
Nilufar Haque
Department of Veterinary Physiology and Biochemistry, College of Veterinary Science and Animal Husbandry, SDAU, SK Nagar, 385505, Gujarat, India
Mahendra Singh
National Dairy Research Institute, Karnal, Haryana, India
S.A. Hossain
National Dairy Development Board, Anand, Gujarat, India
International Journal of Dairy Science
Year: 2015 | Volume: 10 | Issue: 3 | Page No.: 95-106







ABSTRACT
Animal husbandry plays an important role in strengthening the rural economy. Heat is a major constraint on animal productivity in the tropical belt and arid areas. Heat stress in dairy cows as all temperature-related forces encourage changes or adjustments which may occur from the cellular to the total animal level to help the cows stay away from physiological disorders and then to better adapt to an adverse thermal environment. Milk production is impaired as a result of the drastic changes in biological functions under heat stress. Activation of the hypothalamus-pituitary-adrenocortical axis by external stress liberates cortisol into blood plasma, which in turn induces the liberation of Plasminogen Activator (PA) from the mammary epithelial cells, where it activates the Plasminogen-Plasmin (PG-PL) system. Thus, this PA-PG-PL system is a milk-borne factor which down regulates the milk secretion during the summer stress. This negative feedback system specifically forms β-casein (CN) fragment (f) (1-28) from β-CN, which acts as the negative control signal of mammary gland. Based on these findings, casein hydrolyzate preparation is developed to reduce suffering from mammary gland engorgement associated with abrupt cessation of milking and to treat common clinical and subclinical infections of udder in dairy cows.
PDF Abstract XML References Citation
Received: April 27, 2015;
Accepted: June 11, 2015;
Published: June 29, 2015
How to cite this article
Nilufar Haque, Mahendra Singh and S.A. Hossain, 2015. PA-PG-PL System in Dairy Animals and its Significance: A Review. International Journal of Dairy Science, 10: 95-106.
DOI: 10.3923/ijds.2015.95.106
URL: https://scialert.net/abstract/?doi=ijds.2015.95.106
DOI: 10.3923/ijds.2015.95.106
URL: https://scialert.net/abstract/?doi=ijds.2015.95.106
INTRODUCTION
India has been the largest milk producing country in the world since, 1997 and the recently announced data for 2013-14 with an estimated milk production of 137.6 million ton reiterated the strong position of the country in the global market. The country’s share in world milk production stands at 18% (NDDB., 2014). A large number of farmers in India depend on animal husbandry for their livelihood. Dairy farming provides supplementary employment, an additional source of income to many small and marginal farmers, household nutrition security. Thus, animal husbandry plays an important role in strengthening the rural economy. Over the last five years, India’s milk production has increased by over 4.0% (21 million ton) compared to an increase of about 2% increase in world milk production reported by FAO.
However, heat is a major constraint on animal productivity in the tropical belt and arid areas. Heat stress is a heavy load for the cow’s zoo technical performance and health status that costs the dairy industry millions of dollars every year. In temperatures above 28°C, even without humid conditions, lactating cows show evidence of hyperthermia along-with reduced feed intake, milk yield, milk fat and protein production, as well as fertility rate (West, 2003). High-yielding cows are affected more than low-yielding ones (Maltz et al., 2000; Barash et al., 2001) because the upper critical temperature shifts downward as milk production increase (Silanikove, 2000).
According to IPCC predictions, the global average surface temperature may increase between 1.8 and 4°C by year 2100 (IPCC., 2007). The negative effects of global warming will be manifest in animal agriculture of both developed and developing countries, but the pressure will be greater on developing countries because of their deficiency of resources, their lack of veterinary and extension services and their limitations on research technology development (FAO., 2007). Using simulation models in a dairy basin located in Australia, researchers estimated that by 2025, production of greenhouse gas emissions will increase 25% heat stress days, which could account for a decline of 35-210 kg of milk per cow per year. Projecting this scenario by year 2050, there will be a 60% increase in heat stress days, which may result in a decline from 85-420 kg of milk per cow per year (Crimp et al., 2010). Hot weather costs dairy farmers $ 900/million per year considering milk production and fertility. This economic loss is higher in dairy cattle compared to any other livestock species (St-Pierre et al., 2003). Thus, this PA-PG-PL system is a milk-borne factor which down regulates the milk secretion during the summer stress.
Heat stress and PA-PG-PL (Plasminogen activator-plasminogen-plasmin) system: Heat stress in dairy cows is all temperature-related forces that encourage changes or adjustments, which may occur from the cellular to the total animal level to help the cows stay away from physiological disorders and then to better adapt to an adverse thermal environment (Kadzere et al., 2002). Acclimation involves phenotypic responses to environmental changes, which are reflected in hormonal signals and also in alterations in target tissue responsiveness to hormonal stimuli (Horowitz, 2001; Collier et al., 2006). The time required for acclimation varies according to tissue types and ranges from a few days to several weeks (Rhoads et al., 2005; Collier et al., 2008).
Some external environmental stresses, such as dehydration (Silanikove, 1994) and acute HS (Maltz and Silanikove, 1996; Silanikove, 2000) may very quickly (within 24-48 h) take animals beyond their current acclimatized-adaptive range, which necessitates the induction of emergency physiological responses in order to avoid sudden death. Such immediate measures include an acute reduction in milk yield in high-yielding dairy cows, to protect of these external stresses (Silanikove, 1994, 2000). Hence, the milk secretion and mammary functions are regulated acutely by local autocrine feedback mechanisms that involve milk-borne factors, sensitive to the frequency and efficiency of milking (Daly et al., 1993). This negative feedback system was shown to comprise an endogenous milk enzymatic system, the Plasminogen Activator (PA)-Plasminogen (PG)-Plasmin (PL).
PA-PG-PL system in milk: Lactation is central to mammalian reproduction and critical for survival of the neonate. Milk synthesis starts in the epithelial cells of the mammary gland at the end of pregnancy to support the nutrition and promote the health of the off-spring. Milk contain vital nutrients, such as proteins, carbohydrates, lipids, minerals and vitamins, together with bioactive substances including immunoglobulins, peptides, antimicrobial factors, hormones, growth factors and enzymes (Clare and Swaisgood, 2000).
The principal enzyme in milk is plasmin, a heat-stable alkaline serine proteinase with a relatively broad specificity on the caseins; it exists in milk as a component of a complex system, including its zymogen, plasminogen, Plasminogen Activators (PAs) and inhibitors of both plasmin and PAs. Plasmin originates from blood and enters milk either in soluble form and then associates with the casein micelles. There are effects of factors, such as lactation, seasonality, stress, diet and mastitis on the plasmin system (Bastian and Brown, 1996; Kelly and McSweeney, 2003). This system is particularly intensively studied in respect to its role in the lysis of blood clots (Sidelmann et al., 2000) and regulation of cell activity and cell attachment in various tissues (Saksela and Rifirin, 1988). The involvement of this system in various activities reflects the ubiquitous presence of plasminogen in biological fluids and the ability of numerous cell types to synthesize, in a highly regulated manner, PA and inhibitors of plasmin and PA.
However, this autocrine factor may also affect mammary cell differentiation by modulating the number of cell surface hormone receptors for prolactin, thereby changing their sensitivity to circulating hormones (Wilde and Peaker, 1990). Sustained changes in the frequency of milking and milk secretion are associated with longer-term adaptation in the degree of differentiation and ultimately, the number of mammary epithelial cells. In addition to the regulation by milking, milk secretion also depends on external factors, such as emotional stress, as well as harsh physical conditions, such as heat stress and water deprivation (Silanikove, 2000).
Components of PA-PG-PL system
Plasmin: It is the principal proteolytic enzyme in milk (a serine proteinase with trypsin-like activity), is found mainly in its inactive or zymogen form, plasminogen. Plasmin concentration in fresh milk is 0.1-0.7 μg mL-1. The plasminogen to plasmin ratio in milk is estimated at 50:1-2:1. Plasmin preferentially cleaves polypeptide chains bonds of the type Lys-X and to a lesser extent, Arg-X (Ueshima et al., 1996). Plasmin has a pH optimum of 7.5.
Plasminogen: Plasminogen is a single-chain glycoprotein with a molecular mass of ~ 92 kDa. This is the zymogen for the proteolytically active glycoprotein, plasmin. Plasminogen concentration in fresh milk (reports of 0.8-2.8 mg mL-1). The plasminogen molecule (Fig. 1 for a model of plasminogen) consists of an amino-terminal pre-activation peptide (PAP, Fig. 2) followed by five characteristic kringle structures and the serine proteinase domain containing the catalytic triad (Sottrup-Jensen et al., 1978). Plasminogen is synthesized mainly in the liver and this inactive form circulates in the vasculature but is also found ubiquitously in other body fluids.
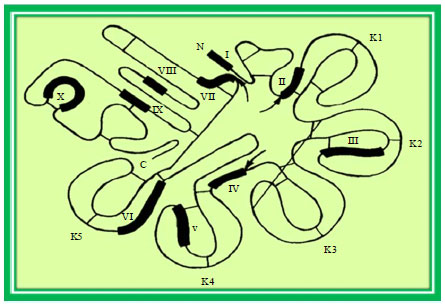 | |
| Fig. 1: | Schematic diagram for plasminogen molecule |
 | |
| Fig. 2: | Amino acid sequences identified in plasminogen from bovine milk |
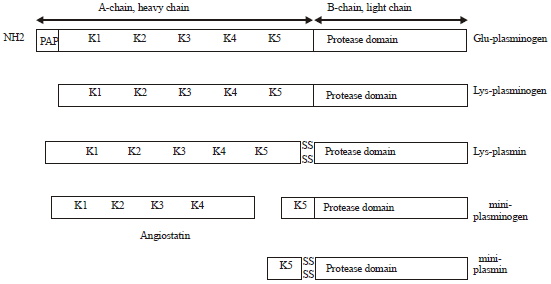 | |
| Fig. 3: | Schematic diagram of plasminogen and its cleavage products, PAP: Pre-activation peptide, K: Kringle, Glu: Glutamic acid, Lys: Lysine |
Plasminogen activator: The conversion of plasminogen to plasmin is modulated by PA, two types of which exist in mammals: Urokinase type PA (u-PA) and tissue-type PA (t-PA) (Heegaard et al., 1994). Plasmin, plasminogen and t-PA are closely associated with the casein micelles, whereas u-PA is associated with neutrophils (Politis et al., 2002, 2003). The close proximity of plasmin to its substrate ensures that hydrolysis is an efficient process and it is the primary agent of proteolysis in good quality milk (Kelly and McSweeney, 2003). The association of t-PA with the casein micelle suggests that it plays a pivotal role in regulating plasmin activity under nonpathological conditions. However, during bacterial infection, neutrophil numbers increase in the mammary gland and u-PA activity increases (Leitner et al., 2004).
Conversion of plasminogen to plasmin: Plasminogen is activated by a tPA or uPA catalyzed proteolytic cleavage of the Arg561-Va1562 bond located between the fifth kringle domain and the catalytic domain, combined with an autocatalytic cleavage of the Lys77-Lys78 bond releasing the amino-terminal PAP domain (Fig. 3). This results in the formation of the two-chained plasmin molecule associated by two disulphide bridges. A modified form of plasminogen, mini-plasminogen, created by limited proteolysis of plasminogen by elastase has been described, consisting of kringle 5 and the serine proteinase domain (Sottrup-Jensen et al., 1978). Activation of bovine plasminogen by uPA or tPA results in a cleavage at the activation site of the Arg557-Ile558 bond as well as in autocatalytic cleavage of the bond Lys77-Arg78, releasing the PAP fragment. Surprisingly, a further hydrolysis occurs in the bovine system, as the Arg342-Met343 bond is susceptible to cleavage by plasmin, resulting in a truncated form containing kringle 4, kringle 5 and the serine proteinase part (Benfeldt, 1992). This form has been called midiplasmin (Christensen et al., 1995).
Casein in milk: The main protein in milk is casein, of which there are 4 types in bovine milk, arranged as large colloidal particles commonly known as micelles. High resolution field-emission scanning electron microscope micrographs of casein micelles revealed spherical particles in the range of 200 nm; the surface of the micelle is not smooth but contains gaps between the tubular substructures (Dalgleish et al., 2004). Casein hydrolyzates exhibits myriad bioactivities such as immunoregulation and antimicrobial and anticarcinogenic actions, which support aspects of infant development, growth and survival beyond that, provided by nutrition alone. O-phospho-L-serine residues in CNH are essential for excreting its biological activity.
4 types of caseins are αS1-Casein, αS2-Casein, β-casein and κ-casein. β-CN is the preferred substrate for plasmin and its hydrolysis results in the production of γ-caseins and proteose-peptones (Andrews, 1983). The specificity of plasmin on β-casein is well documented. As well as producing y1-(B-CNf29-209), γ2- (B-CNfl06- 209), γ3- (B-CNfl08-209) caseins and their complementary N-terminal boiling-resistant peptides (proteose-peptone), plasmin can also cleave bonds Lys, 13-Tyr,'4 and Arg'83-AsP'84 in β-casein fairly rapidly. The αS1-casein (McSweeney et al., 1993) and αS2-casein (Le Bars and Gripon, 1989) are also susceptible to proteolysis by plasmin and the l-caseins are products of hydrolysis of αS1-casein. However, κ-casein is resistant to proteolysis by plasmin (Diaz et al., 1996).
β-casein and down regulation of milk: There is evidence for the existence of a close association between the PA-PG-PL system and gradual involution of mammary gland (the decline phase of lactation). Relative amounts of γ-caseins are highly negatively correlated with milk yield in the declining phase of lactation, reflecting the gradual involution of the gland at this time (Brown et al., 1995). The counter-part of γ-casein is the N-terminal fraction β-CN f (1-28) (Andrews, 1983) and therefore this peptide should also be highly negatively correlated with milk yield in the declining phase of lactation. Thus, the concept of Brown et al. (1995) support that β-CN f (1-28) plays a role in the regulation of milk secretion (Silanikove et al., 2000). β-CN f (1-28) is resistant to further degradation by plasmin (Andrews, 1983). No genetic substitutions in this part of β-CN have been found, so β-CN f (1-28) is an ideal candidate for negative feedback control of milk secretion.
MECHANISM OF ACTION
β-casein: The K+ channel blocker: Milk volume is determined by osmotic-coupled water flow. In bovines, the secretion of K+, Na+ and Cl- determines approximately 40% of the driving force, with the rest being determined by lactose (Shennan and Peaker, 2000). Lactose and monovalent ions are secreted into the lumen of the mammary gland mainly via vesicles. However, a direct contact between monovalent ions inside the epithelial cells and fluid stored in the lumen of the gland is possible, since the apical membrane of the epithelial cells contains K+, Na+ and Cl‾ channels (Shennan and Peaker, 2000). Indeed, it has been proved that a distinct plasmin-induced β-CN peptide f (1-28), defined as PPCB, is a potent blocker of K+ channels in the apical membrane of mammary epithelial cells (Silanikove et al., 2000).
The involvement of apically located K+-channel in the mechanism of feedback regulation of milk secretion is inspired by the need for extensive apical K+ conductance, since K+-concentration of milk is considerably higher than that of plasma. Indeed, K+ channels have been localized in the apical aspect of mammary secretory epithelia and have been shown to be associated in milk-born negative feedback regulation of milk secretion (Silanikove et al., 2000). The voltage-depended nature of the apical K+-channel, suggests that it belongs to the family of the voltage-gated potassium channel, KCNQ1. Assembly of KCNQ1 with the accessory units may shift the K+ channel to an open state, associated with hyperpolarization of the membrane potential, which in general facilitates secretary phenomena (VanTol et al., 2007). Thus, the milk-born regulatory element i.e., β-CN f (1-28), which blocks apically located K+-channel (Silanikove et al., 2000), cause depolarization of the membrane potential and in turn to down regulate cells metabolism and secretory activity.
β-casein: Ca2+ chelator: The concentration of plasmin is high during late lactation and increased with infrequent milking and interruption of milk removal. Simultaneously, during extended milk stasis, or mastitis, casein may undergo extensive degradation. It should be emphasized that the down-regulation of milk secretion is associated with mild activation of plasmin (10-40% increase in activity). Under such conditions, the response induced by β-CN f (1-28) affects specifically fluid secretion (water, lactose and ions), without affecting lactose concentration or fat and protein secretion and without affecting the integrity of tight junctions and the leukocyte level in the gland. Consequently, mild activation of the plasmin system results in reduced milk yield but with higher concentrations of fat and proteins, as seen in the response to injecting cows with dexamethasone (Shamay et al., 2000). On the other hand, massive activation of the plasmin activity (X150%) is associated with extensive degradation of casein, an inflammatory response (increase in leukocytes and NAGase activity), reduction in lactose concentration and disruption of tight junctions. As a result, fluid, fat and protein secretion are negatively affected, as seen in the response to treatment with CNH of goats (Shamay et al., 2002) and cows (Shamay et al., 2003).
Tight junction: The tight junction or zonula occludens, a ‘gasket-like’ structure, is part of the junctional complex and surrounds endothelial or epithelial cells in close proximity to the cell’s apical domain. When intact, it prevents the vectorial movement of small molecules and ions between the basolateral and apical sides of cells. This ‘barrier’ function of the tight junction permits the existence of pools of ions and/or small molecules differing in concentration on either side of the cell. In addition, tight junctions are instrumental in maintaining the polarised state of secretory cells, by preventing redistribution of plasma membrane components (e.g., ion channels and ion pumps) and maintaining a difference in lipid and protein composition between the basal and apical side of the plasma membrane. The latter function of the tight junction is often referred to as its ‘fence’ function (Mandel et al., 1993).
In the mammary gland, epithelial tight junctions, when ‘tight’, form a barrier between the blood, or more precisely the interstitial fluid (basolateral side) and milk in the alveolar lumen (apical side), thus preventing serum components from entering into milk and vice versa. In the periparturient period and again as the mammary gland involutes, towards the end of lactation, tight junctions are ‘leaky’. However, it was shown recently that the permeability of tight junctions can increase during established lactation if milk is allowed to accumulate. In cows, tight junctions switch to a leaky state after approximately 17 h of milk accumulation and in goats after 21 h (Stelwagen et al., 1997). Furthermore, this increase in permeability is associated with a reduction in milk secretion and an increase in protease activity in milk (Stelwagen et al., 1994).
Milk stasis causes the accumulation of local signals, which makes the TJ leaky. Stress activates the PPS system, leading to the release from b-CN (fraction 1-28) of multi phosphorylated peptide. Casein-ophospho-peptides, through their phospho-serine residues, may bind 20-40 M of Ca2+ (FitzGerald, 1998). Maintenance of extracellular Ca2+ levels is essential for maintaining the TJ integrity of the mammary secretory epithelium. During lactation, mammary epithelial cells extract large quantities of ionized calcium from plasma and produce a calcium rich secretion. In doing so, the mammary gland generates a large trans epithelial calcium gradient in favour of milk. Indeed, the concentration of calcium in milk can exceed 60 mM (Neville, 2005). Milk calcium exists in a variety of forms including, calcium bound to casein, colloidal calcium phosphate/citrate and free ionized calcium. The majority of calcium is associated with casein micelles allowing milk to maintain calcium at levels which, if fully ionized, would put mammary secretory cells under considerable osmotic stress. Casein micelle formation requires the presence of significant amounts of calcium (Holt, 2004). The association between calcium and casein appears to occur intra cellularly as it is apparent that secretory vesicles contain electron dense material, which is believed to be casein micelles.
During lactation, when the ducts and alveoli are filled with milk, the secretory epithelium is positioned between two very different environments; the milk, containing high concentration of lactose and low concentrations of sodium and chloride and interstitial fluid containing low concentration of lactose and high concentrations of sodium and chloride. Thus, when TJs are disrupted the Na+ concentration in the milk rises, whereas that of K+ declines. The stretching associated with milk stasis has been implicated in a mechano-transduction signaling pathway, which in turn can alter both milk synthesis and TJ permeability (Nguyen and Neville, 1988). The disruptions of blood vessels and of alveolus integrity are typical events in either inflammation or involution and account for the influx of lymphocytes and phagocytes into the alveolar lumen. During the onset of involution, the activity increased by as much as 500% (Athie et al., 1997), compared with an increase of 30% measured, when cows were exposed to stress (Silanikove et al., 2000). Thus, the precipitous dry-up of milk secretion may be related either to necrosis caused by the inflammation or to induction of involution.
PA-PG-PL system and hormonal interrelationship: Infusion of a solution composed of a casein digest enriched with β-CN f (1-28) into the cistern of cows or infusion of pure β-CN f (1-28) into the cistern of goats, led to a transient reduction in milk secretion in the treated gland (Silanikove et al., 2000). Stress and stress-related hormones such as glucocorticoids inhibit lactation in cows (Shamay et al., 2000) proposed a novel mechanism connecting stress with the PA-plasminogen-plasmin system. Stress activates the PA-plasminogen-plasmin system leading to an increase in plasmin activity and to the formation of β-CN f (1-28) (Silanikove et al., 2000).
The reduction in milk production due to dehydration stress or glucocorticoid (dexamethsone) is correlated with the activity of plasmin and channel blocking activity in the milk of cows (Silanikove et al., 2000). Complexes of both types of PA with PA inhibitor-1 have been detected in the culture medium of bovine mammary epithelial cells (Heegard et al., 1994).
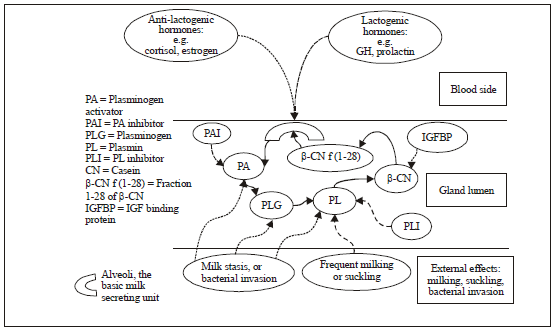 | |
| Fig. 4: | Overview of the PA-plasminogen-plasmin negative feedback mechanism that down-regulate milk secretion. (The contribution of most elements is described in the text. Bold arrows indicate flow signal along the feedback loop, dotted arrows positive effects and dashed arrows suppressive effects) |
Activation of the hypothalamus-pituitary-adrenocortical axis by external stress liberates cortisol into blood plasma, which in turn induces the liberation of PA from the mammary epithelial cells into the mammary cistern, where it activates the plasmin system and enhances the release of β-CN f (1-28) from β-CN. Inhibition of ion channels by β-CN f (1-28) triggers an as yet unknown process, which reduces the secretion of lactose and monovalent ions into the lumen of the gland, leading to the decrease in milk volume. This rapid modulation of milk secretion increases the potential for survival in response to stress (Fig. 4). Regulation of PA activity serves as a bridge between systemic hormonal influences and the local regulatory system (Silanikove et al., 2000). In the organ culture of the mouse mammary gland, hormones that fostered involution induced PA synthesis, whereas hormones that promoted lactation repressed PA secretion (Ossowski et al., 1979). The treatments of cows and sheep (Baldi et al., 1997) with bovine somatotrophin increased milk yield and reduced the conversion of PLG to PL. On the other hand, stress, stress hormones and estrogen, which induced down-regulation of milk yield, increased PL activity in milk (Athie et al., 1997; Silanikove et al., 2000).
Based on the evidence described above, this system may explain also the effects of frequency and efficiency of udder emptying and the response in levels of milk secretion to the presence of bacterial infection of the udder. A direct association between decreased milking frequency and an increase in the activity of the PA-plasminogen-plasmin system has been found (Stelwagen et al., 1994; Kelly et al., 1998).
Therapeutic role casein: Treatment with Casein Hydrolyzate (CNH), which contains active casein-derived peptides, have been shown to be very effective in eradicating bacterial infection and curing infected single udders of lactating cows from both subclinical and chronic infections. It has been shown also that within 8 h after the first application of CNH treatment induced marked bactericidal and bacteriostatic responses in the mammary secretion, against the main udder pathogens. This is resulted from the rapid and forceful activation of the innate immune system and the drastic reduction in nutrient availability for bacterial growth. Infusion of CNH dramatically accelerates the involution, which is completed within 3 days and results in a marked reduction in milk yield even after the first day of treatment. The CNH treatment at dry off prevents build up of udder pressure and is clearly associated with signs (lying behaviour and step numbers) that the treatment prevents the suffering associated with drying-off of high-yielding modern dairy cows.
Based on these findings, a casein hydrolyzate preparation is developed to reduce the suffering from mammary gland engorgement associated with abrupt cessation of milking (the conventional procedure to induce involution in modern dairy cows) and to treat and prevent common clinical and subclinical infections of the udder in dairy cows. This strongly highlights the potential of this method to serve as a viable non-antibiotic dry cow therapy treatment.
CONCLUSION
The success of future dairy sector depends on milk production of superior quality under harsh environmental condition which needs comprehensive knowledge about the role PA-PG-PL system in milk secretion. Hence, study of this system has become a key research priority and challenge for dairy scientists. Further investigations about the system will not only be helpful for more and sustainable milk production, but also provide knowledge for practical implication of the system in dry cow management and in reducing animal suffering.
REFERENCES
- Andrews, A.T., 1983. Proteinases in normal bovine milk and their action on caseins. J. Dairy Res., 50: 45-55.
CrossRefDirect Link - Athie, F., K.C. Bachman, H.H. Head, M.J. Hayen and C.J. Wilcox, 1997. Milk plasmin during bovine mammary involution that has been accelerated by estrogen. J. Dairy Sci., 80: 1561-1568.
CrossRefDirect Link - Baldi, A., V. Chiofalo, G. Savoini, R. Greco, F. Polidori and I. Politis, 1997. Changes in plasmin, plasminogen and plasminogen activator activities in milk of late lactating ewes: Effects of bovine somatotropin (BST) treatment. Livestock Prod. Sci., 50: 43-44.
CrossRefDirect Link - Barash, H., N. Silanikove, A. Shamay and E. Ezra, 2001. Interrelationships among ambient temperature, day length and milk yield in dairy cows under a mediterranean climate. J. Dairy Sci., 84: 2314-2320.
Direct Link - Bastian, E.D. and R.J. Brown, 1996. Plasmin in milk and dairy products: An update. Int. Dairy J., 6: 435-457.
CrossRefDirect Link - Brown, J.R., A.J. Law and C.H. Knight, 1995. Changes in casein composition of goats' milk during the course of lactation: Physiological inferences and technological implications. J. Dairy Res., 62: 431-439.
CrossRefDirect Link - Christensen, S., L. Sottrup-Jensen and U. Christensen, 1995. Stopped-flow fluorescence kinetics of bovine alpha 2-antiplasmin inhibition of bovine midiplasmin. Biochem. J., 305: 97-102.
Direct Link - Clare, D.A. and H.E. Swaisgood, 2000. Bioactive milk peptides: A prospectus. J. Dairy Sci., 83: 1187-1195.
CrossRefPubMedDirect Link - Collier, R.J., J.L. Collier, R.P. Rhoads and L.H. Baumgard, 2008. Invited review: Genes involved in the bovine heat stress response. J. Dairy Sci., 91: 445-454.
CrossRefDirect Link - Collier, R.J., G.E. Dahl and M.J. Van Baale, 2006. Major advances associated with environmental effects on dairy cattle. J. Dairy Sci., 89: 1244-1253.
CrossRefDirect Link - Crimp, S.J., C.J. Stokes, S.M. Howden, A.D. Moore and B. Jacobs et al., 2010. Managing Murray-darling basin livestock systems in a variable and changing climate: Challenges and opportunities. Rangeland J., 32: 293-304.
CrossRefDirect Link - Dalgleish, D.G., P.A. Spagnuolo and H.D. Goff, 2004. A possible structure of the casein micelle based on high-resolution field-emission scanning electron microscopy. Int. Dairy J., 14: 1025-1031.
CrossRefDirect Link - Daly, S.E., R.A. Owens and P.E. Hartmann, 1993. The short‐term synthesis and infant‐regulated removal of milk in lactating women. Exp. Physiol., 78: 209-220.
CrossRefDirect Link - Diaz, O., A.M. Gouldsworthy and J. Leaver, 1996. Identification of peptides released from casein micelles by limited trypsinolysis. J. Agric. Food Chem., 44: 2517-2522.
CrossRefDirect Link - FitzGerald, R.J., 1998. Potential uses of caseinophosphopeptides. Int. Dairy J., 8: 451-457.
CrossRefDirect Link - Heegard, C.E., J.H. White, B. Zavizion, J.D. Turner and I. Politis, 1994. Production of various forms of plasminogen activator and plasminogen activator inhibitor by cultured mammary Eplthelial cells. J. Dairy Sci., 77: 2949-2958.
CrossRefDirect Link - Heegaard, C.W., L.K. Rasmussen and P.A. Andreasen, 1994. The plasminogen activation system in bovine milk: Differential localization of tissue-type plasminogen activator and urokinase in milk fractions is caused by binding to casein and urokinase receptor. Biochimica et Biophysica Acta (BBA)-Mol. Cell Res., 1222: 45-55.
CrossRefDirect Link - Holt, C., 2004. An equilibrium thermodynamic model of the sequestration of calcium phosphate by casein micelles and its application to the calculation of the partition of salts in milk. Eur. Biophys. J., 33: 421-434.
CrossRefDirect Link - Horowitz, M., 2001. Heat acclimation: Phenotypic plasticity and cues to the underlying molecular mechanisms. J. Thermal Biol., 26: 357-363.
CrossRefDirect Link - Kadzere, C.T., M.R. Murphy, N. Silanikove and E. Maltz, 2002. Heat stress in lactating dairy cows: A review. Livestock Prod. Sci., 77: 59-91.
CrossRefDirect Link - Kelly, A.L., S. Reid, P. Joyce, W.J. Meaney and J. Foley, 1998. Effect of decreased milking frequency of cows in late lactation on milk somatic cell count, polymorphonuclear leucocyte numbers, composition and proteolytic activity. J. Dairy Res., 65: 365-373.
Direct Link - Le Bars, D. and J.C. Gripon, 1989. Specificity of plasmin towards bovine alphaS2-casein. J. Dairy Res., 56: 817-821.
PubMedDirect Link - Leitner, G., M. Chaffer, A. Shamay, F. Shapiro, U. Merin, E. Ezra, A. Saran and N. Silanikove, 2004. Changes in milk composition as affected by subclinical mastitis in sheep. J. Dairy Sci., 87: 46-52.
Direct Link - Maltz, E., O. Kroll, H. Barash, A. Shamy and N. Silanikove, 2000. Lactation and body weight of dairy cows: Interrelationships among heat stress, calving season and milk yield. J. Anim. Feed Sci., 9: 33-46.
Direct Link - Maltz, E. and N. Silanikove, 1996. Kidney function and nitrogen balance of high yielding dairy cows ate the onset of lactation. J. Dairy Sci., 79: 1621-1626.
CrossRefPubMedDirect Link - Mandel, L.J., R. Bacallao and G. Zampighi, 1993. Uncoupling of the molecular fence and paracellular gate functions in epithelial tight junctions. Nature, 361: 552-555.
CrossRefDirect Link - McSweeney, P.L.H., N.F. Olson, P.F. Fox, A. Healy and P. Hojrup, 1993. Proteolytic specificity of plasmin on bovine αs1‐Casein. Food Biotechnol., 7: 143-158.
CrossRefDirect Link - Neville, M.C., 2005. Calcium secretion into milk. J. Mammary Gland Biol. Neoplasia, 10: 119-128.
CrossRefDirect Link - Nguyen, D.A.D. and M.C. Neville, 1998. Tight junction regulation in the mammary gland. J. Mammary Gland Biol. Neoplasia, 3: 233-246.
CrossRefDirect Link - Ossowski, L., D. Biegel and E. Reich, 1979. Mammary plasminogen activator: Correlation with involution, hormonal modulation and comparison between normal and neoplastic tissue. Cell, 16: 929-940.
CrossRefDirect Link - St-Pierre, N.R., B. Cobanov and G. Schnitkey, 2003. Economic losses from heat stress by US livestock industries. J. Dairy Sci., 86: E52-E77.
CrossRefDirect Link - Politis, I., A. Voudouri, I. Bizelis and G. Zervas, 2003. The effect of various vitamin E derivatives on the urokinase-plasminogen activator system of ovine macrophages and neutrophils. Br. J. Nutr., 89: 259-265.
CrossRefDirect Link - Politis, I., B. Zavizion, F. Cheli and A. Baldi, 2002. Expression of urokinase plasminogen activator receptor in resting and activated bovine neutrophils. J. Dairy Res., 69: 195-204.
CrossRefDirect Link - Saksela, O. and D.B. Rifkin, 1988. Cell-associated plasminogen activation: Regulation and physiological functions. Ann. Rev. Cell Biol., 4: 93-120.
CrossRefDirect Link - Shamay, A., F. Shapiro, G. Leitner and N. Silanikove, 2003. Infusions of casein hydrolyzates into the mammary gland disrupt tight junction integrity and induce involution in cows. J. Dairy Sci., 86: 1250-1258.
CrossRefDirect Link - Shamay, A., F. Shapiro, S.J. Mabjeesh and N. Silanikove, 2002. Casein-derived phosphopeptides disrupt tight junction integrity and precipitously dry up milk secretion in goats. Life Sci., 70: 2707-2719.
CrossRefDirect Link - Shennan, D.B. and M. Peaker, 2000. Transport of milk constituents by the mammary gland. Physiol. Rev., 80: 925-951.
Direct Link - Sidelmann, J.J., J. Gram, J. Jespersen and C. Kluft, 2000. Fibrin clot formation and lysis: Basic mechanisms. Semin Thromb Hemost., 26: 605-618.
PubMedDirect Link - Silanikove, N., A. Shamay, D. Shinder and A. Moran, 2000. Stress down-regulates milk yield in cows by plasmin induced beta-casein product that blocks K+ channels on the apical membranes. Life Sci., 67: 2201-2212.
CrossRefDirect Link - Silanikove, N., 2000. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livestock Prod. Sci., 67: 1-18.
CrossRefDirect Link - Silanikove, N., 1994. The struggle to maintain hydration and osmoregulation in animals experiencing severe dehydration and rapid rehydration: The story of ruminants. Exp. Physiol., 79: 281-300.
CrossRefDirect Link - Sottrup-Jensen, L., H. Claeys, M. Zajdel, T.E. Petersen and S. Magnusson, 1978. The Primary Structure of Human Plasminogen: Isolation of the two Lysine-Binding Fragments and one Mini-Plasminogen (MW. 38000) by Elastase Catalysed, Specific Limited Proteolysis. In: Progress in Chemical Fibrinolysis and Thrombolysis, Davidson, J.F., R.M. Rowan, M.M. Samama and P.C. Desnoyers (Eds.). Vol. 3, Raven Press, New York, pp: 191-209.
- Stelwagen, K., V.C. Farr, H.A. McFadden, C.G. Prosser and S.R. Davis, 1997. Time course of milk accumulation-induced opening of mammary tight junctions and blood clearance of milk components. Am. J. Physiol. Regulatory Integrative Comp. Physiol., 273: R379-R386.
Direct Link - Stelwagen, K., C.G. Prosser, S.R. Davis, V.C. Farr, I. Politis, J.H. White and B. Zavizion, 1994. Effect of milking frequency and somatotropin on the activity of plasminogen activator, plasminogen, and plasmin in bovine milk. J. Dairy Sci., 77: 3577-3583.
CrossRefDirect Link - Ueshima, S., K. Okada and O. Matsuo, 1996. Stabilization of plasmin by lysine derivatives. Clin. Chimica Acta, 245: 7-18.
CrossRefDirect Link - VanTol, B.L., S. Missan, J. Crack, S. Moser, W.H. Baldridge, P. Linsdell and E.A. Cowley, 2007. Contribution of KCNQ1 to the regulatory volume decrease in the human mammary epithelial cell line MCF-7. Am. J. Physiol. Cell Physiol., 293: C1010-C1019.
CrossRefDirect Link - West, J.W., 2003. Effects of heat-stress on production in dairy cattle. J. Dairy Sci., 86: 2131-2144.
CrossRefPubMedDirect Link - Wilde, C.J. and M. Peaker, 1990. Autocrine control in milk secretion. J. Agric. Sci., 114: 235-238.
CrossRefDirect Link