
Ibrahim A.A. Abou Ayana
Department of Dairy Research, Food Technology Research Institute (FTRI), Agriculture Research Center, Ministry of Agriculture, Giza, Egypt
LiveDNA: 20.14045
Amany M. El-Deeb
Department of Dairy Research, Food Technology Research Institute (FTRI), Agriculture Research Center, Ministry of Agriculture, Giza, Egypt
Amal Elsady Ibrahim
Department of Dairy Research, Food Technology Research Institute (FTRI), Agriculture Research Center, Ministry of Agriculture, Giza, Egypt
International Journal of Dairy Science
Year: 2016 | Volume: 11 | Issue: 3 | Page No.: 114-123







ABSTRACT
Moulds and yeasts are a major cause of fermented foods spoilage, such as kareish cheese with the low pH being selective for them and can produce off-flavors, sticky texture and cheese becomes softer. Seventy two samples of kareish cheese were collected, examined for incidence of moulds and yeasts. Positive samples were non-conforming, contained 4-5 log10 CFU g–1 moulds and yeasts. Ten species of moulds and nine species of yeasts were detected in collected samples. All propionibacteria strains showed antagonistic effect against moulds and yeasts isolates using agar well diffusion technique. Cheese made with Propionibacterium thoenii 119 and L. lactis subsp. lactis (LPT) contained the highest folate, vitamin B12, diacetyl and acetaldehyde followed by cheese made with Propionibacterium jensenii 118 and L. lactis subsp. lactis (LPJ) then made with Propionibacterium acidicpropionici 117 and L. lactis subsp. lactis (LPA). The LPT cheese achieved the greatest of yield and sensory evaluation. The populations of Aspergillus flavus and Candida albicans decreased by (1.41, 1.24 and 1.2) and (1.49, 1.28 and 1.1) log cycle in kareish cheese made with LPT, LPJ and LPA after 30 days, respectively. Experimental cheeses were higher adhesiveness, cohesiveness, gumminess, hardness and springiness than control and contained the lowest counts of moulds and yeasts.
PDF Abstract XML References Citation
Received: January 30, 2016;
Accepted: March 08, 2016;
Published: April 29, 2019
How to cite this article
Ibrahim A.A. Abou Ayana, Amany M. El-Deeb and Amal Elsady Ibrahim, 2016. Using of Dairy Propionibacteria as Bio-preservative in Kareish
Cheese. International Journal of Dairy Science, 11: 114-123.
DOI: 10.3923/ijds.2016.114.123
URL: https://scialert.net/abstract/?doi=ijds.2016.114.123
DOI: 10.3923/ijds.2016.114.123
URL: https://scialert.net/abstract/?doi=ijds.2016.114.123
INTRODUCTION
More than a quarter of sold dairy products are cheeses, kareish cheese is a soft cheese commonly made and consumed in Egypt. This cheese is an excellent source of protein, amino acids, calcium, phosphorus, vitamins and many micronutrients. It has low fat and salt, so doctors advise patients to eat it.
Nevertheless, high numbers of moulds and yeasts frequently can grow best on/in kareish cheese, producing off-flavors described as fermented, yeasty, gassy and moldy appearance and sticky texture as well as cheese becomes softer, so the shelf-life of this type of cheese is short. Their occurrence may be attributed to the mould and yeast’s ability to grow at low temperatures, the assimilation of organic acids like succinic, lactic and citric acid their proteolytic and lipolytic activities, resistance against high salt concentration, low αw and resistance to cleaning compounds and sanitizers (Alvarez-Martin et al., 2007). Elbagory et al. (2014) confirmed that 35 kareish cheese samples were positive for moulds and yeasts and they isolated A. fumigatus, A. terreus, Penicillium spp. and Fonsecaea spp., Candida albicans, Rhodotorula spp. and Candida famata. Moulds and yeasts were detected in kareish cheese in range 27-35×102 CFU g–1 (Metwalli, 2011). About 90% of kareish cheese samples were contaminated with mould and only Rhizopus spp. was identified (El-Kest et al., 2015). An average of yeasts 5.6 log10 was detected in kareish cheese they were S. cerevisiae, K. lactis and Candida sp. (El-Shafei et al., 2008). Many species of fungi were isolated from cheese types (Abou Ayana et al., 2014).
Chemical preservatives and artificial additives causing severe harmful to consumers, scientists make unremitting efforts to find out and selection of natural compounds for use as safe alternatives to chemical preservatives. Newly, using lactic acid bacteria and propionibacteria as bio-preservatives (Dabiza, 2006) to enhance food safety and stability as the antimicrobial systems possessed by these bacteria offer potential for effective natural preservation methods. MicrogardTM (Wesman Foods Inc. OR, USA) is a commercially available milk product fermented by P. freudenreichii subsp. shermanii and pasteurized after fermentation. The product promises effectiveness against most Gram-negative bacteria, some yeast and moulds (Daeschel, 1993). Bio profit contains viable cells of P. freudenreichii subsp. shermanii strain JS and co-culture of Lactobacillus rhamnosus and is effective for inhibiting yeasts growth in dairy products, Bacillus spp. in sourdough bread (Schwenninger and Meile, 2004). Dairy propionibacteria have a long history of safe use in human diet and animal feed. P. freudenreichii is widespread consumed in Swiss cheese in which they are present in concentrations close to 109 CFU g–1. Propionibacteria can synthesis vitamin B2, B7 (biotin), B9 and B12 and vitamin K (Burgess et al., 2009) beside homo and heteropolysaccharides (Dobruchowska et al., 2007) and produce propionic, acetic and succinic acid.
Therefore, this study aimed to isolate, identify and determine moulds and yeasts in kareish cheese samples, study the inhibitory effect of some propionibacteria strains against spoilage isolates and using them as bio-preservatives in kareish cheese.
MATERIALS AND METHODS
Sampling: Seventy two of kareish cheese samples were collected from milk shops, retail farmers, street vendors and supermarkets (18 each) from Dakahlia governorate. All samples were transported in an ice-box to laboratory and immediately analyzed on arrival to determine, isolate and identify of moulds and yeasts.
Microbial cultures: The strains of moulds and yeasts used in this study were isolated from kareish cheese samples. Aspergillus flavus and Candida albicans were selected to study their behavior in the presence of propionibacteria strains (PB) in kareish cheese. Propionibacterium acidipropionici 117, Propionibacterium jensenii 118 and Propionibacterium thoenni 119 were provided by the Institute of Food Biotechnology, Olsztyn University of Agriculture and Technology, Heweliusz, Olsztyn, Poland. The L. lactis subsp. lactis (LL) was obtained from Chr. Hansen’s Lab., Hoersholm, Denmark. Stock cultures of PB and LL were grown at 30°C in sodium lactate broth (NLB) and MRS broth, respectively and maintained at 4°C. Before using PB and LL were transferred three times at daily intervals in Enriched Whey Medium (EWM) and MRS broth, respectively.
Isolation of moulds and yeasts: According to Van der Walt and Yarrow (1984), 10 g of each sample was taken, diluted in 90 mL of sterile solution of 2% (w/v) sodium citrate and homogenized in a stomacher for 30 sec. For all samples, ten fold serial dilutions were prepared in a sterile solution of 2% (w/v) sodium citrate and the numbers of yeasts and moulds were determined by surface plating on yeast Potato Dextrose Agar (PDA) with 0.01% of chloramphenicol and Czapek-Dox’s medium, respectively. Each sample analyzed in duplicate, incubated at 25°C for 5 days. Moulds and yeasts colonies were purified, morphologically identified in mycological laboratory-botany department, Faculty of Science, Mansoura University and mentioned on selective media.
Assay of antimicrobial activity using agar well diffusion technique: Sterilized skimmed milk (12%) was separately inoculated with P. acidicpropionici 117, P. jensenii 118, P. thoenii 119 and L. lactis subsp. lactis (2%), incubated at 30°C for 48 h, centrifuged at 400 rpm to get a very clear supernatant, then sterilized by passing through sterile 0.45 μm syringe filter to obtain Cell Free Filtrate (CFF). Each filtrate was divided into 3 parts; the 1st part was taken as it is (CFF), the 2nd has been treated ammonium sulphate until full saturation, the mixtures were left overnight at 4.0°C, then the precipitation of protein were occurred by centrifugation for 15 min at 10000 rpm at 4.0°C. Any precipitated protein compounds (peptides and bacteriocins) pellets were dissolved immediately in a minimum volume of 0.1 M phosphate buffer at pH 7.0 then has been re-mitigation in sterile water (PPB). The 3rd part was heated at 121°C for 15 min to destroy the protein compounds, while retaining the effect of organic acids (HFF). Amounts of 20 mL of sterile nutrient agar were poured into sterile petri dishes. After solidification, 100 μL of fresh activity of culture suspensions of each mould or yeast isolates were swabbed on the respective plates. All parts of CFF, PPB and HFF carefully added in the perfect wells in agar. The plates were kept in the fridge for 2 h then incubated for suitable time (3-5 days) at 30°C. After incubation the diameter of inhibition zones formed around each well were measured in mm and recorded.
Manufacture of kareish cheese: Buffalo’s milk (The chemical composition of milk is given in Table 1) was defatted and heated at 80°C for 15 sec then cooled to 32°C. Milk was divided into 4 portions, cultures were added as follows: (1) L. lactis subsp. lactis (2% v/v), served as a control, (2) L. lactis subsp. lactis and P. acidicpropionici 117 (2%, 1:1 v/v) (LPA), (3) L. lactis subsp. lactis and P. jensenii 118 (2%, 1:1 v/v) (LPJ) and (4) L. lactis subsp. lactis and P. thoenii 119 (2%, 1:1 v/v) (LPT), 0.5% glucono-δ-lactone (GDL) was added to each treatment. Each portion was divided into two sub portion, A and B. All portions coded A were directly incubated at 30°C. All portions coded B inoculated with A. flavius and Candida albicans to give initial count of 103 CFU mL–1 then incubated at 30°C. Resultant cheeses were packed into plastic bags and stored at 6±1°C for 30 days. Samples of each cheese were taken, zero time, 5, 10, 15 and 30 days after manufacture for chemical composition, rheological, microbiological analysis and sensory evaluation. Data were reported as the average of three independent trials. Organoleptic evaluation was carried out according to the scheme of El-Shafei et al. (2008).
| Table 1: | Gross chemical composition of milk used for making kareish cheese |
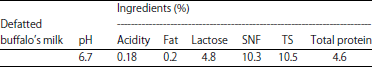 | |
Chemical analysis: Chemical analysis in this study were determined according to AOAC (2003). Determination of folate and vitamin B12, the cheese sample extraction procedure was carried out according to literature (Albala-Hurtado et al., 1997) to determine of water-soluble vitamins in infant milk using High Performance Liquid Chromatography (HPLC ) analysis. Acelatldehyde and diacetyl content of all cheeses were measured using a Shimadzu (240 UV-VIS) spectrameter (Japan) as describes by Lees and Jago (1970).
Determination of PB and LL: For Propionibacteria (PB) counts, NLB containing agar (15 g L–1) and bromocresol purple (0.032 g L–1) as an indicator with anaerobic incubation at 30°C was used. Both PB and LL grew on NLB agar. The LB formed big, white colonies after 48 h and PB formed small, yellow colonies with yellow zones after 96 h incubation (Babuchowski et al., 1999), Lac. lactis subsp. lactis (LL) also was enumerated on M 17 agar at 30°C for 24 h.
Texture determination: Texture profile analysis of the kareish cheese samples were carried out by using the texture analyzer (CNS-Farnell, England). Cheese samples were cut into cubes 5 cm 3 and kept at 12°C for 1 h before analysis. The probe was TA 15 (45° and 30 mm diameter), at speed 1 mm sec–1 and 10 mm distance, using cycle or hold programs. Hardness, cohesiveness, springiness, gumminess and adhesiveness were calculated as described by Szczesniak and Kleyn (1963) and Bourne (1978).
Data were subjected to statistical analysis by the computer program of using the General Linear Model (GLM).
RESULTS AND DISCUSSION
Determination of moulds and yeasts: The first step in this study was collection of kareish cheese samples from 4 sources to isolate, identify and determine counts of contaminated moulds and yeasts, listed results in Table 2, illustrated that samples of retail farmers and street vendors were the worst in mold and yeast counts, all these samples were contaminated with moulds (100%), while 17 out of 18 (94.4%) retail farmers samples were contaminated with yeast, street vendor samples were completely contaminated with yeasts.
| Table 2: | Positive kareish cheese samples for moulds and yeasts with mentioning the source and conformance with EOSQC (2005) |
 | |
| *Standards according to EOSQC (2005), Moulds ≤10 CFU g–1, Yeast ≤400 CFU g–1 N-C means Non-conforming | |
| Table 3: | Antimicrobial activity of dairy propionibacteria cell free against isolated moulds and yeasts from kareish cheese (the diameter of inhibition zone mm*) |
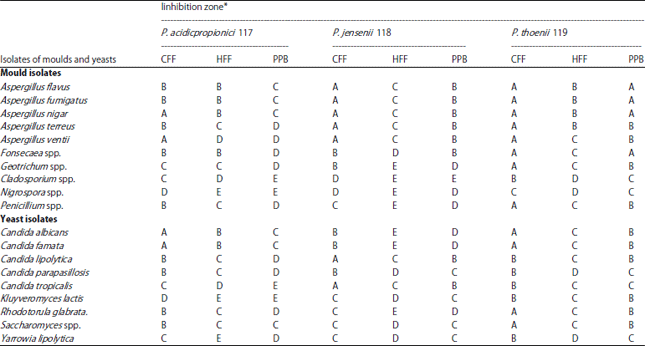 | |
*Diameter of the inhibition zone, E≤8 mm, D 9-13 mm, C 14- 18 mm, B 19- 22 mm, A 22-29 mm. Maximum inhibitory zone of L. lactis subsp. lactis was nearly 2 mm (data not shown), CFF: Crude cell free filtrate, HFF: Heated cell free filtrate and PPB: Precipitated peptides and bacteriocins | |
Milk shop and supermarket samples were positive for moulds at ratios 88.9 and 83.3%; the same samples included 77.8 and 72.2% contaminated samples with yeasts, respectively. It should be noted that positive samples contained 4-5 log10 CFU g–1 moulds or yeasts, thereby led to confirm that all positive sample were non-conforming because the mold and yeast counts exceeded 10 and 400 CFU g–1, respectively based on EOSQC (2005). These findings surely related to reduction of the hygiene condition during the manufacturing and trading. Counts of mould in kareish cheese were 8×108 CFU g–1 (average, 3×104 CFU g–1). So, 87% of kareish cheese samples were non-conforming due to high mycological counts exceeded 10 CFU g–1 mold or 400 CFU g–1 yeast, according to the Egyptian Standard (ES) 1008-2000 (El Sayed et al., 2011). Yeasts were isolated with an incidence of 52% from fresh kareish cheese samples (Hakim et al., 2013). Similar results were reported by Elbagory et al. (2014) who found that 100% kareish cheese samples were contaminated with moulds and all samples were non-conforming for EOSQC (2005). Abou Ayana et al. (2014) could isolate many toxigenic fungi from types of cheese.
Identification of moulds and yeasts: In Table 3 it could prove that there are 19 mould and yeast species belong to 11 fungal genera. Aspergillus and Candida were the most common genera recovered from the examined cheese samples. Aspergillus was represented by 5 mould species (A. flavus, A. fumigatus, A. nigar, A. terreus and A. ventii). Candida was represented by 5 yeasts species (C. albicans, C. famata, C. lipolytica, C. parapasillosis and C. tropicalis) too. Five other mould species (Fonsecaea spp., Geotrichum spp., Cladosporium spp., Nigrospora spp. and Penicillium spp.) were isolated. Four other yeast species (Kluyveromyces lactis, Rhodotorula glabrata, Saccharomyces spp. and Yarrowia lipolytica) were isolated and identified. The enormous counts and species of moulds and yeasts associated with the low pH, the nutritional profile of kareish cheese, which are favorable for the growth of these organisms in the presence of oxygen, surface moisture, often containing lactic acid, peptides and amino acids favors rapid growth. Similar results were recorded by Elbagory et al. (2014). Growth of Asperigillus, Cladosporium and Penicillium species may responsible for bitterness and rancidity of cheese. This great diversity of contaminated moulds and yeasts for the cheese surely carries future connotations of spoilage or healthy risks. Chemically, Geotrichum candidum reduced diacetyl concentrations by 52-56% in low fat cottage cheese after 15-19 days of storage at 4-7°C (Antinone and Ledford, 1993).
Organoleptically, yeasty, fermented off-flavors, mouldy and gassy appearance and sticky texture are often detected when yeast and mould grow to 5-6 log10 CFU g–1. Giudici et al. (1996) confirmed that galactose, which results from lactose hydrolysis by the lactic starter cultures was fermented by galactose-positive strains of yeasts such as Saccharomyces cerevisiae and Hansenula anomala. Lipolysis produces short-chain fatty acids that combine with ethanol to form fruity esters. Some proteolytic yeast strains produce sulfides, resulting in an egg odor. Common contaminating yeasts of cheeses include Candida spp., Kluyveromyces marxianus, Geotrichum candidum, Debaryomyces hansenii and Pichia spp. (Johnson, 2001).
Antagonism assay: The results in Table 3 also indicated to CFF of all tested propionibacteria strains had the strongest inhibitory effect against all isolated moulds and yeasts, PPB of P. thoenii 119 and P. jensenii 118 were higher inhibition than HFF, while HFF of P. acidicpropionici 117 was higher inhibition than PPB. Generally, there are not clear differences in the inhibitory effects from tested propionibacteria strains against mould and yeast isolates, these results confirmed by following -up the growth and behavior of A. flavus and C. albicans in the presence of propionibacteria cultures in kareish cheese during cold storage. In terms of efficiency, P. thoenii 119 was the most efficient in the inhibition all tested moulds and yeasts followed by P. jensenii 118 then P. acidicpropionici 117. Furthermore, protective cultures produce protein compounds (peptides and bacteriocins) and organic acids (e.g., acetic, propionic and succinic acids) that own anti-microbial properties. The P. acidicpropionici 117 has acidity inhibitory effect higher than bacteriocins unlike P. jensenii 118 and P. thoenii 119, which own higher bacteriocins effects than acidity inhibition. Propionibacterium shermanii and their metabolites use as a food grade biopreservative against fungi and Gram negative bacteria (Al-Zoreky et al., 1991). Bio profit contains viable cells of P. freudenreichii subsp. shermanii strain JS and is effective for inhibiting yeasts growth in dairy products, Bacillus spp. in sourdough bread (Suomalainen and Mayra-Makinen, 1999). El-Shafei et al. (2008) could inhibit Sacchromyces cerevisiae and Candida sp., using bacterial combinations contain lactic acid bacteria and P. thoenii P. 127. Bacteriocin propionicin PLG-1 and GBZ-1 produced by P. thoenii P-127 (Lyon et al., 1993), Propionicins SM1 and SM2 produced by P. jensenii DF1 (Miescher et al., 2000). Thoenicin 447 isolated from P. thoenii 447 (Van der Merwe et al., 2004). These bacteriocins are active against moulds (Aspergillus spp., Apiotrichum curvatum, Fusarium tricinctum and Phialophora gregata) and yeasts (Saccharomyces, Candida and Scopularopsis sp.). Lastly, propionibacteria also produce other peptides and organic acids (2-pyrrolidone-5-carboxylic acid, 3-phenyllactic acid, hydroxyphenyl lactic acid 3-phenyllactic acid) with antiviral, anti-yeasts and antifungal activities (Lind et al., 2007).
| Table 4: | Yield percentage, total points of sensory evaluation and changes in chemical composition of kareish cheese manufactured using some propionibacteria strains during storage at 6±1°C |
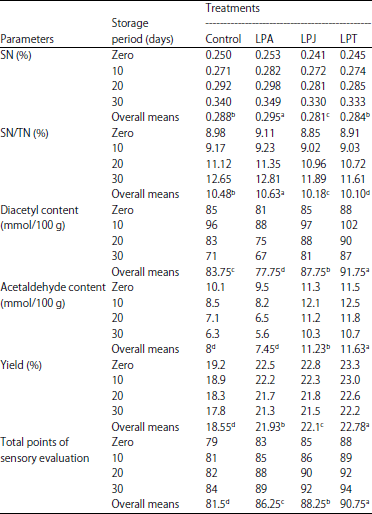 | |
LPA: P. acidicpropionici 117+L. lactis subsp. lactis, LPJ: P. Jensenii 118+L. lactis subsp. lactis and LPT: P. acidicpropionici 117+L. lactis subsp. lactis | |
Evaluation of cheese: Many factors involving milk composition, amount and genetic variants of casein, milk quality, milk pasteurization, coagulant type, vat design, curd firmness at cutting and manufacturing parameters influence on cheese yield. From Table 4 the results showed that the yield affected the cultures used, LPT produced the greatest yield (23.3%), which reached 22.2% at the end of storage period. This treatment followed by LPJ, LPA, then control that recorded the lowest yield. Generally, cheese yield gradually declined along storage period. Surely, these results due to the deference of cultures used particularly presence of propionibacteria, it can biosynthesis exopolysaccharides. Dobruchowska et al. (2007) confirmed ability of propionibacteria to biosynthesis exopolysaccharides.
Concerning ripening parameters, data in Table 4 show that both Soluble Nitrogen (SN) content and the SN/TN ratio had similar trends. No differences were found between the control and experimental cheeses at the first stages of storage indicating that the presence of protective cultures in kareish cheese did not contribute to primary proteolysis. During storage, a gradual increment in SN and SN/TN was observed in cheese from all treatments till the end of storage. Similar trends were reported for kareish cheese by El-Shafei et al. (2008).
Data listed in the same table show that kareish cheese produced with P. thoenii 119 and L. lactis subsp. lactis had the highest acetaldehyde and diacetyl contents, whether when fresh or stored. These increases may be due to both actions of aroma cultures and P. thoenii 119 metabolize all the citrate and produce appreciable amounts of diacetyl (Frohlich-Wyder et al., 2002). Both acetaldehyde and diacetyl contents increased to reach maximum values after 10 days then decreased until the end of storage in all treatments. The concentration of acetaldehyde and diacetyl can differ to a great extent, depending on the medium composition, growth conditions and the specific activity of bacteria and their enzymes. In cheese, the pathway leading to acetaldehyde production is generally considered to be via lactose degradation (Salem et al., 2007). Some of the end products of citrate and pyruvate metabolism, such as diacetyl and acetaldehyde have distinct aroma properties and contribute significantly to the quality of fermented foods (Helland et al., 2004). In general, these results coincided with those obtained by El-Shafei et al. (2008).
For sensory evaluation, the cheeses were evaluated for taste, flavour, body and texture and appearance during storage at 6°C are recorded in Table 4. According to the panelists, cheese made with LPT culture received more score points; it enhanced the flavor, body and texture of the treated kareish cheese compared with the control.
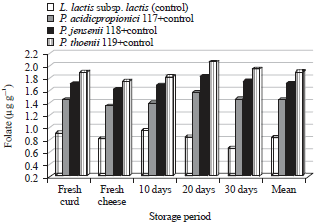 | |
| Fig. 1: | Folate (μg g–1) in kareish cheese made with L. lactis subsp. lactis or L. lactis subsp. lactis+Propionibacteria strains during storage periods |
This trend continued along storage. The LPJ treatment came in the second place in total points of sensory evaluation then LPA treatment which was lower flavor, appearance and body and texture, hence total score against treatments. All experimental cheeses had good appearance, flavor and texture and body, the yeasty and mouldy flavor disappeared opposite the control that had some changes, it was slightly sticky, had slightly yeasty taste and lower flavor. These changes due to the high load of moulds and yeasts which can hydrolysis diacetyl (Antinone and Ledford, 1993). Enhancing of body and texture in experimental cheeses due to exopolysaccharides that are produced by propionibacteria strains.
Vitamins B9 and B12: Dairy propionibacteria have the ability to biosynthesis a package of important and necessary vitamins for the host health, such as B2 (riboflavin), B7 (biotin), B9 (folic acid) and B12 (cobalamin), vitamin K (MK-9 (4H) and its precursor DHNA with bifidogenic activity. Folate or folic acid, is an important B-group vitamin, participates in many metabolic pathways, such as DNA and RNA biosynthesis and amino acid inter-conversions. Mammalian cells cannot synthesize folate; therefore, an exogenous supply of this vitamin is necessary to prevent nutritional deficiency (LeBlanc et al., 2007). Among lactic acid bacteria many Lactobacillus spp., Lactococcus spp., L. plantarum, L. bulgaricus, L. lactis, S. thermophilus and Enterococcus spp. have the ability to produce folate so it was necessary to determine folate. Figure 1 shows the levels of folate in fresh curd and cheese after 0, 10, 20 and 30 days of storage. Propionibacteria strains were used simultaneously with L. lactis subsp. lactis that was only used in control.
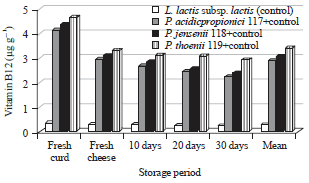 | |
| Fig. 2: | Vitamin B12 (μg g–1) in kareish cheese made with L. Lactis subsp. lactis or L. lactis subsp. lactis+Propionibacteria strains during storage periods |
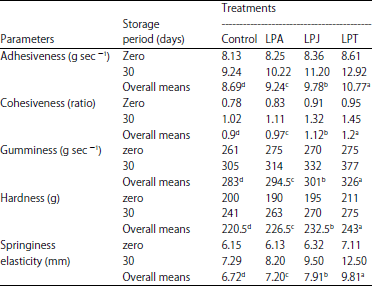
| Table 5: | Textural characteristics of Kareish cheese manufactured with and without protective cultures during storage at 6±1°C |
 | |
LPA: P. acidicpropionici 117+L. lactis subsp. lactis, LPJ: P. Jensenii 118+L. lactis subsp. lactis and LPT: P. acidicpropionici 117+L. lactis subsp. lactis | |
The results show significantly differences between the control and treatments. L. lactis subsp. lactis (control) produced a limited quantity of folate compared to the treatments. The LPT (control+P. thoenii 119) produced the greatest level of folate followed by LPJ (P. jensenii 118+control) then LPA (P. acidicpropionici 117+control) generally, folate slightly increased to reach the maximum level on the 20th day of storage at 6±1°C then decreased. Hugenholtz et al. (2002) found some propionibacteria strains as productive as Streptococcus thermophiles. Folate decreased in experimental fresh cheeses against fresh curds, fresh cheeses recorded the lowest folate levels. Furthermore, folate decreased on the 30th day but still more than in fresh cheese. These results nearly similar obtained results by Holasova et al. (2004) for fermented milks, Ibrahim et al. (2014) confirmed that some lactic acid bacteria isolated from Egyptian dairy products able to synthesize levels of folate. These increases of folate during storage are associated with the growth of propionibacteria.
Vitamin B12 or cobalamin is an essential nutrient for the human body that plays a key role in the normal functioning of the brain and nervous system, the formation of blood and also the metabolism of every cell, especially affecting DNA synthesis and regulation, fatty acid synthesis and energy production. Its deficiency leads to a serious physiological disorder called pernicious anemia. Many microorganisms such as bacteria able to produce vitamin B12 by fermentative processes. Propionibacteria produce vitamin B12 intracellularly and excrete mainly propionic acid and acetic acid extracellularly. In this study, remarkable differences in vitamin B12 production among the different starters used to make kareish cheeses are presented in Fig. 2. Production of vitamin B12 by single L. lactis subsp. lactis (C) or accompanied with P. acidicpropionici 117 (LPA), P. jensenii 118 (LPJ) and P. thoenii 119 (LPT) was determined. The LPT treatment was significantly higher in vitamin B12 production compared to control, LPA or LPA. Fresh curds contained the greatest vitamin B12 against fresh or stored cheeses. Sharp decline was noticed in vitamin B12 in fresh cheeses followed by a gradual increase till the 10th day of storage period. The quantities of vitamin B12 declined again on the 20th day and continued to decrease until the end of storage period. These results related to the moisture content of the curds and the growth of bacteria, Vitamin B12 is water- soluble so loses in the syneresis of whey. Furthermore, genetic differences of strains in ability to produce vitamin B12. Sharaf et al. (2014) obtained similar findings using encapsulated Propionibacterium shermanii or free cells.
Textural measurements: It was necessary to determine the textural parameters in cheese made with and without propionibacteria cultures (Table 5). Textural profile was determined at fresh and after 30 days. It is clear that all the textural properties gradually increased till the end of the storage period. This could be mainly due to proteolysis of casein to compounds that are very soluble in water and that do not contribute to the protein network responsible for the cheese rigidity. Similar findings in kareish cheese was reported by El-Shafei et al. (2008). All the textural properties of kareish cheese were influenced by using the protective cultures. Cheese made of protective cultures containing P. acidicpropionici 117, P. jensenii 118 or P. thoenii 119 were higher adhesiveness (g sec–1), cohesiveness (ratio), gumminess (g sec–1), hardness (g) and springiness elasticity (mm) than control which recorded the lowest texture profile parameters. The LPT treatment was the highest in textural parameters followed by LPJ, LPA. These results could be related to exopolysaccharides synthesis by protective cultures.
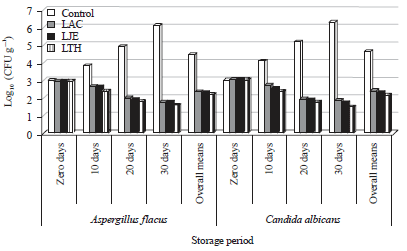 | |
| Fig. 3: | Behavior of A. flavus and C. albicans in kareish cheese in the presence of propionibacteria cultures |
| Table 6: | Changes of propionibacteria strains populations, L. lactis subsp. lactis, moulds and yeasts in kareish cheese during storage at 6±1°C |
 | |
| LPA: P. acidicpropionici 117+L. lactis subsp. lactis, LPJ: P. Jensenii 118+L. lactis subsp. lactis and LPT: P. acidicpropionici 117+L. lactis subsp. lactis | |
The differences observed in springiness and cohesiveness values may be attributed to the amount of protein matrix present and its strength. These results in agreement Abou Ayana and Ibrahim (2015).
Microbial properties of kareish cheese: In Table 6 LPA, LPJ and LPT had an inhibitory effect against moulds and yeasts have been shown. In control, moulds andyeasts could growth on the 10th day and rapidly increased along storage to reach the maximum populations among all treatments (1.27 and 2.32 log10 CFU g–1), respectively. In the presence of propionibacteria strains, LPT restricted and prohibited the growth of these organisms till 20 days, only 1.17, 1.42 and 1.32 log10 CFU g–1 of yeasts were detected after 20 days in LPT, LPJ and LPA, respectively. Moulds appeared in LPA after 20 days but were detected after 30 days in LPJ and LPT. At the end of storage period moulds and yeasts recorded (0.84 and1.34), (0.9 and 1.88) and (1.11and1. 63) log10 CFU g–1 in LPT, LPJ and LPA respectively. Nevertheless, all experimental cheeses were conformance with EOSQC (2005). Off-flavor, yeasty and moldy appearance disappeared. Thus, it is possible to prolong the shelf life of kareish cheese to month in refrigerator. On the other hand, propionibacteria strains slightly increased then decreased on the 20th day to reach 9.85, 9.97 and 9.95 log10 CFU g–1 for P. acidicpropionici 117, P. jensenii 118 and P. thoenii 119 at the end of cold storage, respectively. The L. lactis subsp. lactis took the same trend.
Antifungal activity in kareish cheese: The most isolated moulds and yeasts from collected samples were Aspergillus and Candida, therefore, Aspergillus flavus and Candida albicans were selected to further study. Data presented in Fig. 3 clearly showed the inhibition of both tested microorganisms in experimental kareish cheese made with propionibacteria cultures. The protective cultures (Propionibacteria) were able to totally inhibit the growth of A. flavus and C. albicans during storage for 30 days at 6±1°C. In the control (without propionibacteria strains), the growth of C. albicans was faster than A. flavus; it was 5.2 log CFU g–1 on 20th day. The LPJ and LPT treatments inhabited C. albicans more than A. flavus whilst LPA treatment inhabited A. flavus more than C. albicans. Generally, P. thoenii 119 could inhabit A. flavus and C. albicans more than P. jensenii 118 and P. acidicpropionici 117. The populations of A. flavus and C. albicans decreased by (1.41, 1.24 and 1.2) and (1.49, 1.28 and 1.1) log cycle in kareish cheese made with LPT, LPJ and LPA, respectively, after 30 days. Therefore, isolates of A. flavus and C. albicans did not clearly differ in their sensitivity to the tested protective cultures. These results reflect the antagonism effects of propionibacteria cultures against the mould and yeast, this antagonism resulting due to biosynthesis compounds of propionibacteria cultures that revealed by the results recorded in Table 3. The results of this research point was confirmed the results of Schwenninger and Meile (2004) and El-Shafei et al. (2008). The visible fungal growth occurred in the control cheese after 10 days, it became softer in texture and developed a less attractive odor, unacceptable and rejected at the end of the storage period.
CONCLUSION
Finally, the moulds and yeasts represent a nuisance to producers and consumers of soft cheese particularly manufactured by fermentation as kareish cheese. Using protective cultures contain propionibacteria strains such as P. thoenii 119, P. jensenii 118 and P. acidicpropionici 117 can be limiting the growth of moulds and yeasts allowing prolonging the validity of this type of cheese up to a month with improving sensory evaluation, microbial, rheological properties and increase the content of some of the necessary vitamins.
REFERENCES
- Abou Ayana, I.A.A. and A.E. Ibrahim, 2015. Attributes of low-fat yogurt and kareish cheese made using exopolysaccharides producing lactic acid bacteria. Am. J. Food Technol., 10: 48-57.
CrossRefDirect Link - Abou Ayana, I.A.A., A.A.G. El-Deen, K.M.K. Ayyad and M.A. El-Metwally, 2014. Influence of some abiotic anti-fungi and Saccharomyces cerevisiae on aflatoxin B1 and isolated fungi from some cheese types. Int. J. Dairy Sci., 9: 32-44.
CrossRefDirect Link - Albala-Hurtado, S., M.T. Veciana-Nogues, M. Izquierdo-Pulido and A. Marine-Font, 1997. Determination of water-soluble vitamins in infant milk by high-performance liquid chromatography. J. Chromatogr. A, 778: 247-253.
CrossRefDirect Link - Al-Zoreky, N., J.W. Ayres and W.E. Sandine, 1991. Antimicrobial activity of Microgard™ against food spoilage and pathogenic microorganisms. J. Dairy Sci., 74: 758-763.
CrossRefDirect Link - AOAC., 2003. Official Methods of Analysis of the Association of Official's Analytical Chemists. 17th Edn., Association of Official Analytical Chemists, Arlington, Virginia.
Direct Link - Babuchowski, A., L. Laniewska-Moroz and I. Warminska-Radyko, 1999. Propionibacteria in fermented vegetables. Le Lait, 79: 113-124.
Direct Link - Burgess, C.M., E.J. Smid and D. van Sinderen, 2009. Bacterial vitamin B2, B11 and B12 overproduction: An overview. Int. J. Food Microbiol., 133: 1-7.
CrossRefDirect Link - Dobruchowska, J.M., G.J. Gerwig, A. Babuchowski and J.P. Kamerling, 2007. Structural studies on exopolysaccharides produced by three different propionibacteria strains. Carbohydr. Res., 343: 726-745.
CrossRefDirect Link - El-Kest, M.M., M. El-Hariri, N.I.M. Khafaga and M.K. Refai, 2015. Studies on contamination of dairy products by aflatoxin M1 and its control by probiotics. J. Global Biosci., 4: 1294-1312.
Direct Link - El Sayed, M.A., I.M. Hosny, W.I. El Kholy, R.K. El Dairouty and S.H.S. Mohamed, 2011. Microbiological evaluation of Egyptian white soft cheeses style. J. Am. Sci., 7: 517-526.
Direct Link - Elbagory, A.M., M. Amal, A.M. Hammad and A. Salwa, 2014. Prevalence of fungi in locally produced cheese and molecular characterization of isolated toxigenic molds. Benha Vet. Med. J., 27: 9-20.
Direct Link - El-Shafei, K., M.A.M. Abd El-Gawad, N. Dabiza, O.M. Sharaf and B.A. Effat, 2008. A mixed culture of Propionibacterium thoenii P-127, Lactobacillus rhamnosus and Lactobacillus plantarum as protective cultures in Kareish cheese. Polish J. Food Nutr. Sci., 58: 433-441.
Direct Link - Hakim, A.S., S.M.A. Azza and M.H.S.E.A. Rasha, 2013. Isolation, biochemical identification and molecular detection of yeasts from kareish cheese. Int. J. Microbiol. Res., 4: 95-100.
Direct Link - Helland, M.H., T. Wicklund and J.A. Narvhus, 2004. Growth and metabolism of selected strains of probiotic bacteria in milk- and water-based cereal puddings. Int. Dairy J., 14: 957-965.
CrossRefDirect Link - Holasova, M., V. Fiedlerova, P. Roubal and M. Pechacova, 2004. Biosynthesis of folates by lactic acid bacteria and propionibacteria in fermented milk. Czech J. Food Sci., 22: 175-181.
Direct Link - Hugenholtz, J., J. Hunik, H. Santos and E. Smid, 2002. Nutraceutical production by propionibacteria. Le Lait, 82: 103-112.
Direct Link - Ibrahim, G.A., K. El-Shafei, H.S. El-Sayed and O.M. Sharaf, 2014. Isolation, identification and selection of lactic acid bacterial cultures for production of riboflavin and folate. Middle East J. Applied Sci., 4: 924-930.
Direct Link - LeBlanc, J.G., G.S. de Giori, E.J. Smid, J. Hugenholtz and F. Sesma, 2007. Folate Production by Lactic Acid Bacteria and other Food-Grade Microorganisms. In: Communicating Current Research and Educational Topics and Trends in Applied Microbiology, Mendez-Vilas, A. (Ed.). Formatex Research Center Publisher, Badajoz, Spain, ISBN: 9788461194223, pp: 329-339.
- Lind, H., J. Sjogren, S. Gohil, L. Kenne, J. Schnurer and A. Broberg, 2007. Antifungal compounds from cultures of dairy propionibacteria type strains. FEMS Microbiol. Lett., 271: 310-315.
CrossRefDirect Link - Lyon, W.J., J.K. Sethi and B.A. Glatz, 1993. Inhibition of psychrotrophic organisms by propionicin PLG-1, a bacteriocin produced by Propionibacterium thoenii. J. Dairy Sci., 76: 1506-1513.
CrossRefDirect Link - Alvarez-Martin, P., A.B. Florez, T.M. Lopez-Diaz and B. Mayo, 2007. Phenotypic and molecular identification of yeast species associated with Spanish blue-veined Cabrales cheese. Int. Dairy J., 17: 961-967.
CrossRefDirect Link - Metwalli, S.A.H., 2011. Extended shelf life of Kareish cheese by natural preservatives. Egypt. J. Agric. Res., 89: 639-649.
Direct Link - Miescher, S., M.P. Stierli, M. Teuber and L. Meile, 2000. Propionicin SM1, a bacteriocin from Propionibacterium jensenii DF1: Isolation and characterization of the protein and its gene. Syst. Applied Microbiol., 23: 174-184.
CrossRefDirect Link - Salem, M.M.E., Mona A.M. Abd El-Gawad, Fatma A.M. Hassan and B.A. Effat, 2007. The use of synbiotics for production of functional low fat Labneh. Pol. J. Food Nutr. Sci., 16: 151-159.
Direct Link - Schwenninger, S.M. and L. Meile, 2004. A mixed culture of Propionibacterium jensenii and Lactobacillus paracasei subsp. paracasei inhibits food spoilage yeasts. Syst. Applied Microbiol., 27: 229-237.
CrossRefPubMedDirect Link - Sharaf, O.M., K. El-Shafei, S.A. El-Gizawy, O.S. Barakat, F.A. Fathy and H.S. El-Sayed, 2014. Enhancement in vitamin B12 production by encapsulated Propionibacterium shermanii. Middle East J. Applied Sci., 4: 252-261.
Direct Link - Suomalainen, T.H. and A.M. Mayra-Makinen, 1999. Propionic acid bacteria as protective cultures in fermented milks and breads. Lait, 79: 165-174.
CrossRefDirect Link - Van der Merwe, I.R., R. Bauer, T.J. Britz and L.M.T. Dicks, 2004. Characterization of thoeniicin 447, a bacteriocin isolated from Propionibacterium thoenii strain 447. Int. J. Food Microbiol., 92: 153-160.
CrossRefDirect Link - Frohlich-Wyder, M.T., H.P. Bachmann and M.G. Casey, 2002. Interaction between propionibacteria and starter/non-starter lactic acid bacteria in Swiss-type cheeses. Le Lait, 82: 1-15.
Direct Link - Lee, G.J. and G.R. Jago, 1970. The estimation of diacetyl in the presence of other carbonyl compounds. J. Dairy Res., 37: 129-132.
CrossRefDirect Link