
Suharlina
Graduate School of Nutrition and Feed Science, Faculty of Animal Science, Bogor Agricultural University, Indonesia
Dewi Apri Astuti
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Bogor Agricultural University, Indonesia
Nahrowi
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Bogor Agricultural University, Indonesia
Anuraga Jayanegara
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Bogor Agricultural University, Indonesia
LiveDNA: 62.7299
Luki Abdullah
Department of Animal Nutrition and Feed Technology, Faculty of Animal Science, Bogor Agricultural University, Indonesia
International Journal of Dairy Science
Year: 2016 | Volume: 11 | Issue: 3 | Page No.: 100-105







ABSTRACT
This study aimed to evaluate nutritional quality of total mixed rations containing Indigofera zollingeriana for dairy goats by using in vitro rumen fermentation technique (RUSITEC). Four rations with different levels of I. zollingeriana were tested, i.e., 20% of dry matter (R1), 40% (R2), 60% (R3) and 80% (R4). Each ration was repeated three times by following a randomized complete block design in which different batch of rumen fluid served as the block. Variables observed included chemical composition, In Vitro Dry Matter Digestibility (IVDMD) and In Vitro Organic Matter Digestibility (IVOMD), methane emission, Volatile Fatty Acid (VFA) profiles and protozoa population. Results revealed that R4 contained significantly higher crude protein content than that of R1-R3 (p<0.05). Rations containing higher proportions of I. zollingeriana (60 and 80% dry matter) had significantly higher IVDMD and IVOMD as compared to their lower proportions (20 and 40% dry matter, p<0.05). Methane concentration was lowest in ration containing the highest proportion of I. zollingeriana and it was accompanied with the lowest protozoa population. It can be concluded that higher inclusion levels of I. zollingeriana in rations improved their nutritional values while decreasing methane emission as a main greenhouse gas.
PDF Abstract XML References Citation
Received: January 11, 2016;
Accepted: February 29, 2016;
Published: April 15, 2016
How to cite this article
Suharlina, Dewi Apri Astuti, Nahrowi, Anuraga Jayanegara and Luki Abdullah, 2016. Nutritional Evaluation of Dairy Goat Rations Containing Indigofera zollingeriana by Using in vitro Rumen Fermentation Technique (RUSITEC). International Journal of Dairy Science, 11: 100-105.
DOI: 10.3923/ijds.2016.100.105
URL: https://scialert.net/abstract/?doi=ijds.2016.100.105
DOI: 10.3923/ijds.2016.100.105
URL: https://scialert.net/abstract/?doi=ijds.2016.100.105
INTRODUCTION
Although milk production worldwide is dominated from dairy cow, some other species such as buffalo and goat also contribute to the supply of milk and they are significantly produced in some countries. With regard to goat milk, South Asian countries including India, Bangladesh and Pakistan are among the main producers of goat milk (Pal et al., 2011). Goat milk is believed to possess a number of advantages over cow milk, it provides more balance nutritional source for infants and can be used as a medicinal food due to its higher concentrations of health promoting components, which may be related to feeding pattern of goats in consuming large proportions of natural browses (Silanikove et al., 2010). It is also rich in beneficial fatty acid contents such as omega 3 fatty acids and trans-9 and trans-11 conjugated linoleic acids (Zervas and Tsiplakou, 2013; Chilliard et al., 2014). Thus, it is not surprising that the price of goat milk is much higher than that of cow milk and is considered as a premium food.
Dairy goats are important component of livestock and play a significant role in the socio-economic structure of rural poor. They are sometimes fed with poor quality feed given by small-holder farmers, which further lead to low milk production. Feeding of dairy goats with grasses is not sufficient to meet the nutritional requirement of high-producing dairy goats due to the low protein content in many grass species, particularly tropical grasses (Poppi and McLennan, 1995) in which the protein contents are normally ranging between 4-9% from Dry Matter (DM), while the protein requirement of dairy goat ration is 14% DM or more. Providing concentrate to fulfill the nutritional requirements of dairy goats may considerably improve their milk production (Bala et al., 2009; Giger-Reverdin et al., 2014; Gomes et al., 2015) but the price is unfortunately expensive and thus may not affordable for small-holder farmers. An alternative solution to reduce the use of concentrate for dairy goats is through utilization of legumes, such as Leucaena leucocephala, Gliricidia sepium or Flemingia macrophylla that rich in protein (Ondiek et al., 2000; Mui et al., 2002; Fagundes et al., 2014). Another potential legume for feeding of dairy goats is a forage namely Indigofera zollingeriana. The plant has a rapid growth in the defoliation interval of 60 days with a production of 51 t ha–1 year–1 (Abdullah, 2010). Indigofera zollingeriana is very adaptive to low fertility rates, easy on maintenance and low prize and high seed production potential throughout the season (Abdullah and Suharlina, 2010).
Since the plant has been rarely investigated so far, therefore, the purpose of this study was to evaluate nutrient contents, rumen fermentation and methane emission of dairy goat rations when supplemented with different levels of I. zollingeriana as performed by using RUSITEC.
MATERIALS AND METHODS
Sample preparation: All experimental procedures in the present study had been approved by the Faculty of Animal Science, Bogor Agricultural University, Indonesia. Leaf and twig samples of Indigofera zollingeriana were collected from University Farm Research Station, Darmaga, Bogor Agricultural University, Indonesia. The samples were obtained from three plots with the dimension of 4×6 m each. Each plot consisted of I. zollingeriana plants with the planting dimension of 1×1.5 m. After 68 days of planting period as recommended from our previous study (Abdullah and Suharlina, 2010) the plants were harvested and leaf and twig samples were collected. Approximately, 3 kg dry matter of leaf and twig samples were collected from each plot. The samples were then air-dried in a greenhouse for 3 h and subsequently oven-dried at 70°C for another 3 h. The materials were ground to pass a 2 mm sieve size and mixed homogeneously for further in vitro RUSITEC incubation and chemical analyses.
In vitro incubation: Prior to in vitro incubation, the ground I. zollingeriana samples were formulated into experimental rations at varying proportions, i.e., 20, 40, 60 and 80% DM and named as R1, R2, R3 and R4, respectively (Table 1). Other feedstuffs used to formulate the rations were napier grass, rice bran, ground corn, soybean meal, CaCO3, NaCl and premix. The in vitro incubation was performed by using RUSITEC apparatus equipped with eight vessels (Sanshin, Tokyo, Japan) as described by Kajikawa et al. (2003). Experiment was performed in three periods and each period lasted for 10 days, comprised of 7 days adaptation period and 3 days data collection period. Rumen fluid was obtained from three lactating Etawa dairy goats and drawn before morning feeding through stomach tube technique.
| Table 1: | Feed ingredients of the experimental rations (in dry matter percentage) |
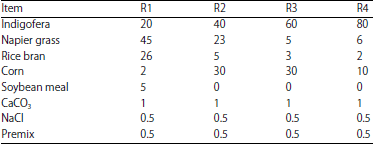 | |
| R1: Ration containing 20% DM Indigofera zollingeriana, R2: Ration containing 40% DM Indigofera zollingeriana, R3: Ration containing 60% DM Indigofera zollingeriana and R4: Ration containing 80% DM Indigofera zollingeriana | |
Rumen fluid was filtered through four layers of gauze before being used. An amount of 15 g DM of each ration was put into a nylon bag (100 μm pore size). Each vessel was filled with 400 mL strained rumen fluid and 400 mL McDougall buffer and the ration was also put into the vessel. About 70 g of solid rumen content was put into a nylon bag, serving as microbial inoculum for the initial 24 h. After that, the samples were incubated for every 48 h and replaced with similar samples throughout the experimental period. During the data collection period, fluid samples were measured for VFA profiles and protozoa count and feed residual samples in the nylon bags were determined for degradability values (both IVDMD and IVOMD). Fermentation gas was collected in a gas-proof bag for each vessel and measured for methane concentration by using an infrared methane analyzer (Sable System MA-10a, USA).
Pre and post-incubation analyses: Dry Matter (DM), Organic Matter (OM), Crude Protein (CP), Ether Extract (EE) and Crude Fiber (CF) composition of the rations were determined by proximate analysis (AOAC., 2000). Feed residuals were analyzed for DM and OM to obtain IVDMD and IVOMD values, respectively. Analysis of VFA was performed by injecting fermentation fluid samples into a gas chromatograph by following the description of Jayanegara et al. (2015). Protozoa population was counted by using a burker counting chamber (Blau Brand, Wertheim, Germany).
Statistical analysis: The data obtained were analyzed using analysis of variance (ANOVA) with the following statistical model:
where, Yij is the observed value, μ is the overall mean, τi is the effects of rations containing varying levels of I. zollingeriana (fixed effect), βj is the block effect (fixed effect) and εij is the random residual error. If there was a significant difference among the rations for a particular parameter at p<0.05, the analysis was continued a post hoc test i.e., Least Significance Difference (LSD) test. All the statistical analyses were performed by using SPSS statistical software version 20 (IBM Corp., Armonk, New York, USA).
RESULTS AND DISCUSSION
The CP content of R4 was significantly higher than that of R1, R2 and R3 (p<0.05, Table 2). Rations of R1 and R2 had significantly higher CF contents as compared to R3 (p<0.05). Other chemical constituents were similar among the four rations.
| Table 2: | Chemical composition of the experimental rations (in dry matter percentage) |
 | |
a,b: Different superscripts within the same row are significantly different at p<0.05, R1: Ration containing 20% DM Indigofera zollingeriana, R2: Ration containing 40% DM Indigofera zollingeriana, R3: Ration containing 60% DM Indigofera zollingeriana and R4: Ration containing 80% DM Indigofera zollingeriana | |
| Table 3: | Degradability, Volatile Fatty Acid (VFA) profiles and protozoa population of the experimental rations |
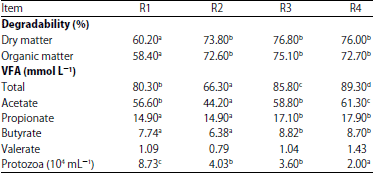 | |
a,b,c: Different superscripts within the same row are significantly different at p<0.05, R1: Ration containing 20% DM Indigofera zollingeriana, R2: Ration containing 40% DM Indigofera zollingeriana, R3: Ration containing 60% DM Indigofera zollingeriana and R4: Ration containing 80% DM Indigofera zollingeriana | |
With regard to CP and CF contents, R3 and R4 are recommended to satisfy dairy goats nutrient requirements for optimal milk production due to high CP and low CF. Higher inclusion levels of I. zollingeriana apparently improve nutritional properties of rations especially the CP contents since the plant contained high level of CP i.e., 29.2% DM. Similar to the finding of high CP content in I. zollingeriana, Tscherning et al. (2006) reported that the plant species had 23.8% DM of CP. Other Indigofera species such as I. arrecta and I. tinctoria were also reported to have high CP contents i.e., 23.1 and 26.0% DM, respectively (Kaitho et al., 1997; Bhatta et al., 2013).
The IVDMD and IVOMD values of R1 were lower than those of R2-R4 (p<0.05, Table 3). At low level of I. zollingeriana proportion as in R1, the ration contained high level of fiber at simultaneously low level of protein. Consistent with the results obtained in the present study, high fiber and low protein have been observed to be negatively correlated with digestibility coefficients in a number of domestic ruminant species i.e., sheep, goat, cattle and buffalo (Riaz et al., 2014). It is well accepted that protein is more digestible as compared to fiber by rumen microbes. Since, I. zollingeriana is characterized by its high protein and low fiber contents, increasing amount of the plant within a ration leads to an increase in its digestibility.
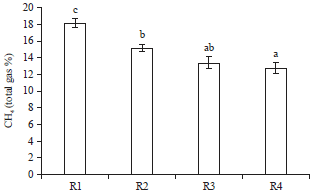 | |
| Fig. 1: | Methane (CH4) emission the experimental rations. Error bars indicate standard error values for the rations, different letters above the error bars are significantly different at p<0.05, R1: Ration containing 20% DM Indigofera zollingeriana, R2: Ration containing 40% DM Indigofera zollingeriana, R3: Ration containing 60% DM Indigofera zollingeriana and R4: Ration containing 80% DM Indigofera zollingeriana and DM: Dry matter |
The study of Kaitho et al. (1997) revealed that degradability of Indigofera arrecta measured at 24 h after incubation was higher than other commonly used high-protein legumes such as Calliandra calothyrsus, Flemingia macrophylla, Gliricidia sepium and Leucaena leucocephala. Further, increasing proportion of I. zollingeriana over C. calothyrsus increased the rate of in vitro gas production and apparently degraded N (Tscherning et al., 2006). These results may suggest the potentiality of Indigofera as a protein supplement in ruminant diets including those of dairy goats.
Production of total VFA was in line with IVDMD and IVOMD, in which higher proportion of I. zollingeriana led to an increase in total VFA production, especially the acetate and propionate (except for R2). Rumen VFA is produced as a result of microbial degradation and fermentation on feed entering the rumen. Thus it is unsurprising that VFA is positively correlated with feed degradability. Both acetate and propionate are required for milk production in dairy animals such as dairy cows and dairy goats; acetate is important for milk fat synthesis, whereas propionate is required for milk production (Sutton et al., 2003; Serment et al., 2011).
In vitro methane emission obtained in this study ranged from 12.8-18.2% from total gas, being highest in R1 and lowest in R4 (Fig. 1). The figure indicated that higher proportion of I. zollingeriana in diet led to a lower methane emission. This suggests that the plant is beneficial not only in term of improving animal production, but it also contributes to environmental conservation. Methane is considered as a main greenhouse gas after carbon dioxide although its capacity to retain heat is more than 20 folds than that of carbon dioxide. Accumulation of these greenhouse gas contribute to global warming and increase the earth’s temperature (Moss et al., 2000; Hristov et al., 2013). Methanogenesis in the rumen occurs due to a reduction of carbon dioxide and hydrogen by the action of archaea methanogen (Morgavi et al., 2010). Part of the methanogen is living in symbiosis with protozoa especially on the surface of the fauna (Tokura et al., 1997; Xia et al., 2014), methanogen gets benefit from protozoa through inter-species hydrogen transfer. Thus among the strategies to mitigate ruminal methane emission are: (1) Decreasing population and/or activity of methanogen, (2) Lowering the utilization of carbon dioxide and hydrogen by methanogen and (3) Defaunation of protozoa (Hegarty, 1999).
Lower methane emission with increasing proportion of I. zollingeriana in ration is partly due to its lower fiber content. Lower fiber is associated with lower acetate to propionate (A:P) ratio since fiber fermentation leads to more acetate formation rather than that of propionate. In the present study, A:P ratio in R1 was 3.80 and decreased to 3.42 in R4, this supports the above explanation. Other explanation of lower methane emission by the plant was due to lower protozoa population in ration with high I. zollingeriana proportion as can be seen in Table 3. Protozoa population decreased by 53.8, 58.8 and 77.1% in R2, R3 and R4, respectively, in comparison with R1 (p<0.05). Anti-protozoa effect of I. zollingeriana might be due to the presence of secondary compounds in the plant. It was analyzed that the plant used contained tannins and saponins i.e., 2.9 g kg–1 and 2.6 mg kg–1 DM, respectively. Tannins and saponins have been known to possess anti-protozoa effects and may be used as defaunating agents (Wang et al., 2009; Bhatta et al., 2012; Jayanegara et al., 2014). Such amounts of tannins and saponins in I. zollingeriana are considered to be relatively low in comparison to other plants (Jayanegara et al., 2012, 2014). These substances may negatively influence digestion processes in ruminants by acting as anti-nutritive factors when present at high concentrations. Low to moderate concentrations of these compounds are desirable in order to positively modulate rumen fermentation for higher animal productivity and lower environmental pollution.
CONCLUSION
Inclusion of I. zollingeriana improved nutritional value of dairy goat ration by increasing its crude protein content as well as its digestibility. Together with excellent agronomy characteristics of the plant, I. zollingeriana is a potent legume for supporting optimum production of dairy goat. It has been proven not only beneficial in term of animal production, but it also advantageous with regard to environmental conservation by mitigating enteric methane emission.
ACKNOWLEDGMENT
All authors are grateful to Directorate General of Higher Education of the Republic of Indonesia for providing a research grant under the scheme of "Riset Andalan Perguruan Tinggi dan Industri (RAPID)".
REFERENCES
- Abdullah, L., 2010. Herbage production and quality of shrub Indigofera treated by different concentration of foliar fertilizer. Media Peternakan, 33: 169-175.
Direct Link - Abdullah, L. and Suharlina, 2010. Herbage yield and quality of two vegetative parts of Indigofera at different times of first regrowth defoliation. Media Peternakan, 33: 44-49.
Direct Link - AOAC., 2000. Official Methods of Analysis of AOAC International. 17th Edn., Association of Official Analytical Chemists, Gaithersburg, Maryland.
Direct Link - Bala, P., R. Malik and B. Srinivas, 2009. Effect of fortifying concentrate supplement with fibrolytic enzymes on nutrient utilization, milk yield and composition in lactating goats. J. Anim. Sci., 80: 265-272.
CrossRefDirect Link - Bhatta, R., M. Saravanan, L. Baruah and K.T. Sampath, 2012. Nutrient content, in vitro ruminal fermentation characteristics and methane reduction potential of tropical tannin-containing leaves. J. Sci. Food Agric., 92: 2929-2935.
CrossRefDirect Link - Bhatta, R., M. Saravanan, L. Baruah, K.T. Sampath and C.S. Prasad, 2013. Effect of plant secondary compounds on in vitro methane, ammonia production and ruminal protozoa population. J. Applied Microbiol., 115: 455-465.
CrossRefPubMedDirect Link - Chilliard, Y., P.G. Toral, K.J. Shingfield, J. Rouel, C. Leroux and L. Bernard, 2014. Effects of diet and physiological factors on milk fat synthesis, milk fat composition and lipolysis in the goat: A short review. Small Rumin. Res., 122: 31-37.
CrossRefDirect Link - Fagundes, G.M., E.C. Modesto, C.E.M. Fonseca, H.R.P. Lima and J.P. Muir, 2014. Intake, digestibility and milk yield in goats fed Flemingia macrophylla with or without polyethylene glycol. Small Rumin. Res., 116: 88-93.
CrossRefDirect Link - Giger-Reverdin, S., K. Rigalma, M. Desnoyers, D. Sauvant and C. Duvaux-Ponter, 2014. Effect of concentrate level on feeding behavior and rumen and blood parameters in dairy goats: Relationships between behavioral and physiological parameters and effect of between-animal variability. J. Dairy Sci., 97: 4367-4378.
CrossRefPubMedDirect Link - Gomes, L.C., C.R. Alcalde, G.T. Santos, A.C. Feihrmann, B.S.L. Molina, P.A. Grande and A.A. Valloto, 2015. Concentrate with calcium salts of fatty acids increases the concentration of polyunsaturated fatty acids in milk produced by dairy goats. Small Rumin. Res., 124: 81-88.
CrossRefDirect Link - Hegarty, R.S., 1999. Reducing rumen methane emissions through elimination of rumen protozoa. Aust. J. Agric. Res., 50: 1321-1327.
CrossRefDirect Link - Hristov, A.N., J. Oh, J.L. Firkins, J. Dijkstra and E. Kebreab et al., 2013. SPECIAL TOPICS-Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci., 91: 5045-5069.
CrossRefDirect Link - Kajikawa, H., H. Jin, F. Terada and T. Suga, 2003. Operation and characteristics of newly improved and marketable artificial rumen (Rusitec). Mem. Natl. Inst. Livest. Grassland Sci., 2: 1-49.
Direct Link - Jayanegara, A., M. Kreuzer and F. Leiber, 2012. Ruminal disappearance of polyunsaturated fatty acids and appearance of biohydrogenation products when incubating linseed oil with alpine forage plant species in vitro. Livest. Sci., 147: 104-112.
CrossRefDirect Link - Jayanegara, A., E. Wina and J. Takahashi, 2014. Meta-analysis on methane mitigating properties of saponin-rich sources in the rumen: Influence of addition levels and plant sources. Asian-Australasian J. Anim. Sci., 27: 1426-1435.
CrossRefDirect Link - Jayanegara, A., G. Goel, H.P.S. Makkar and K. Becker, 2015. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol., 209: 60-68.
CrossRefDirect Link - Kaitho, R.J., I.V. Nsahlai, B.A. Williams, N.N. Umunna, S. Tamminga and J. van Bruchem, 1997. Relationships between preference, rumen degradability, gas production and chemical composition of browses. Agrofor. Syst., 39: 129-144.
CrossRefDirect Link - Morgavi, D.P., E. Forano, C. Martin and C.J. Newbold, 2010. Microbial ecosystem and methanogenesis in ruminants. Animal, 4: 1024-1036.
CrossRefPubMedDirect Link - Moss, A.R., J.P. Jouany and J. Newbold, 2000. Methane production by ruminants: Its contribution to global warming. Annales Zootechnie, 49: 231-253.
CrossRefDirect Link - Mui, N.T., I. Ledin, P. Uden and D. van Binh, 2002. The foliage of flemingia (Flemingia macrophylla) or jackfruit (Artocarpus heterophyllus) as a substitute for a rice bran-soya bean concentrate in the diet of lactating goats. Asian-Austr. J. Anim. Sci., 15: 45-54.
CrossRefDirect Link - Ondiek, J.O., J.K. Tuitoek, S.A. Abdulrazak, F.B. Bareeba and T. Fujihara, 2000. Use of Leucaena leucocephala and Gliricidia sepium as nitrogen sources in supplementary concentrates for dairy goats offered Rhodes grass hay. Asian-Aust. J. Anim. Sci., 13: 1249-1254.
Direct Link - Pal, U.K., P.K. Mandal, V.K. Rao and C.D. Das, 2011. Quality and utility of goat milk with special reference to India: An overview. Asian J. Anim. Sci., 5: 56-63.
CrossRefDirect Link - Poppi, D.P. and S.R. McLennan, 1995. Protein and energy utilization by ruminants at pasture. J. Anim. Sci., 73: 278-290.
Direct Link - Riaz, M.Q., K.H. Sudekum, M. Clauss and A. Jayanegara, 2014. Voluntary feed intake and digestibility of four domestic ruminant species as influenced by dietary constituents: A meta-analysis. Livestock Sci., 162: 76-85.
CrossRefDirect Link - Serment, A., P. Schmidely, S. Giger-Reverdin, P. Chapoutot and D. Sauvant, 2011. Effects of the percentage of concentrate on rumen fermentation, nutrient digestibility, plasma metabolites and milk composition in mid-lactation goats. J. Dairy Sci., 94: 3960-3972.
CrossRefPubMedDirect Link - Silanikove, N., G. Leitner, U. Merin and C.G. Prosser, 2010. Recent advances in exploiting goat's milk: Quality, safety and production aspects. Small Rumin. Res., 89: 110-124.
CrossRefDirect Link - Sutton, J.D., M.S. Dhanoa, S.V. Morant, J. France, D.J. Napper and E. Schuller, 2003. Rates of production of acetate, propionate and butyrate in the rumen of lactating dairy cows given normal and low-roughage diets. J. Dairy Sci., 86: 3620-3633.
CrossRefDirect Link - Tokura, M., K. Ushida, K. Miyazaki and Y. Kojima, 1997. Methanogens associated with rumen ciliates. FEMS Microbiol. Ecol., 22: 137-143.
CrossRefDirect Link - Tscherning, K., C. Lascano, E. Barrios, R. Schultze-Kraft and M. Peters, 2006. The effect of mixing prunings of two tropical shrub legumes (Calliandra houstoniana and Indigofera zollingeriana) with contrasting quality on N release in the soil and apparent N degradation in the rumen. Plant Soil, 280: 357-368.
CrossRefDirect Link - Wang, C.J., S.P. Wang and H. Zhou, 2009. Influences of flavomycin, ropadiar, and saponin on nutrient digestibility, rumen fermentation and methane emission from sheep. Anim. Feed Sci. Technol., 148: 157-166.
CrossRefDirect Link - Xia, Y., Y.H. Kong, R. Seviour, R.J. Forster, S. Kisidayova and T.A. McAllister, 2014. Fluorescence in situ hybridization probing of protozoal Entodinium spp. and their methanogenic colonizers in the rumen of cattle fed alfalfa hay or triticale straw. J. Applied Microbiol., 116: 14-22.
CrossRefPubMedDirect Link