | Volume: 12 | Issue: 1 | Page No.: 41-46







ABSTRACT
Objective: This study aimed to investigate αs1-casein (CSN1S1) gene polymorphisms in Taiwan dairy goats by using PCR-restriction fragment length polymorphism (PCR-RFLP) in combination with allele-specific-PCR (AS-PCR). Methodology: One hundred and sixty Taiwan dairy goats were used to detect the presence of CSN1S1 gene polymorphisms. After DNA amplification, the PCR products were digested with XmnI restriction enzyme or were directly subjected to gel electrophoresis. Results: The following 11 CSN1S1 genotypes were identified: AA, AE, AF, A01, EE, EF, EN, E01, FF, F01 and N01 with frequencies of 0.13, 0.13, 0.29, 0.02, 0.02, 0.19, 0.01, 0.03, 0.13, 0.04 and 0.01, respectively. Among the CSN1S1 alleles, the frequency of the F allele (0.40) was the highest, indicating that the CSN1S1 F allele was the most common allele in all examined Taiwan dairy goats. Conclusion: In conclusion, a quick and accurate technique was used for typing goats independently of CSN1S1 genotype. By examining CSN1S1 gene polymorphisms, we could identify breeds carry a high percentage of homozygous CSN1S1 F allele for use in the fresh milk industry in Taiwan.
PDF Abstract XML References Citation
Received: August 10, 2016;
Accepted: October 20, 2016;
Published: December 15, 2016
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Jhy- Ming Li, Shu- Yu Hsieh, Yan- Ming Horng and Chean- Ping Wu, 2017. Genotyping of the CSN1S1 Locus in Taiwan Dairy Goats using PCR-RFLP and AS-PCR. International Journal of Dairy Science, 12: 41-46.
DOI: 10.3923/ijds.2017.41.46
URL: https://scialert.net/abstract/?doi=ijds.2017.41.46
DOI: 10.3923/ijds.2017.41.46
URL: https://scialert.net/abstract/?doi=ijds.2017.41.46
INTRODUCTION
Casein is the main protein fraction of ruminant milk and is one of the most valuable components owing to its nutritional value and processing properties. The casein fraction is encoded by four tightly linked genes, αs1-casein (CSN1S1), αs2-casein (CSN1S2), β-casein (CSN2) and κ-casein (CSN3), which cluster in a genomic DNA segment of approximately 250 kb1-3. The goat CSN1S1 gene consists of 19,400 nucleotides, with 19 exons. The CSN1S1 protein contains 214 amino acids, including a 15-amino acid signal peptide and 199 functional amino acids. The goat CSN1S1 locus is characterized by at least 16 alleles and various CSN1S1 alleles can produce different levels of CSN1S1 in caprine milk. The alleles A, B1, B2, B3, B4, C, H, L and M are related to a high content of CSN1S1, whereas alleles I and E are associated with an intermediate content and alleles D, F and G with a low content. Alleles 01 and 02 are ‘null’ alleles associated with the absence of CSN1S14-10. Moreover, the F allele is derived from a deletion of the 23rd nucleotide of the 9th exon and insertions of 11 and 3 bp within the 9th intron11. The N allele is derived from the F allele via an exonic mutation at nucleotide 1319 of the F allele promoter region, which creates an extra putative activator protein (AP-1) binding motif but without insertions of 11 bp and 3 bp within the 9th intron of the F allele sequence8. The N allele has also been identified at the CSN1S1 locus and is associated with lack of CSN1S1 synthesis12.
In some countries, caprine milk is produced for cheese making; however, in Taiwan it is consumed directly. The caprine milk industry in Taiwan mainly uses hybrid breeds dominant for milk production, with an emphasizes on the heat stability of caprine milk. Therefore, the potential for improving caprine milk stability properties, such as heat stability, by selecting for low CSN1S1 milk producers, might be significant. We aimed to investigate the combined use of PCR-restriction fragment length polymorphism (PCR-RFLP) and allele-specific-PCR (AS-PCR) to characterize allelic polymorphisms of the CSN1S1 locus.
MATERIALS AND METHODS
Sample collection and DNA isolation: The research was carried out on 160 individual DNA samples from local Taiwan dairy goats that were mainly of a Sanan×Alpine×Nubia cross raised on the animal research farm of National Chiayi University and in Zhongpu in Chiayi country, Taiwan. Genomic DNA was extracted from whole blood using the phenol-chloroform method with minor modifications. Ten milliliters of blood taken on EDTA was mixed with 25 mL cold sucrose-triton and the volume was made up to 50 mL with autoclaved double-distilled water. The solution was mixed well and a nuclear pellet was obtained by centrifugation for 10 min at 3,000 rpm and discarding the supernatant. The nuclear pellet was suspended in lysis buffer (10 mM tris base, 150 mM NaCl and 10 mM sodium EDTA) pH 8.2, with 10% Sodium Dodecyl Sulfate (SDS), 50 μL proteinase K (10 mg mL–1) and 40 μL colleagenase (3.8 IU μL–1) and incubated overnight in a shaking water-bath at 45°C.
Nucleic acid was extracted once with phenol saturated with tris-EDTA (TE) buffer (10 mM tris, 10 mM NaCl and 1 mM EDTA), followed by extraction with phenol:chloroform: isoamyl alcohol (25:24:1) until there was no protein at the interface. This was followed by extraction with chloroform: isoamyl alcohol (24:1). To each extraction mixture, an equal volume of the solvent was added, followed by thorough mixing and centrifugation for 10 min at 2000 rpm. The top layer was transferred to a fresh Falcon tube for the next extraction. To the final aqueous phase, a 0.1 volume of 2.5 M sodium acetate and 2.5 volumes of cold 95% ethanol were added. The tubes were agitated gently to mix the liquids and a fluffy white ball of DNA was formed. The DNA was picked up with a heat-sealed Pasteur pipette and washed briefly in ethanol. The DNA was finally dissolved in an appropriate volume of 1×TE buffer. The DNA concentrations were determined and diluted to a working concentration of 25 ng μL–1, which is suitable for polymerase chain reactions.
Polymerase Chain Reaction (PCR): The PCR-RFLP primers used in the present study were synthesized from the published nucleotide sequence of the goat αs1-casein gene: (forward: 5-TTCTAAAAGTCTCAGAGGCAG-3), (reverse: 5-GGGTTGATAGCCTTGTATGT-3)13. Similarly, the AS-PCR primers for CSN1S1E were synthesized from the published nucleotide sequence of the goat αs1-casein gene: (forward: 5-TCAGGAGCAGTGGGTATGTG-3), (reverse: 5-CCTCCCAATGG AATAATGACA-3 )14. The AS-PCR primers for CSN1S101 were synthesized from the published nucleotide sequence of the goat αs1-casein gene: (forward: 5 -CCCCAGCTGGTAATGTTTTA-3), (reverse: 5-GGTCCATCAA TTCCCTGTGT-3)14.
The PCR reaction mixtures consisted of 1.0 μM forward and reverse primers, 0.2 mM dNTPs, 10 mM tris (pH 9), 50 mM KCl, 1.5 mM MgCl2, 0.01% gelatin (w/v), 0.1% triton X-100 and 1.25 U of Taq polymerase. The mixture was aliquoted into tubes containing 100 ng goat DNA. The reaction mixture was overlaid with sterile mineral oil and amplified in a Coy Temp Cycler II (Coy Corporation, MI, USA), using the following amplification protocol: 30 cycles of 94°C for 1 min, 60°C for 1 min and 72°C for 2 min.
PCR-RFLP, AS-PCR and agarose gel electrophoresis: Twenty microliters of each PCR product was digested with 10 U of XmnI restriction enzyme in a final reaction volume of 25 μL. The reaction mixture was incubated at 37°C in a water bath overnight. After restriction digestion, the restricted fragments were analyzed by electrophoresis on a 0.4% agarose/1×TBE gel stained with ethidium bromide. A 100 bp ladder was used as a molecular size marker. The bands were visualized under UV light and photographed with a yellow filter using black and white film.
RESULTS
It has been reported that the F allele is derived from a C deletion at the 23rd nucleotide of the 9th exon and short insertions of 11 and 3 bps in the subsequent intron11. Furthermore, the gene mutation of the F allele could be digested using the XmnI restriction enzyme13. Additionally, a further report indicated that the E allele has a repeat fragment (approximately 457 bp) in exon 1915. Therefore, we used PCR-RFLP in combination with AS-PCR to analyze the CSN1S1 polymorphisms of a population of Taiwan dairy goats. Eight RFLPs were obtained after digesting the PCR product with XmnI (bp: 223, 223+161+63, 223+150+63, 212+161+63, 212+150+63, 161+63, 161+150+63 and 150+63) (Fig. 1). Our data showed that PCR-RFLP in combination with AS-PCR facilitated identification of the genotypes F/F at 223 bp, N/N at 212 bp, A/A with two fragments at 161 and 63 or 150 and 63 bp, E/E at 1040 bp and 01/- at 249 bp (Fig. 1-3).
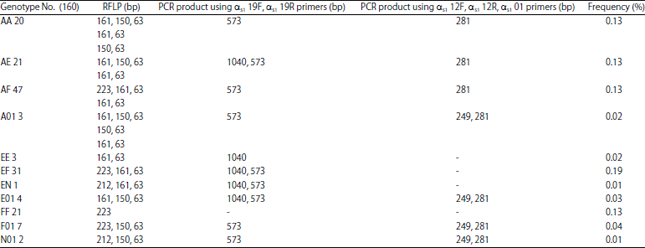
Samples from the 160 Taiwan dairy goats were further examined for the presence of 11 polymorphisms of goat CSN1S1. We detected these 11 CSN1S1 polymorphisms in the following number of goats: 20 goats (AA), 21 (AE), 47 (AF), 3 (A01), 3 (EE), 31 (EF), 1 (EN), 4 (E01), 21 (FF), 7 (F01) and 2 (N01) (Table 1). Our data showed that the presence of A/A is indicated by three fragments at 161, 150 and 63 bp, A/E by four fragments at 161, 150, 63 and 1040 bp, A/F by four fragments at 223, 161, 150 and 63 bp, A01 by four fragments at 161, 150, 63 and 249 bp, EF by four fragments at 223, 161, 63 and 1040 bp, EN by four fragments at 212, 161, 63 and 1040 bp, E01 by five fragments at 161, 150, 63, 1040 and 249 bp, F01 by four fragments at 223, 150, 63 and 249 bp and N01 by four fragments at 212, 150, 63 and 249 bp. The genotype frequencies of AA, AE, AF, A01, EE, EF, EN, E01, FF, F01 and N01 were 0.13, 0.13, 0.29, 0.02, 0.02, 0.19, 0.01, 0.03, 0.13, 0.04 and 0.01, respectively. The highest genotype frequency in Taiwan dairy goats was that of AF, whereas the lowest were EN and N01. Table 2 shows that A, E, F, N and 01 alleles were present at the CSN1S1 locus on genomic DNA in Taiwan dairy goats. The allelic frequencies of A, E, F, N and 01 were 0.35, 0.2, 0.4, 0.04 and 0.01, respectively. Our results indicate that the frequency of the CSN1S1 F allele was highest and that of the 01 allele was the lowest in Taiwan dairy goats.
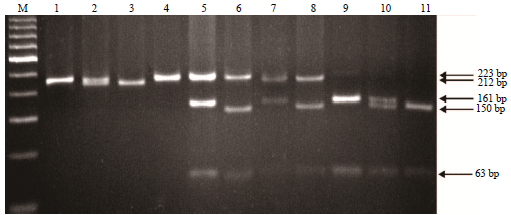 | |
| Fig. 1: | DNA electrophoretic patterns obtained from Taiwan dairy goat αS1-casein (CSN1S1) after PCR amplification and digestion with XmnI, M: 100 bp, Lane 1: Control 1 (223 bp), Lane 2: Control 2 (223 and 212 bp), Lane 3: Control 3 (212 bp), Lane 4: Homozygous FF genotype (223 bp), Lane 5: Heterozygous AF and EF genotype (223, 161 and 63 bp), Lane 6: Heterozygous AF and F01 genotype (223, 150 and 63 bp), Lane 7: Heterozygous EN genotype (212, 161 and 63 bp), Lane 8: Heterozygous N01 genotype (212, 150 and 63 bp), Lane 9: Heterozygous AA, AE, A01 and EE genotype (161 and 63 bp), Lane 10: Heterozygous AA, AE, A01 and E01 genotype (161, 150 and 63 bp), Lane 11: Heterozygous AA and A01 genotype (150 and 63 bp) |
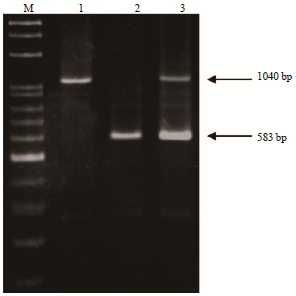 | |
| Fig. 2: | DNA electrophoretic patterns obtained by PCR amplification of Taiwan dairy goat αS1-casein E allele, M: 100 bp, Lane 1: Homozygous EE genotype (1040 bp), Lane 2: Heterozygous AA, AF, A01, F01 and N01 genotype (583 bp), Lane 3: Heterozygous AE, EF, EN and E01 genotype (1040 and 583 bp) |
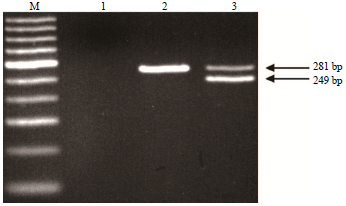 | |
| Fig. 3: | DNA electrophoretic patterns obtained by PCR amplification of Taiwan dairy goat αS1-casein 01 allele. M: 100-bp, Lane 1: Negative control, Lane 2: Non CSN1S101, Lane 3: Heterozygous A01, E01, F01 and N01 genotype (281 and 249 bp) |
| Table 1: | Genotype frequency of αs1-casein from Taiwan dairy goat DNA determined using PCR-restriction fragment length polymorphism and allele-specific-PCR |
 | |
| Table 2: | Allelic frequencies of αs1-casein in 160 Taiwan dairy goats |
 | |
DISCUSSION
It has been shown that the CSN1S1 locus is characterized by at least 13 alleles and is associated with different levels of caprine milk protein. Cosenza et al.16 have reported that using HphI, DdeI and MnlI restriction enzymes with PCR-RFLP yielded alleles B1, B2, B3 and C of CSN1S1. Additionally, the reports have also indicated that PCR-RFLP is used to analyze CSN1S1 and CSN3 genotypes for other animals17-19. However, PCR-RFLP is not suitable for detecting specific casein alleles and thus AS-PCR can be used to compensate for the deficiencies of PCR-RFLP16,20. Accordingly, in the present study, it was investigated whether the combination of PCR-RFLP with AS-PCR could be used as a rapid method to analyze a larger number of CSN1S1 gene polymorphisms in Taiwan dairy goats. Our results showed that 11 genotypes and five alleles could be detected by using RFLP-PCR in combination with AS-PCR (Table 1, 2). The allelic frequency of F and 01 were highest and lowest, respectively, in Taiwan dairy goats. Because the Taiwan dairy goats we examined were mainly Sanan×Alpine×Nubia hybrids, they are comparable with European goats. Additionally, we made the interesting discovery that the 01 allele (null) is present in Taiwan dairy goats but not in European and Indian goats16,20,21.
Previous studies have indicated that the amount of CSN1S1 is positively correlated with the presence of the various CSN1S1 alleles22-24. The amount of CSN1S1 mRNA transcribed from the F allele was at least 6 times lower than that transcribed from the A allele11. The weak F allele (0.6 g L–1) is most frequent in both Alpine and Sanan goats1,25, which is consistent with our population. Furthermore, characterization of CSN1S1 variability is important owing to its relationship with direct consumption and dairy production. The allelic frequencies of F and 01 are important in terms of a low content of CSN1S1 in caprine milk. It is therefore possible that the F and 01 alleles were selected in populations of Taiwan dairy goats.
Previous studies have also indicated a positive correlation between good cheese-making properties and a high content of CSN1S1; however, caprine milk contains a low content of CSN1S1 with poor stability properties4,26. Therefore, caprine milk is more suitable for use as fresh milk in Taiwan. Moreover, milk with high levels of CSN1S1 has higher total protein, casein, total solids and lower pH than caprine milk with low levels of CSN1S127. Singh28 further indicated that a lower pH would affect the heat stability of milk. If caprine milk contains a high casein content and satisfactory rennet stability, this is likely to affect the production of caprine milk. Selection of Taiwan dairy goats that produce milk with low solid content is recommended for those who are interested in producing and improving the heat stability of caprine milk for the caprine milk industry in Taiwan.
CONCLUSION
In cattle, the CSN1S1 B1, B2, B3, B4 and C alleles have been genotyped using isoelectric focusing (IEF), however, this method is slow and leads to ambiguous typing of individuals when using caprine milk. Here, we developed a quick, cheap and accurate technique for typing goats independently of CSN1S1 genotype. Our results indicate that Taiwan dairy goats possess high levels of the CSN1S1 F allele. We also discovered that the 01 allele (null) is present in Taiwan dairy goats but not in European and Indian goats. Therefore, breeding Taiwan dairy goats with F and 01 (null) alleles, which results in the production of milk with low solid content is significant for the caprine milk industry.
ACKNOWLEDGMENTS
We thank the Animal Research Farm, National Chiayi University, Chiayi, Taiwan for providing the apparatus, facilities and animal care required in this study. Dr. Li, J.M., Dr. Wu, C.P. and Dr. Horng, Y.M. designed the experiments, edited of manuscript and wrote-revision. Hsieh, S.Y. extracted the DNA of blood, digested by PCR-RFLP and AS-PCR.
REFERENCES
- Grosclaude, F., M.F. Mahe, G. Brignon, L. di Stasio and R. Jeunet, 1987. A mendelian polymorphism underlying quantitative variations of goat αs1-casein. Genet. Sel. Evol., 19: 399-412.
CrossRefDirect Link - Ferretti, L., P. Leone and V. Sgaramella, 1990. Long range restriction analysis of the bovine casein genes. Nucl. Acids Res., 18: 6829-6833.
Direct Link - Rijnkels, M., 2002. Multispecies comparison of the casein gene loci and evolution of casein gene family. J. Mammary Gland Biol. Neoplasia, 27: 327-345.
PubMedDirect Link - Clark, S. and J.W. Sherbon, 2000. Alphas1-casein, milk composition and coagulation properties of goat milk. Small Rumin. Res., 38: 123-134.
CrossRefDirect Link - Bevilacqua, C., P. Ferranti, G. Garro, C. Veltri and R. Lagonigro et al., 2002. Interallelic recombination is probably responsible for the occurrence of a new αs1‐casein variant found in the goat species. EFBS J., 269: 1293-1303.
CrossRefDirect Link - Jordana, J., M. Amills, E. Diaz, C. Angulo, J.M. Serradilla and A. Sanchez, 1996. Gene frequencies of caprine αs1-casein polymorphism in Spanish goat breeds. Small Rumin. Res., 20: 215-221.
CrossRefDirect Link - Martin, P., M. Ollivier-Bousquet and F. Grosclaude, 1999. Genetic polymorphism of caseins: A tool to investigate casein micelle organization. Int. Dairy J., 9: 163-171.
Direct Link - Ramunno, L., G. Cosenza, A. Rando, A. Pauciullo and R. Illario et al., 2005. Comparative analysis of gene sequence of goat CSN1S1 F and N alleles and characterization of CSN1S1 transcript variants in mammary gland. Gene, 345: 289-299.
CrossRefDirect Link - Zullo, A., C.M.A. Barone, L. Chianese, P. Colatruglio, M. Occidente and D. Matassino, 2005. Protein polymorphisms and coagulation properties of Cilentana goat milk. Small Rumin. Res., 58: 223-230.
CrossRefDirect Link - Rodrigues, M.T., M.A.M. Soares, A.A. Zacaro, M.M.C. Silva, O.S.R. Garcia and A.C.M. Magalhaes, 2015. Differences in the defective alleles E and F for the locus CSN1S1 in goats affects the profile of milk caseins. Small Rumin. Res., 123: 47-54.
CrossRefDirect Link - Leroux, C., N. Mazure and P. Martin, 1992. Mutations away from splice site recognition sequences might cis-modulate alternative splicing of goat alpha s1-casein transcripts. Structural organization of the relevant gene. J. Biol. Chem., 267: 6147-6157.
Direct Link - Ramunno, L., G. Cosenza, M. Pappalardo, N. Pastore, D. Gallo, P. Di Gregorio and P. Masina, 2000. Identification of the goat CSN1S1F allele by means of PCR-RFLP method. Anim. Genet., 31: 342-342.
CrossRefDirect Link - Dettori, M.L., G.M. Vacca, V. Carcangiu, M. Pazzola, M.C. Mura and A.M. Rocchigiani, 2009. A reliable method for characterization of the goat CSN1S1 E allele. Livest. Sci., 125: 105-108.
CrossRefDirect Link - Perez, M.J., C. Leroux, A.S. Bonastre and P. Martin, 1994. Occurrence of a LINE sequence in the 3'UTR of the goat αs1-casein E-encoding allele associated with reduced protein synthesis level. Gene, 147: 179-187.
CrossRefDirect Link - Cosenza, G., A. Pauciullo, D. Gallo, L. Colimoro, A. D'Avino, A. Mancusi and L. Ramunno, 2008. Genotyping at the CSN1S1 locus by PCR-RFLP and AS-PCR in a Neapolitan goat population. Small Rumin. Res., 74: 84-90.
CrossRefDirect Link - El Nahas, S.M. and A.A. Mossallam, 2014. AcuI identifies water buffalo CSN3 genotypes by RFLP analysis. J. Genet., 93: e94-e96.
PubMedDirect Link - Erhardt, G., E.T.S. Shuiep, M. Lisson, C. Weimann, Z. Wang, L.Y. El Zubeir and A. Pauciullo, 2016. Alpha S1-casein polymorphisms in camel (Camelus dromedarius) and descriptions of biological active peptides and allergenic epitopes. Trop. Anim. Health Prod., 48: 879-887.
CrossRefDirect Link - Selvaggi, M., A.G. D'Alessandro and C. Dario, 2015. Comparative characteristics of DNA polymorphisms of κ-casein gene (CSN3) in the horse and donkey. Genet. Mol. Res., 14: 14567-14575.
Direct Link - Kumar, A., P.K. Rout, A. Mandal and R. Roy, 2007. Identification of the CSN1S1 allele in Indian goats by the PCR-RFLP method. Animal, 1: 1099-1104.
CrossRefDirect Link - Kusza, S., G. Veress, S. Kukovics, A. Javor, A. Sanchez, A. Angiolillo and Z. Bosze, 2007. Genetic polymorphism of αs1-and αs2-caseins in Hungarian milking goats. Small Rumin. Res., 68: 329-332.
CrossRefDirect Link - Roncada, P., A. Gaviraghi, S. Liberatori, B. Canas, L. Bini and G. Greppi, 2002. Identification of caseins in goat milk. Proteomics, 2: 723-726.
CrossRefPubMedDirect Link - Vacca, G.M., M.L. Dettori, G. Piras, F. Manca, P. Paschino and M. Pazzola, 2014. Goat casein genotypes are associated with milk production traits in the Sarda breed. Anim. Genet., 45: 723-731.
CrossRefDirect Link - Pirisi, A., O. Colin, F. Laurent, J. Scher and M. Parmentier, 1994. Comparison of milk composition, cheesemaking properties and textural characteristics of the cheese from two groups of goats with a high or low rate of αS1-casein synthesis. Int. Dairy J., 4: 329-345.
CrossRefDirect Link - Ambrosoli, R., L. di Stasio and P. Mazzocco, 1988. Content of αs1-casein and coagulation properties in goat milk. J. Dairy Sci., 71: 24-28.
CrossRefDirect Link