
Trini Susmiati
Department of Biochemistry, Faculty of Veterinary Medicine, Gadjah Mada University, 55281 Yogyakarta, Indonesia
LiveDNA: 62.16928
Prabowo Purwono Putro
Department of Reproduction and Obstetrics, Faculty of Veterinary Medicine, Gadjah Mada University, 55281 Yogyakarta, Indonesia
Sarmin
Department of Physiology, Faculty of Veterinary Medicine, Gadjah Mada University, 55281 Yogyakarta, Indonesia
LiveDNA: 62.15329
Claude Mona Airin
Department of Physiology, Faculty of Veterinary Medicine, Gadjah Mada University, 55281 Yogyakarta, Indonesia
Hartiningsih
Department of Surgery and Radiology, Faculty of Veterinary Medicine, Gadjah Mada University, 55281 Yogyakarta, Indonesia
Pamungkas Bagus Satria
Faculty of Veterinary Medicine, Gadjah Mada University, 55281 Yogyakarta, Indonesia
International Journal of Dairy Science
Year: 2017 | Volume: 12 | Issue: 4 | Page No.: 282-288







ABSTRACT
Background and Objective: Polymorphisms of kappa-casein protein genes are closely related to the quality of milk production and composition. The purpose of the study was to investigate polymorphisms in genes encoding kappa-casein (κ-casein) milk protein produced by the dairy cattle of KP4/PIAT UGM Yogyakarta and BBPTU-HPT Baturraden. Materials and Methods: Twenty-three cattle were used to investigate kappa-casein polymorphisms in dairy cows from two different animal farms. Nine cattle were distributed from BBPTU-HPT Baturraden, Central Java and 14 cattle were allocated from KP4/PIAT UGM Yogyakarta. The research procedures included blood sampling, DNA extractions and DNA amplification using Polymerase Chain Reaction (PCR). Kappa-casein gene PCR products were analyzed by using the PCR-RFLP technique with restriction enzymes Hinf I and Hind III. Results: The results of restriction digestion using the Hinf I showed four restriction fragments in the blood sample from BPPTU-HPT Baturraden and three restriction fragments from KP4/PIAT UGM Yogyakarta. The digestion of PCR product using Hind III produced one restriction fragment in the blood samples from KP4/PIAT UGM Yogyakarta and 4 fragments in the samples from BPPTU-HPT Baturraden. Conclusion: The dairy cattle of BPPTU Baturraden offer a more favorable genetic selection of animals for milk production and industry due to their genetic variants in terms of genotypes and alleles.
PDF Abstract XML References Citation
Received: October 06, 2016;
Accepted: April 28, 2017;
Published: June 15, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Trini Susmiati, Prabowo Purwono Putro, Sarmin, Claude Mona Airin, Hartiningsih and Pamungkas Bagus Satria, 2017. Polymorphisms in Genes Encoding Kappa Casein Milk Protein in Dairy Cattle. International Journal of Dairy Science, 12: 282-288.
DOI: 10.3923/ijds.2017.282.288
URL: https://scialert.net/abstract/?doi=ijds.2017.282.288
DOI: 10.3923/ijds.2017.282.288
URL: https://scialert.net/abstract/?doi=ijds.2017.282.288
INTRODUCTION
Indonesia has many imported dairy cattle that have potential in developing and improving the productivity of national dairy farms. There are some groups of dairy cattle that have such specific characteristics as to be productive in milk production and potential to inherit good cattle heredity to the next generation (offspring). The existence and the potential of dairy cattle that have been experiencing domestication can support the economy in the rural areas in Indonesia1.
There are numerous factors that affect the productivity of dairy cattle. The environmental factors are the main causes affecting the quantity and quality of milk production. In addition, genetic variations also contribute to the milk production and composition and reproduction of dairy cattle. Characterizing the frequency of the genes that encode milk proteins, especially casein, is a basic method to determine the change rate of livestock selection value2. The variation of the milk protein genes in dairy cattle basically determines the development of the national dairy farm industry. For example, in Indonesia, fresh milk is generally produced from dairy cattle imported from Australia or New Zealand, such as the Holstein-Friesian breeds. The improvement of milk production traits can be done through the identification of the diversity of the DNA fragments of kappa-casein genes3.
There are four types of casein protein molecule, alpha-S1-casein (αS1-casein), alpha-S2-casein (αS2-casein), beta-casein (β-casein) and κ-casein (κ-casein)4-6. All four types of casein are characterized by specific properties such as low solubility at pH 4.6 and organization into a chained group of proteins called micelles7. Furthermore, three of the four caseins (αS1-casein, αS2-casein and β-casein) have a high level of sensitivity to the precipitation of calcium. Whey protein constitutes 20% of total milk protein and consists of α-lactalbumin, which is involved in lactose synthesis and β-lactoglobulin, which may be a carrier of hydrophobic molecules. In contrast to calcium-sensitive αS1-casein, αS2-casein and β-casein; κ-casein plays an important role in the formation, stabilization and aggregation of the casein micelles and thereby influences the technological and nutritional properties of milk8. In the last few decades, many studies on κ-casein protein genes have been conducted to obtain the κ-casein genotype variations. Therefore, the relationship between gene variations and milk protein production has been generally known. However, the gene variations that encode the milk protein genes in Indonesian dairy cows are still less frequently observed. These points provide an opportunity for the authors to study polymorphism in kappa casein genes as the marker assisted assistance strategy developed to increase the productivity and improve the quality of milk4.
The objective of the present study was to investigate the polymorphisms in kappa-casein genes of dairy cattle as an effort to identify more genotype variants of kappa casein useful for the improvement of the quantity and quality of dairy products in Indonesia.
MATERIALS AND METHODS
Materials of the study: The laboratory tools used in this research included a 0.5-10 μL micropipette (Nichiryo Nichipet EX), 10-100 μL and 100-1000 μL micropipettes (Thermo Scientific Finn pipette), 10, 100 and 1000 μL pipette tips produced by SBS Beijing Genentech Co. Ltd, a water bath (Eyela, Uni Thermo Shaker NTS 3000), a centrifuge (5804R), a freezer; a PCR machine (Infigen, Biotech, Inc.), electrophoretic devices, a microwave (Sanyo model EM-ST68), one set of gel casters (with plate and comb), a scale, a magnetic stirrer, a vortex mixer, a spin down, a measuring cup, gloves, Erlenmeyer tubes and a UV transilluminator.
The materials used for this study included dairy cattle blood from KP4/PIAT UGM Yogyakarta, Gadjah Mada University teaching farm and BBPTU-HPT Baturraden (a breeding farm in Purwokerto, Central Java, enzymes Hinf I and Hind III, Tris-HCl, EDTA, absolute ethanol, PCR reaction buffer, dNTP mix, MgCl2, a Genomic DNA Mini Kit (Cat. No. GB 100/Lot No. JM02202), two primers [(forward and reverse: K1 (5’-CAC GTC ACC CAC ACC CAC ATT TAT-3’) and K2 (5’-TAA TTA GCC CAT TTCGCC TTC TCT-3’)], Bioatlas (0.5 μg mL–1), BRL as a marker, TBE (Tris-boric acid-EDTA) buffer, TE buffer and agarose (SBS, Beijing, Genentech, Co. Ltd).
DNA isolation: The DNA was isolated and purified from dairy cattle blood using the DNA isolation kit. The blood samples were stored at 4°C. The DNA quality was analyzed by its displacement on 1% agarose gel using 1X TBE buffer (89 mM Tris, 89 mM boric acid and 2 mM EDTA, pH 8.0) in submarine electrophoresis devices (Hoefer, USA) with gel dyed using Bioatlas (0.5 μg mL–1), which was followed by observation under UV light (λ = 300 nm).
DNA electrophoresis: Electrophoresis was performed in agarose to see the results of the DNA isolation and PCR. The first step was to make the gel. The concentration of the gel used to examine the PCR results was 1%, consisting of 0.25 g of agarose dissolved in 25 mL Aquades water. The solution was then heated using a microwave until it boiled.
Next, 2 μL Bioatlas was added to the liquid gel. The agarose solution was allowed to stand until it reached approximately 55°C and then it was poured into a caster with a comb.
After the gel hardened, the plate filled with the agarose gel was put into an electrophoresis tank filled with the same buffer solution used to make the agarose gel, using 0.5X TBE buffer. Then, a 3 μL sample of the DNA isolation and PCR result was taken by micropipette and mixed with 1 μL of Glycerin Bromophenol Blue (GBB) and placed into the agarose gel well. A 1 kb DNA marker was also included in one of the wells. Electrophoresis was performed at 100 V and the process was completed in 20-30 min The molecular bands were then observed with the help of a UV transilluminator (λ = 260 nm).
Amplification of DNA fragments: The oligonucleotide primers were designed specifically for the gene (in the DNA genome) with some modifications. The 50 μL PCR reaction mixture consisted of 2.5 mM MgCl2, 10 mM dNTPs, 100-300 ng DNA template, 10 pmol of each primer and 2 U Taq polymerase with buffer. The DNA amplification was performed using a PCR machine. The gene for milk casein protein was amplified under the following conditions: initial denaturation for 5 min at 96°C followed by primer attachment (annealing) at 72°C for 50 sec for elongation, the amplification process was performed in 335 cycles, ending with 10 min at 72°C. The PCR results were detected by the displacement on a 1% agarose gel using the 1X TBE buffer on a Submarine Electrophoresis device (Hoefer, USA). Observation of the gel was performed with the help of UV light after staining the gel with ethidium bromide. A 100 bp DNA marker was used to estimate the molecular weight.
PCR-product digestion: The PCR product was cut using the Hinf I and Hind III enzymes as follows: 15 μL of each PCR product was digested separately with 7.5 U of each restriction enzyme (Hinf I and Hind III) in a 20 mL final reaction volume containing 1X enzyme buffer, which was incubated at 37°C for 3 h in a water bath. After incubation, the products were subjected to electrophoresis on a 2% agarose gel using 1X TBE buffer containing 0.5 mg mL–1 DNA at 50 V until the bands were fully separated. Each DNA molecule was near 100 bp in size, which was thus used as the marker size. The bands, which were visualized under UV light, revealed polymorphic loci based on the sizes of the DNA fragments.
Statistical analysis: The genotype frequency of kappa casein genes in the present study was calculated with a direct counting9. The data of this study was presented descriptively in the form of genotype distribution10.
RESULTS
Genomic DNA extraction: The results of the isolation of the genomic DNA molecule are shown in Fig. 1. The DNA profile of the genome shows that DNA extraction of the red blood samples from both KP4/PIAT UGM Yogyakarta and BBPTU-HPT Baturraden had been successfully produced.
 | |
| Fig. 1(a-b): | Representative DNA extraction of blood samples from (a) BPPTU-HPT Baturraden and (b) KP4 UGM Yogyakarta analyzed on 1% (polyacrylamide) agarose electrophoretic gel |
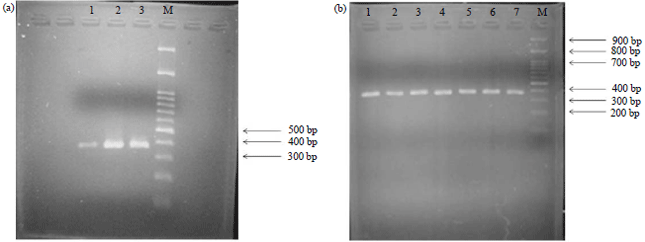 | |
| Fig. 2(a-b): | Representative DNA fragment amplification of blood samples from (a) BPPTU-HPT Baturraden, Central Java and (b) KP4/PIAT UGM, Yogyakarta analyzed on 1% (polyacrylamide) agarose electrophoretic gel with markers (a) 1, 2 and 3 and (b) 1, 2, 3, 4, 5, 6 and 7 |
| Table 1: | Sequence and number of oligonucleotide primers and melting temperature for each primer used in DNA amplification |
The DNA genomes were obtained with a molecular weight greater than 1000 bp and were then amplified with the DNA genomes used as a template for Polymerase Chain Reaction (PCR) with K1 as forward primer and K2 as the reverse primer . The primer base sequences and melting temperatures are presented in Table 1.
Amplification of DNA: In the amplification of DNA, the primary sequences used were K1 (5’-CAC GTC ACC CAC ACC CAC ATT TATC 3’) and K2 (5’-TAA TTA GCC CAT TTC GCC TTC TCT GT 3’). The electrophoretic result of kappa casein genes through PCR was then detected with a 1% agarose gel as presented in Fig. 2.
Digestion of PCR product: In this study, PCR-RFLP techniques were utilized to investigate polymorphisms in kappa-casein genes on the cows and to differentiate the genotypes of κ-casein alleles, especially DNA band patterns which can be clearly distinguished from the results of the selected restriction enzymes11. The PCR-RFLP procedure uses restriction enzymes to cut certain nucleotide sequences and performs PCR cycles on one gene. The restriction enzymes used in this study were Hinf I and Hind III using a 2% agarose electrophoretic gel. The result of the amplification of 379 bp PCR product with the restriction enzyme Hind III showed that 22 samples from KP4/PIAT UGM Yogyakarta and BBPTU-HPT Baturraden were the AA genotype with the restriction fragment of 379 bp because the PCR product remained undigested. Three restriction fragments of 379, 220 and 170 bp digested in one sample of BBPTU-HPT Baturraden, identified as AB genotype. Meanwhile, the result of the PCR-RLFP with the restriction enzyme Hinf I showed that there were three restriction fragments of 170, 140 and 90 bp identified as AA genotype and four restriction fragments of 270, 170, 140 and 90 identified as AB genotype (Fig. 3).
DISCUSSION
The current study demonstrates that the polymorphism has occurred in the DNA extracted blood samples from dairy cattle in KP4/PIAT UGM Yogyakarta and BBPTU-HPT Baturraden. The digestion of 379 bp amplified fragment of kappa-casein genes with Hinf I and Hind III generated two restriction patterns of AA (170, 140 and 90 ) and AB (270, 220, 170 and 90 bp) and AA (379 bp) and AB (379, 220 and 170 bp), respectively. All the blood samples of KP4 UGM Yogyakarta were homozygous with the identified genotype AA (100%). Therefore, the samples were classified into monomorphic kappa-casein genotypes. On the contrary, the samples of BPPTU Baturraden were heterozygous with two identified patterns of genotypes AA (89%) and AB (11%).
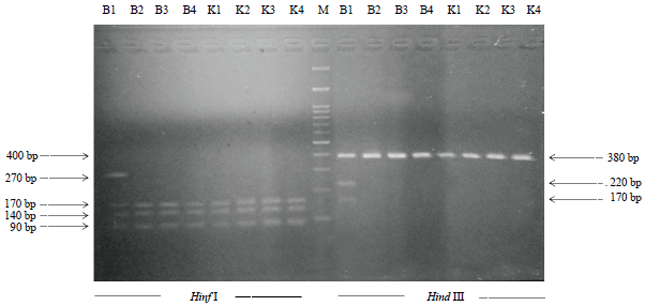 | |
| Fig. 3: | Electrophoretic patterns of 379 bp PCR products of Kappa casein gene digested with Hinf I and Hind III endonuclease on 2% (polyacrylamide) agarose electrophoretic gel |
Hence, these samples were identified as polymorphic genotypes. Overall the present study also showed that the genotype frequencies are dominated by AA genotype (96%), followed by AB genotype (4%) and BB genotype (0%). The genotype findings of this study are in agreement with the findings by Azevedo at al.12. They reported that frequencies of genotype AA, AB and BB in Gyr cows were respectively 97, 3 and 0%.
Meanwhile, Botaro et al.13 revealed AA genotype (68.83%), AB genotype (31.84%), and BB genotype (1.33%) in Holstein dairy cattle and that AA genotype (71.38%), AB genotype (27.90%) and BB genotype (0.72%) in Girolando dairy cattle. Trakovicka et al.14 reported three genotypes AA (58%), AB (36%) and BB (6%). The genetic characterization of Holstein-Friesian dairy cattle in KP4 UGM Yogyakarta and BBPTU-HPT Baturraden shows the A allele frequency (0.94) of kappa casein is higher than the B allele frequency (0.06). The present study of are in accordance with the previous study presented by Dedovic et al.15 who reported that the frequencies of A and B alleles in examined breeds were 0.667 and 0.333 for the Simmental breed, 0.875 and 0.125 for cross breeds and 0.667 and 0.333 for Busha cattle, respectively. Cortes-Lopez et al.16 reported the similar research findings in double-purpose cattle. They revealed that the allele frequency of A and B were 0.67 and 0.33. From the comparison of allele frequency with the other studies, the A allele frequency in Holstein-Friesian dairy cattle of KP4/PIAT UGM Yogyakarta and BBPTU-HPT Baturraden is the highest. This finding is in accordance with the research findings reported by Azevedo et al.12 They reported that the A allele was the most frequent, with values ranging from 0.7 to 0.99 for 1316 animals from the Gyr, F1 Gyr-Holstein crossbreeds, Guzerat, Nellore and Sindhi breeds12. Trakovicka et al.14 reported that the A allele (0.7563) was more dominant than the B allele (0.2437) in Simmental-Holstein cross breeds. The current study conclusively shows that the A allele of kappa casein is dominant among all alleles. The AA genotype (A allele) dominance in Indonesian dairy cattle was caused by the artificial insemination applied to the dairy cattle crossbreeding programs conducted by the Indonesian government. Most of the active male cattle used in the artificial insemination were of AA genotype17. This identified kappa casein genes, then, can be employed as a genetic marker which positively affects the milk protein and milk production18 . Sitkowska et al.19 reported that higher milk production was significantly correlated to the AA genotypes in Holstein cows. Sulimova et al.6 discovered that the cows with BB genotype produced higher cheese yield than the cows with AA genotype. Alipanah et al.20 reported that Red-pied cows with AA genotype produced higher milk yield. Sitkowska et al.19 demonstrated that the AA genotype showed higher milk, fat and protein yield than others genotypes. Tracovicka et al.14 concluded that AA genotype (A allele) had positive effects on milk yields and B allele on protein and fat yield. Meanwhile, the B allele has a better thermal resistance and shorter coagulation time, better curdles and contains micelles of different size, which is preferable for cheese-making. The milk of cows with genotype BB produced higher cheese yield than the milk of AA genotype6. From the A allele dominance over the (minority) B allele, the dairy cattle of BPPTU-HPT Baturraden and KP4 UGM are more suitable for cow milk yield production but not suitable for cheese-making industry. From the polymorphisms in kappa-casein genes in the dairy cattle, the cattle from BPPTU Baturraden show more genetic variants in genotypes and alleles (polymorphism) with the complete milk protein production traits, which can be a good genetic marker for the next animal selection.
CONCLUSION
In the present study, the digestion of PCR product using Hinf I and Hind III produced two patterns of genotypes AA and AB. The genotype frequency of kappa-casein genes in the two dairy cattle breeding farms showed that the dairy cattle have genotype AA (96%), AB (4%) and BB (0%). The Samples of KP4 UGM Yogyakarta were monomorphic and the samples of BPPTU Baturraden were polymorphic. The dairy cattle from BPPTU Baturraden offer a more favorable genetic selection of animals for milk production and industry due to their genetic variants in terms of genotypes and alleles.
SIGNIFICANT STATEMENTS
The polymorphism in kappa-casein protein genes has occurred in Holstein-Frisian dairy cattle from the two dairy farms in general. The blood samples from Yogyakarta (KP4) were identified as monomorphic kappa-casein genotypes, while the blood samples from Baturraden (BPPTU-HPT) were identified as polymorphic kappa-casein genotypes in particular. The dairy cattle from Baturraden offer more favorable genetic selection of animals for milk production and industry due to their genetic variants.
ACKNOWLEDGMENTS
We greatly thank to KP4/PIAT UGM Yogyakarta (Gadjah Mada University teaching farm) and BPPTU-HPT Baturraden (a city administration’s breeding farm) for their cooperation. The Directorate General of Higher Education, Ministry of Research and Education, the Republic of Indonesia is acknowledged for financial support (PUPT Grant, Contract number: 109/LPPM/2015, March 02, 2015).
REFERENCES
- Sutarno, S. and A.D. Setyawan, 2015. Review: Genetic diversity of local and exotic cattle and their crossbreeding impact on the quality of Indonesian cattle. Biodiversitas, 16: 327-354.
Direct Link - Anggraeni, A., 2013. Genetic improvement of milk quantity and milk quality in Holstein Friesian cattle by selection. Wartazoa: Indonesian Bull. Anim. Vet. Sci., 22: 1-11.
Direct Link - Rachagani, S. and I.D. Gupta, 2008. Bovine kappa-casein gene polymorphism and its association with milk production traits. Genet. Mol. Biol., 31: 893-897.
Direct Link - Riaz, M.N., N.A. Malik, F. Nasreen and J.A. Qureshi, 2008. Molecular marker assisted study of kappa-casein gene in Nili-Ravi (buffalo) breed of pakistan. Pak. Vet. J., 28: 103-106.
Direct Link - Sulimova, G.E., M. Ahani-Azari, J. Rostamzadeh, M.R. Mohammad Abadi and O.E. Lazebny, 2007. κ-casein gene (CSN3) allelic polymorphism in russian cattle breeds and its information value as a genetic marker. Russian J. Genet., 43: 73-79.
CrossRefDirect Link - Choobini, Z.M., M. Shadkhast, H. Moshtaghi, S.H. Dehkordi and H.R. Shahbazkia, 2014. Polymorphism of κ-casein gene in Iranian Holsteins. Iranian J. Biotechnol., 12: 56-60.
Direct Link - Byrne, G., 2007. A statistical primer: Understanding descriptive and inferential statistics. Evid. Library Inform. Pract., 2: 32-47.
CrossRefDirect Link - Stafuzza, N.B., K.J. Kochan, P.K. Riggs and M.E.J. Amaral, 2013. Single nucleotide variations in the buffalo kappa-casein gene (CSN3). Buffalo Bull., 32: 675-679.
Direct Link - Azevedo, A.L.S., C.S. Nascimento, R.S. Steinberg, M.R.S. Carvalho, M.G.C.D. Peixoto, R.L. Teodoro and M.A. Machado, 2008. Genetic polymorphism of the kappa-casein gene in Brazilian cattle. Genet. Mol. Res., 7: 623-630.
Direct Link - Botaro, B.G., Y.V.R. de Lima, C.S. Cortinhas, L.F.P. e Silva, F.P. Renno and M.V. dos Santos, 2009. Effect of the kappa-casein gene polymorphism, breed and seasonality on physicochemical characteristics, composition and stability of bovine milk. R. Bras. Zootec., 38: 2447-2454.
CrossRefDirect Link - Trakovicka, A., N. Moravcikova and A. Navratilova, 2012. Kappa-casein gene polymorphism (CSN3) and its effect on milk production traits. Acta Fytotechnica et Zootechnica, 15: 61-64.
Direct Link - Dedovic, R., V. Bogdanovic, P. Perisic, D. Stanojevic, J. Popovic and M. Brka, 2015. Relationship between genetic polymorphism of κ-casein and quantitative milk yield traits in cattle breeds and crossbreds in Serbia. Genetika, 47: 23-32.
CrossRefDirect Link - Cortes-Lopez, N.G., S. del Moral, J.A. Rueda, C. Luna-Palomera, C.A. Meza-Herrera and J. Abad-Zavaleta, 2012. Allelic and genotypic frequency of kappa casein gene in double purpose cattle. Trop. Subtrop. Agroecosyst., 15: 47-55.
Direct Link - Anggraini, A., C. Sumantri, A. Farajallah and E. Andreas, 2010. Kappa-casein genotypic frequencies in Holstein-Friesian dairy cattle in West Java province. Media Peternakan, 33: 61-67.
CrossRefDirect Link - Memon, I.A., M.A. Raza, W.A. Vistro, M.F. Leghari and A.W. Nizamani et al., 2016. Marker assisted molecular investigation of kappa-casein gene in Bos indicus Sindhi genetic group using HINFI restriction enzyme. Int. Arch. BioMed. Clin. Res., 2: 34-39.
CrossRefDirect Link - Alipanah, M., L. Kalashnikova and G. Rodionov, 2007. Association of prolactin gene variants with milk production traits in Russian Red Pied cattle. Iran. J. Biotech., 5: 158-161.
Direct Link