
Marciele Bogo
Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Karine Lauer Cruz
Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Alvaro Gonzalez Revello
Instituto de Ciencia y Tecnolog�a de los Alimentos de Origen Animal, Departamento de Ciencia y Tecnolog�a de la Leche, Facultad de Veterinaria, UdelaR, Montevideo, Uruguay
Ana Paula Folmer Correa
Laboratorio de Microbiologia Aplicada, Instituto de Ciencia e Tecnologia de Alimentos, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Adriano Brandelli
Laboratorio de Microbiologia Aplicada, Instituto de Ciencia e Tecnologia de Alimentos, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Ana Paula Guedes Frazzon
Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
Amanda de Souza da Motta
Departamento de Microbiologia, Imunologia e Parasitologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brazil
LiveDNA: 55.16387
International Journal of Dairy Science
Year: 2017 | Volume: 12 | Issue: 5 | Page No.: 339-347







ABSTRACT
Background and Objective: Psychrotrophic bacteria produce extracellular proteases, resulting in deterioration and reduced shelf life of dairy products. In this study, 21 species of psychotropic bacteria isolated from buffalo milk were selected and the thermal resistance of the proteases produced by these bacteria was evaluated. Materials and Methods: The isolates were tested to evaluate proteolytic activity of buffalo milk agar. The cell-free supernatants from the growing of isolates were obtained for the quantification of enzymatic activity under different pH values (5.5, 7.0 and 8.0). Thermal resistance and the clotting ability of proteolytic enzymes in buffalo and bovine milk substrates were also evaluated. One-way ANOVA test with a critical probability of p<0.05 followed by the Tukey’s test was used to evaluate the results. Results: All strains were able to produce proteolysis in buffalo milk agar; additionally, all cell-free supernatants showed enzymatic activity, with values of >1 U mL–1 under at least one of the pH tested. Five isolates produced cell-free supernatants resistant to pasteurization (63.5°C/30 min), following which they were able to coagulate buffalo and bovine milk. The crude enzyme of P. fluorescens PL5.4 showed the greatest enzymatic activity within a wide pH range (4-10) and at an optimum temperature of 40°C. The cell-free supernatant of this isolate resisted to tests with detergents and organic solvents. However, it was not possible to identify the type of protease. Conclusion: The results of this study showed the negative impact of the presence of psychrotrophic bacteria producing proteolytic enzymes in buffalo milk. This is because the enzymes studied caused changes in milk samples, revealing a negative impact on the production of derived products. This is significant, since the buffalo milk produced in Brazil is directed to the production of dairy products.
PDF Abstract XML References Citation
Received: June 16, 2017;
Accepted: July 15, 2017;
Published: August 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Marciele Bogo, Karine Lauer Cruz, Alvaro Gonzalez Revello, Ana Paula Folmer Correa, Adriano Brandelli, Ana Paula Guedes Frazzon and Amanda de Souza da Motta, 2017. Thermal Resistance of Proteolytic Enzymes Produced by Psychrotrophic Bacteria Isolated from Buffalo Milk. International Journal of Dairy Science, 12: 339-347.
DOI: 10.3923/ijds.2017.339.347
URL: https://scialert.net/abstract/?doi=ijds.2017.339.347
DOI: 10.3923/ijds.2017.339.347
URL: https://scialert.net/abstract/?doi=ijds.2017.339.347
INTRODUCTION
Buffalo milk presents higher levels of total solids such as crude protein, fat, calcium, phosphorous and slightly higher lactose content when compared with cow milk. However, the consumption of liquid buffalo milk in Brazil is still not widespread although the high level of total solids makes buffalo milk ideal for the processing of dairy products such as cheese1. It is especially used in Mozzarella cheese manufacture and there is no need to concentrate it prior to processing2. It is important to consider that milk is an ideal growth medium for many microorganisms, due to its high nutritional value, high water activity and neutral pH3. A number of microorganisms present in raw milk have a significant impact on the final quality of dairy products. Importantly, the presence of undesirable bacteria, such as spoilage and pathogens, can lead to the loss of organoleptic properties of dairy products or even compromise the general safety of these products. Undesirable microorganisms enter raw milk from a variety of sources, including the teat apex, milking equipment, air, water, feed, grass, soil, the environment and operators4.
The microbiota of raw milk is very diverse and among these contaminating microorganisms, the psychrotrophic bacteria represent a noteworthy group because of their ability to grow under refrigeration temperatures3. Their potential to thrive under cooler conditions leads to reduced milk quality, thus compromising the production of dairy products5. These microorganisms typically represent from 37-54% of the microbiota present in stored refrigerated milk6.
The importance of psychrotrophic bacteria is also due to their production of extracellular enzymes such as proteases that are heat stable and remain active over a broad range of temperatures7. Heat treatment such as pasteurization and Ultra-High Temperature (UHT) are not sufficient to inactivate some bacterial proteases8. The enzymatic profile of these bacteria often causes the degradation of caseins in dairy products, especially k-casein, thus giving the product a bitter taste and a sour smell during storage9.
Proteases can be produced by a variety of bacteria including species of Bacillus, Pseudomonas, Burkholderia, Chryseobacterium and Serratia, which are all found in dairy environments10,11. The proteolytic enzymes are of significant concern for the dairy industry, as they can remain active in milk products during prolonged storage, even at low temperatures7. Understanding the consequences of the presence of these enzymes in milk is essential in order to propose strategies to reduce the transfer of enzyme-producing bacteria.
With the increased consumption of products derived from buffalo milk, the study and evaluation of the quality of the raw material have become more imperative. It is also important to highlight that in the state of Rio Grande do Sul-Brazil, 100% of buffalo milk produced is directed towards the preparation of dairy products. Thus, the present research will contribute to the generation of data regarding the proteolytic capacity of the isolated psychrotrophic bacteria of this raw material, considering that there is little information on buffalo milk in the literature. There is no legislation for this milk in the State of Rio Grande do Sul, Brazil. Thus, this study aimed to evaluate the production of proteolytic enzymes by psychrotrophic bacteria isolated from raw buffalo milk, as well as the possible impact of the presence of these proteases in milk.
MATERIALS AND METHODS
The study was carried out with whole buffalo raw milk collected in a dairy in the Southern region of Brazil. The development of the tests was carried out at the Laboratory of Microbiology of the Department of Microbiology, Immunology and Parasitology of the Institute of Basic Health Sciences of the Federal University of Rio Grande do Sul during the years 2015 and 2016.
Bacterial cultures and growing conditions: For this research, 21 cultures of psychrotrophic bacteria were studied and previously isolated from samples of refrigerated raw buffalo milk and maintained at -20°C in cryotubes containing 10% (v/v) skim milk and 20% (v/v) glycerol (Scientific Hexis). For the reactivation of the isolates, Tryptic Soy Broth medium (TSB, Himedia) was used and incubation occurred at a temperature of 30°C for 48 h. These cultures were inoculated on Tryptone Soy Agar plates (TSA, Himedia) for 48 h in order to observe the purity of the isolates (Table 1).
Evaluation of the proteolytic activity of psychrotrophic bacteria: Proteolytic activity was evaluated as described by Ruaro et al.12. Bacterial cultures were inoculated in buffalo milk agar (5 g L–1 meat peptone, 3 g L–1 yeast extract, 12 g L–1 agar and 10% sterilized buffalo milk) and incubated at 30°C for 48 h. Pseudomonas aeruginosa ATCC 27853 was used as a positive control. Proteolytic activity was observed when a clear halo appeared around the colony and the results were expressed in millimeters (mm).
Evaluation of the thermal stability of proteolytic enzymes: The 21 cultures of psychrotrophic bacteria were grown in 50 mL of sterile milk and incubated at 30°C for 72 h in a shaking incubator at 180 rpm (Certomat® BS-1, Sartorius Stedim Biotech GmbH, Göttingen, Germany).
| Table 1: | Proteolytic evaluation in buffalo milk agar and enzymatic activity of psychrotrophic bacteria isolated from refrigerated raw buffalo milk |
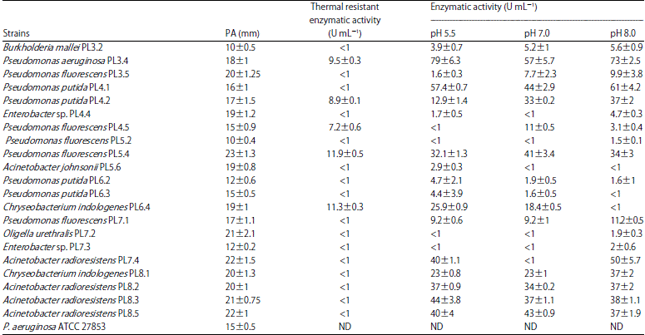 | |
| Values are Means±Standard Deviations. PA: Proteolytic activity; ND: Not determined | |
A volume of each culture was centrifuged at 1.900 rpm for 10 min and the cell-free supernatant was filtered through a membrane filter of 0.22 μm (Millipore). A volume of 1 mL of the crude enzyme was heat treated at pasteurization temperature (63.5°C for 30 min) in thermal drying equipment (Dryblock, Novatécnica, Piracicaba, São Paulo, Brazil)13. Another 1 mL aliquot remained unheated. The enzymatic quantification was performed using azocasein (Sigma) as a substrate, essentially as described by Daroit et al.13.
Milk clotting evaluation: A volume of 1 mL of the crude enzyme from each psychrotrophic bacteria, was mixed with 2 mL of pasteurized bovine milk and another experiment was also conducted with 2 mL pasteurized buffalo milk. Subsequently, the addition of 110 μL of 10 mg mL–1 sodium azide (Merck, Darmstadt, Germany) was made to ensure the inactivation of bacterial cells. The unheated crude enzymes were used as a control. Changes in the inoculated milk were evidenced by visual observation of coagulation over 6, 18, 24, 48, 72, 96 and 120 h incubation at 7 and 30°C, respectively. As a negative control, milk incubation was carried out only with the addition of sodium azide and the positive control was done with commercial chymosin (Sigma) enzyme14.
Quantification of proteolytic activity: The enzyme production was performed on a mineral medium (0.5 g L–1 NaCl, 0.3 g L–1 KH2PO4, 0.4 g L–1 K2HPO4) containing 1 g L–1 casein, according to Nornberg et al.15. The incubation was done at 30°C, 180 rpm for 72 h. After incubation, the inoculum (15 mL) was centrifuged at 1.900 rpm/10 min in order to produce a cell-free supernatant, considered as a crude enzyme. The quantification of the proteolytic activity was determined as described by Daroit et al.13. The absorbance at 420 nm was measured using a spectrophotometer (Biospectro SP-220, Curitiba, Paraná, Brasil). One unit of enzyme was defined as the amount of enzyme required for increasing 0.01 the absorbance at 420 nm under the test conditions.
Characterization of the stability of the crude enzyme: Partial characterization of enzymatic activity was carried out with the psychrotrophic bacterium that had the highest enzymatic activity values in tests done earlier. The methodology was performed according to Mateos et al.16. All assays were performed in triplicate. Different pH values were tested in the range of 4-11 (pH 4 and 5 with 0.2 M acetate buffer; pH 6, 7 and 8 with 0.2 M phosphate buffer; pH 9, 10 and 11 with 0.2 M carbonate-bicarbonate buffer, Sigma). The optimal temperature for enzymatic activity were carried out at temperatures of 30, 37, 40, 50, 60, 70 and 80°C in 0.2 M phosphate buffer pH 7.0. The effect of different metal ions was tested at 1 and 5 mM with the following salts: MgCl2, CaCl2, NaCl, CuCl2, CoCl2, MnCl2, ZnSO4, MgSO4, Al2 (SO4)3 and FeSO4. Sodium sulfate dodecyl (SDS), Triton X-100, Tween 20, dimethylsulfoxide (DMSO), isopropanol and acetonitrile were tested at 0.5 and 1%. The protease inhibitors ethylenediaminetetra acetic acid (EDTA) and phenylmethylsulfonyl fluoride (PMSF) were evaluated at both 1 and 5 mM. For these tests, the crude enzyme was pre-incubated for 10 min at 40°C and the proteolytic activity was determined with the addition of azocasein as described above. Enzyme activity determined without additions was considered as 100%. Furthermore, the thermal stability was evaluated by incubating the crude enzyme at temperatures of 40, 50, 60, 70, 80, 90 and 100°C for 10, 20, 30, 45, 60, 90 and 120 min. In each assay, a negative-enzyme control was used, replacing the sample with 0.2 M phosphate buffer pH 7.0. The reagents used were of the Sigma brand.
Statistical analysis: The tests used in the experiment were the same as those performed by Teh et al.17. Three replicates were performed for each experiment, on 2 separate occasions. Statistic 12.5 Software was used for analysis of variance using one-way ANOVA test with a critical probability of p<0.05 followed by Tukey’s test17.
RESULTS
Evaluation of the proteolytic activity of psychrotrophic bacteria: It was observed that 21 isolates presented a degradation area around the colony growth, indicating proteolysis. It was possible to measure clear halos, with values ranging from 10-23 mm. Some isolates produced larger halos when compared to those of the positive control (Table 1).
Evaluation of the thermal stability of proteolytic enzymes: It was observed that the proteolytic enzymes from 5 psychrotropic bacteria maintained their activity after heating, as noted with values ranging from 7.2-11.9 U m L–1. Four of these bacteria belong to the genus Pseudomonas while one of the isolates belongs to Chryseobacterium genus. The isolated P. fluorescens PL5.4 presented the highest value (Table 1).
Evaluation of enzyme activity at different pH values: The optimum pH for the enzyme activity varied among isolates as seen in Table 1. The maximum enzyme activity has been found in an isolate of P. aeruginosa PL3.4, at pH 5.5. This isolate presents the higher values of proteolytic activity even at pH 7.0 (a value close to milk pH) and pH 8.0. Most strains showed maximum values of enzymatic activity at pH 8.0 and only three strains presented activity lower than 1 U mL–1 at this pH. In addition, 11 psychrotrophic strains showed enzymatic activity at the 3 different pH values tested.
Coagulation of bovine and buffalo milk: It was observed that heat-treated enzymes from 5 isolates were capable of promoting total coagulation of both kinds of milk at the end of 5 days at different temperatures (Table 2). The enzymes from other psychrotrophic isolates evaluated in the study did not cause milk clotting after heat treatment (63.5°C for 30 min). The crude enzymes of the P. fluorescens strain PL4.5 coagulated bovine milk firstly while those of the P. fluorescens strain PL5.4 partially coagulated bovine milk. Furthermore, the crude enzymes of strains Chryseobacterium sp. PL6.4 and P. fluorescens PL5.4, caused total coagulation of bovine milk. At 30°C, the crude enzymes of P. fluorescens PL5.4 and Chryseobacterium sp. PL6.4 showed total coagulation. The coagulation profiles of buffalo milk at 7°C were similar to that observed for bovine milk when considered the same 5 isolates. The crude enzymes of P. fluorescens PL5.4 and Chryseobacterium sp. PL6.4 was coagulated the buffalo milk. However, the total coagulation of buffalo milk was observed with the strain P. putida PL4.2. It was observed that crude enzymes of P. putida PL4.2 and P. fluorescens PL5.4 showed initial coagulation, whereas P. aeruginosa PL3.4, P. fluorescens PL5.4 and Chryseobacterium sp. PL6.4 showed partial coagulation of buffalo milk. As a negative control, the milk was not coagulated. The positive control showed coagulation (Table 2).
Characterization and stability of the crude enzyme of P. fluorescens PL5.4: The maximum activity was observed at 40°C, which is considered the optimal temperature (Fig. 1a). This isolate still showed enzymatic activity at 80°C. The crude enzyme of strain PL5.4 maintained at least 65% of maximum activity within the pH interval between 4.0 and 10.0. Maximum activity was observed at pH 7.0, with 0.2 M phosphate buffer (Fig. 1b). The PMSF at 5 mM caused about 5% inhibition, indicating that the activity of the crude enzyme from isolate PL5.4 could not be an associated with a serine protease. The metalloprotease inhibitor EDTA resulted in partial inhibition, reaching about 35% at 5 mM (Table 3). Organic solvents were tested and the results are shown in Fig. 1c. The crude enzyme remained stable under the conditions tested, no presenting statistical difference. The enzymatic activity was affected by certain surfactants such as Triton X-100, Tween 80 and SDS.
| Table 2: | Clotting bovine milk and buffalo milk at 7 and 30°C by crude enzymes of psychrotrophic bacteria isolated from refrigerated raw buffalo milk |
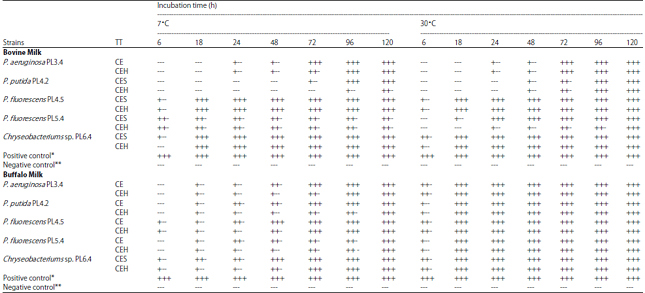 | |
TT, Treatment, CE: Crude enzyme, CEH: Heat-treated crude enzyme; —: No coagulation; +–: Initiation of coagulation; ++-: Partial coagulation; +++: Total coagulation, *Positive control: only milk (2 mL) with 110 μL of 1% sodium azide, **Negative control: milk (2 mL) with commercial chymosin enzyme (1 mL) | |
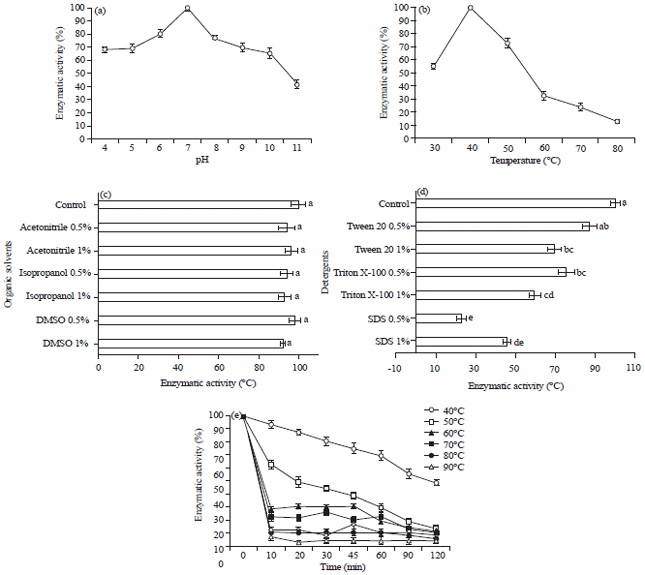 | |
| Fig. 1(a-e): | Effect on incubation different conditions on the proteolytic activity of P. fluorescens PL5.4: (a) Effect of pH, (b) Effect of incubation temperature, (c) Effect of organic solvents, (d) Effect of detergents and (e) Thermal stability after pre-incubation of 10-120 min. Tests using azocasein as substrate, Results are expressed as % of enzyme activity. Values are Means±Standard Deviations of 3 independent determinations. The letters in superscript represent statistical differences: different letters represent a statistically significant difference and the same letters do not present statistical difference |
When SDS was used at concentrations of 0.5 and 1% residual activity was 22.8 and 45.5 %, respectively (Fig. 1d), statistically different when the other detergents were applied. The effect of different salts was also evaluated, resulting in a reduction of about 20-30% in the enzyme activity (Table 4). However, when the enzyme was treated with CuCl2 the inhibition reached 80%. For incubation at temperatures higher than 50oC, a rapid decrease in enzyme activity was observed within the first 10 min incubation (Fig. 1e). It was observed that less than 30% of residual activity was maintained with incubations above 60°C for 10 min. Incubation at 40°C for 10 and 120 min resulted in a residual activity of 94 and 49%, respectively.
| Table 3: | Effect of protease inhibitors on the crude enzyme of P. fluorescens PL5.4, in 0.2 M phosphate buffer pH 7.0, at 40°C using azocasein as substrate |
 | |
PMSF: Phenylmethylsulfonyl fluoride, EDTA: Ethylenediaminetetracetic acid disodium salt. Results are expressed as % inhibition of enzymatic activity. Values are Means±Standard Deviations of three independent determinations | |
At 50°C residual activity of 63 and 13% was observed when time periods of 10 and 120 min were tested, respectively.
| Table 4: | Effect of salts of the crude enzyme of P. fluorescens PL5.4, in 0.2 M phosphate buffer pH 7, at 40°C using azocasein as substrate |
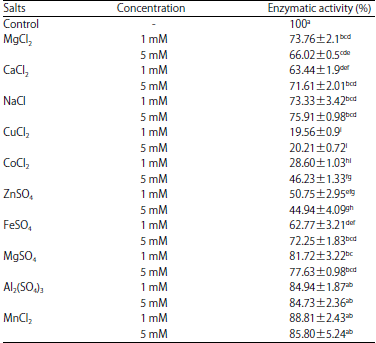 | |
Results are expressed as % of enzyme activity. Values are Means±Standard Deviations of 3 independent determinations. The letters in superscript represent statistical differences: different letters represent a statistically significant difference and the same letters do not present statistical difference. ANOVA test with a critical probability of p<0.05 followed by the Tukey’s test | |
DISCUSSION
In this study, the proteolytic profile displayed by psychrotrophic bacteria in buffalo milk agar demonstrates the potential for hydrolysis of the protein constituent of milk. Among the psychrotrophic bacteria, those from the Pseudomonas genus are the main cause of deterioration of milk and dairy products and are the microorganisms most frequently isolated from raw material and the finished product12. As observed in this study, in accordance with other authors, the presence of Pseudomonas is likely to cause deterioration of raw milk under refrigerated conditions9. Baur et al.7 confirmed a high prevalence (80%) of enzymatically active Gram-negative isolates, particularly various species of Pseudomonas, including P. aeruginosa and P. fluorescens.
In this study, all isolates showed proteolytic activity when tested on azocasein as a substrate, confirming the deteriorating potential of these psychrotrophic bacteria. Furthermore, it was observed that the maximal enzyme activity was demonstrated by P. aeruginosa PL3.4 at pH 5.5. Considering that this psychrotrophic strain showed elevated proteolytic activity in diverse pH, including the typical pH of both buffalo milk (6.4-6.9)18 and bovine milk (6.6-6.8)18, it seems even more important to consider the implications of high quantities of these bacteria in both types of milk.
A relevant aspect of proteolytic enzymes is that they were not eliminated by the conventional thermal treatment of dairy. Thus, once produced by psychrotrophic bacteria in milk, this raw material remains after treatments such as pasteurization7. Five (23.8%) isolates of psychrotrophic bacteria tested in this study were producers of heat-resistant proteolytic enzymes. The clotting ability of heat-treated crude enzymes in bovine or buffalo milk was evaluated, suggesting the risk that these bacteria and their enzymes represent, since their presence in milk and dairy products may lead to sensory and economic losses. Similar results were reported by Teh et al.19, who found that among 153 enzyme-producing isolates from milk tankers, 52 (33.98%) produced heat-stable enzymes.
When the clotting test was performed in milk, it was possible to visualize the effect of the proteolytic enzymes produced by the isolates as well as their resistance to thermal treatment (63.5°C for 30 min). In raw milk, k-casein and β-casein are the protein components most susceptible to proteolysis by psychrotrophic microorganisms20. Once the thermal resistant proteases are present in the milk, they attack milk proteins, thereby affecting the quality of dairy products as proteases degrade casein micelles and release soluble components such as polypeptides and amino acids. These components are lost in the whey and thereby reduce the yield of dairy products and change the standards of identity and quality of food8.
Detection and control of protease activity prior to processing are critical because the heat-resistant and able enzymes remain active in foods under different pH values and temperatures21. In addition, knowing the heat resistance characteristics, pH values and other variables is important for understanding the behavior of enzymes potentially present in milk samples. Thus, a partial characterization of the enzymatic activity of the P. fluorescens PL5.4 was performed.
The crude enzyme of P. fluorescens PL5.4 was active at the highest temperature tested (80°C) with 12.9% residual activity, meaning that in a process of rapid pasteurization where the milk is processed at 72-75°C for 15-20 sec, the enzyme remains active. However, the maximum enzyme activity was observed at 40°C, which is similar to other studies where the proteolytic activity of Pseudomonas isolates has been observed at 40-45°C16,22.
When the influence of the time/temperature binomial on the crude enzyme of P. fluorescens PL5.4 was evaluated, residual activity of 26.5% was noted at 70°C/30 min and 12.9% at 80°C/10 min treatment. Thus, considering the thermal pasteurization treatments used in the dairy industry (pasteurization at 62-65°C/30 min and rapid pasteurization 72-75°C/15-20 sec), the supernatant would survive these processes, thus representing a possible cause of deterioration in dairy products.
The evaluation of different pH conditions demonstrates the maintenance of enzymatic activity at pH value below 7.0. Whereas, the pH of the milk is between 6 and 7, authors observed that the residual activities were about 80% at pH 6.0 and 100% at pH 7.0. Similar results were found in the studies of Zambare et al.23 and Raj et al.22 where the greatest enzymatic activity of P. aeruginosa MCM B-327 was also measured at a pH value of 7. The crude enzyme of P. fluorescens PL5.4 exhibits enzymatic activity over a wide pH range (pH 4-10) with an optimum pH value of 7.0 and an optimum temperature of 40°C. Zhang and Lv21 found similar results, with activity in a pH range of 5-10 and an optimum pH value of 7.0 for the proteolytic activity of P. fluorescens BJ-10.
Some metal ions are important for enzyme activity and their structural properties23. In this study, CuCl2, ZnSO4 and CoCl2 had an inhibitory effect on enzymatic activity P. fluorescens PL5.4. Zhang and Lv21 found that ions Cu2+ had a negative effect on the enzymatic activity of P. fluorescens BJ-10. Other studies show that proteolytic activity can be increased with the ions Ca2+, Mg2+ and Zn2+ 24. This effect of Ca2+ and Mg2+ has been associated with the increase in thermal stability of diverse proteases25. However, these ions caused about 20% reduction in the activity of P. fluorescens PL5.4 enzyme. In addition, the protease activity survives various detergent and organic solvent tests. Similarly, in the study Siti et al.26 the addition of organic solvents showed no effect on the protease of P. aeruginosa. Finally, with the inhibitors used in this study, it was not possible to acquire precise information about the nature of the proteolytic activity studied. Although the crude enzyme of P. fluorescens PL5.4 was partially inhibited by EDTA, suggesting that a metalloprotease is involved, Pseudomonas spp. are recognized to produce multiple proteases27. Thus, further studies are necessary with more purified enzyme preparation to warrant a deeper understanding of the enzyme properties.
CONCLUSION
The results of this study show that psychrotrophic bacteria isolated from buffalo milk have the ability to produce extracellular proteolytic enzymes. The enzymes studied showed thermal resistance at a slow pasteurization temperature (63.5°C for 30 min). The characterization of the crude enzyme of P. fluorescens PL5.4 showed an ideal enzymatic activity at pH 7, an ideal temperature of 40°C, resistant to the detergents and solvents tested and sensitive to some salts. The milk coagulation observed by the effect of the enzymes produced shows the potential for degradation of milk protein. The concern for the presence of psychrotrophic bacteria producing proteolytic enzymes, is their negative effect on the quality of buffalo milk. This work seeks to collaborate with producers and dairy farmers to implement good practices in obtaining milk, ensuring its microbiological quality.
SIGNIFICANCE STATEMENT
This study investigates the proteases produced by psychotrophic bacteria isolated from buffalo milk. This study will help the researchers to declare the need to adopt good practices for obtaining buffalo milk. Therefore, the presence of high amounts of psychrotrophic bacteria can interfere with the constituents in the milk, causing a negative impact on the production of the products derived from buffalo milk.. Thus, the implementation of care for the maintenance of the quality of buffalo milk, will have a positive impact on the production chain as a whole.
REFERENCES
- Han, X., F.L. Lee, L. Zhang and M.R. Guo, 2012. Chemical composition of water buffalo milk and its low-fat symbiotic yogurt development. Funct. Foods Health Dis., 2: 86-106.
Direct Link - Hussain, I., J. Yan, A.S. Grandison and A.E. Bell, 2012. Effects of gelation temperature on Mozzarella-type curd made from buffalo and cows' milk: 2. Curd yield, overall quality and casein fractions. Food Chem., 135: 1404-1410.
CrossRefDirect Link - Quigley, L., O. O'Sullivan, C. Stanton, T.P. Beresford, R.P. Ross, G.F. Fitzgerald and P.D. Cotter, 2013. The complex microbiota of raw milk. FEMS Microbiol. Rev., 37: 664-698.
CrossRefDirect Link - Vacheyrou, M., A.C. Normand, P. Guyot, C. Cassagne, R. Piarroux and Y. Bouton, 2011. Cultivable microbial communities in raw cow milk and potential transfers from stables of sixteen French farms. Int. J. Food Microbiol., 146: 253-262.
CrossRefDirect Link - Sitohy, M., S. Mahgoub and A. Osman, 2011. Controlling psychrotrophic bacteria in raw buffalo milk preserved at 4°C with esterified legume proteins. LWT-Food Sci. Technol., 44: 1697-1702.
CrossRefDirect Link - Raats, D., M. Offek, D. Minz and M. Halpern, 2011. Molecular analysis of bacterial communities in raw cow milk and the impact of refrigeration on its structure and dynamics. Food Microbiol., 28: 465-471.
CrossRefDirect Link - Baur, C., M. Krewinkel, B. Kranz, M. von Neubeck and M. Wenning et al., 2015. Quantification of the proteolytic and lipolytic activity of microorganisms isolated from raw milk. Int. Dairy J., 49: 23-29.
CrossRefDirect Link - Button, P.D., H. Roginski, H.C. Deeth and H.M. Craven, 2011. Improved shelf life estimation of UHT milk by prediction of proteolysis. J. Food Qual., 34: 229-235.
CrossRefDirect Link - Bagliniere, F., A. Mateos, G. Tanguy, J. Jardin and V. Briard-Bion et al., 2013. Proteolysis of ultra high temperature-treated casein micelles by AprX enzyme from Pseudomonas fluorescens F induces their destabilisation. Int. Dairy J., 31: 55-61.
CrossRefDirect Link - Marchand, S., J. de Block, V. de Jonghe, A. Coorevits, M. Heyndrickx and L. Herman, 2012. Biofilm formation in milk production and processing environments; Influence on milk quality and safety. Compr. Rev. Food Sci. Food Saf., 11: 133-147.
CrossRefDirect Link - Bekker, A., P. Jooste, L. Steyn, C. Bothma, A. Hugo and C. Hugo, 2016. Lipid breakdown and sensory analysis of milk inoculated with Chryseobacterium joostei or Pseudomonas fluorescens. Int. Dairy J., 52: 101-106.
CrossRefDirect Link - Ruaro, A., C. Andrighetto, S. Torriani and A. Lombardi, 2013. Biodiversity and characterization of indigenous coagulase-negative staphylococci isolated from raw milk and cheese of North Italy. Food Microbiol., 34: 106-111.
CrossRefDirect Link - Daroit, D.J., A.P.F. Correa and A. Brandelli, 2009. Keratinolytic potential of a novel Bacillus sp. P45 isolated from the Amazon basin fish Piaractus mesopotamicus. Int. Biodeterior. Biodegrad., 65: 358-363.
CrossRefDirect Link - Nornberg, M.F.B.L., M.L. Mentges, S.T. Silveira, E.C. Tondo and A. Brandelli, 2011. A psychrotrophic Burkholderia cepacia strain isolated from refrigerated raw milk showing proteolytic activity and adhesion to stainless steel. J. Dairy Res., 78: 257-262.
CrossRefDirect Link - Nornberg, M.F.B.L., R.S. Friedrich, R.D. Weiss, E.C. Tondo and A. Brandelli, 2010. Proteolytic activity among psychrotrophic bacteria isolated from refrigerated raw milk. Int. J. Dairy Technol., 63: 41-46.
CrossRefDirect Link - Mateos, A., M. Guyard-Nicodeme, F. Bagliniere, J. Jardin and F. Gaucheron et al., 2015. Proteolysis of milk proteins by AprX, an extracellular protease identified in Pseudomonas LBSA1 isolated from bulk raw milk, and implications for the stability of UHT milk. Int. Dairy J., 49: 78-88.
CrossRefDirect Link - Teh, K.H., S. Flint, J. Palmer, P. Andrewes, P. Bremer and D. Lindsay, 2012. Proteolysis produced within biofilms of bacterial isolates from raw milk tankers. Int. J. Food Microbiol., 157: 28-34.
CrossRefDirect Link - Menard, O., S. Ahmad, F. Rousseau, V. Briard-Bion, F. Gaucheron and C. Lopez, 2010. Buffalo vs. cow milk fat globules: Size distribution, zeta-potential, compositions in total fatty acids and in polar lipids from the milk fat globule membrane. Food Chem., 120: 544-551.
CrossRefDirect Link - Teh, K.H., S. Flint, J. Palmer, D. Lindsay, P. Andrewes and P. Bremer, 2011. Thermo-resistant enzyme-producing bacteria isolated from the internal surfaces of raw milk tankers. Int. Dairy J., 21: 742-747.
CrossRefDirect Link - Baglinière, F., J. Jardin, F. Gaucheron, A.F. de Carvalho and M.C.D. Vanetti, 2017. Proteolysis of casein micelles by heat-stable protease secreted by Serratia liquefaciens leads to the destabilisation of UHT milk during its storage. Int. Dairy J., 68: 38-45.
CrossRefDirect Link - Zhang, S. and J. Lv, 2014. Purification and properties of heat-stable extracellular protease from Pseudomonads fluorescens BJ-10. J. Food Sci. Technol., 51: 1185-1190.
CrossRefDirect Link - Raj, A., N. Khess, N. Pujari, S. Bhattacharya, A. Das and S.S. Rajan, 2012. Enhancement of protease production by Pseudomonas aeruginosa isolated from dairy effluent sludge and determination of its fibrinolytic potential. Asian Pac. J. Trop. Biomed., 2: S1845-S1851.
CrossRefDirect Link - Zambare, V., S. Nilegaonkar and P. Kanekar, 2011. A novel extracellular protease from Pseudomonas aeruginosa MCM B-327: Enzyme production and its partial characterization. New Biotechnol., 28: 173-181.
CrossRefPubMedDirect Link - Mu, Z., M. Du and Y. Bai, 2009. Purification and properties of a heat-stable enzyme of Pseudomonas fluorescens Rm12 from raw milk. Eur. Food Res. Technol., 228: 725-734.
CrossRefDirect Link - Daroit, D.J., V. Sant’Anna and A. Brandelli, 2011. Kinetic stability modelling of keratinolytic protease P45: Influence of temperature and metal ions. Applied Biochem. Biotechnol., 165: 1740-1753.
CrossRefDirect Link - Siti, S.D. and R. Hertadi, 2015. Isolation and characterization of organic-solvent stable protease isolated by Pseudomonas stutzeri BK AB-12. Procedia Chem., 16: 341-348.
CrossRefDirect Link - Alves, M.P., R.L. Salgado, M.R. Eller, P.M.P. Vidigal and A.F. de Carvalho, 2016. Characterization of a heat-resistant extracellular protease from Pseudomonas fluorescens 07A shows that low temperature treatments are more effective in deactivating its proteolytic activity. J. Dairy Sci., 99: 7842-7851.
CrossRefDirect Link