
I.A.A. Abou Ayana
Department of Dairy Research, Food Technology Research Institute, Agricultural Research Center, Giza, Egypt
A.A. Gamal El-Deen
Department of Dairy Research, Food Technology Research Institute, Agricultural Research Center, Giza, Egypt
K.M.K. Ayyad
Department of Dairy Technology Research, Animal Production Research Institute, Agricultural Research Center, Giza, Egypt
M.A. El-Metwally
Department of Mycology Research and Plant Diseases Survay, Plant Pathology Research Institute, Agricultural Research Center, Giza, Egypt
International Journal of Dairy Science
Year: 2014 | Volume: 9 | Issue: 2 | Page No.: 32-44







ABSTRACT
The fungi play an important role in cheese manufacture such as Penicillium roqueforti nevertheless subsequent development of filamentous fungi occurs during cheese ripening on its surface, this accompanied with production of some bad flavors and tastes, as well as some mycotoxins. In this study, 125 samples of cheeses were collected to determination of fungi and aflatoxin B1 (AFB1). The results indicated that percentages of Aspergillus flavus, Aspergillus niger, Aspergillus ocnrecious and Aspergillus oryzae were 18.6, 12.4, 8.6 and 5.3%, respectively. Rhizopus spp., Penicillium spp., Mucor spp., trichoderma spp. and Fusarium moniliforme recorded 22.1, 16.8, 11.5, 2.4 and 0.5%, respectively. Aged Ras cheese was the highest contamination with AFB1 (from 8.3a to 19.5a ppb) average 13.9a ppb, fresh Domiate cheese recorded the lowest contamination with toxin with average 0.95f ppb. A. flavus produced the highest quantity of AFB1 in broth medium for 2 months unlike Penicillium spp. was the lowest Production of toxin (0.6c ppb). Counts of fungi and AFB1 production affected by tested anti-fungi, Ascorbic acid and Broken Saccharomyces cerevisiae Cells after Freezing (BSCAF) were stronger set but Salicylic acid and Citric acid were the weaker set anti-fungi in this point. Aspergillus flavus was the greatest resistant to Salicylic acid.
PDF Abstract XML References Citation
Received: November 25, 2013;
Accepted: January 20, 2014;
Published: May 06, 2019
How to cite this article
I.A.A. Abou Ayana, A.A. Gamal El-Deen, K.M.K. Ayyad and M.A. El-Metwally, 2014. Influence of some Abiotic Anti-Fungi and Saccharomyces cerevisiae on Aflatoxin B1 and Isolated Fungi from some Cheese Types. International Journal of Dairy Science, 9: 32-44.
DOI: 10.3923/ijds.2014.32.44
URL: https://scialert.net/abstract/?doi=ijds.2014.32.44
DOI: 10.3923/ijds.2014.32.44
URL: https://scialert.net/abstract/?doi=ijds.2014.32.44
INTRODUCTION
Likely that the cheese industry began nearly 8,000 years ago (Fox et al., 2000), quickly spread around the world, which led to the great diversity in the types of cheese, for the time being is estimated at more than 1000 (Irlinger and Mounier, 2011). Cheese is an important industry, in 2009, for example, France produced 1692369, 93764 and 57328 tons of cow, goat and sheep cheeses, respectively (Agreste, 2009). In cheese production, molds, yeasts and bacteria are inoculated into the milk and that play a major role in obtaining the desired flavor, aroma, texture, color, aspect and nutritive elements of the cheese so must know-how to control the cultivation of them (Beresford et al., 2001; Irlinger and Mounier, 2011). Acidification of milk with lactic acid bacteria is necessary to the next evolution of filamentous fungi occurs during cheese ripening, fungi usually growth only on the surface of the most cheeses. Excepting blue cheeses, the inoculated fungus develops inside the blue “veins” of the cheese. Over and above, indigenous molds of the ripening rooms colonize the surface of cheeses. These unwanted organisms microorganisms are usually connected with undesirable effects such a negative impact on flavor and taste, discoloration that is unacceptable to most consumers or, rarely, production of toxic secondary compounds (Lund et al., 1995, 2003). Under laboratory conditions, most Penicillium species produce mycotoxins even among inoculated species such as Penicillium roqueforti and Penicillium camemberti (Frisvad et al., 2004). Even though no direct evidence of toxicity to humans has been reported for cyclopiazonic acid produced by P. camemberti but absence of toxin in cheeses cannot be taken for granted (Pitt, 2002). P. roqueforti produces many toxic secondary metabolites, which may not pose a threat to public health, even with the daily consumption of blue-veined cheeses (Teubler and Engels, 1983), P. roqueforti toxin has been shown to be unstable in cheese (Chang et al., 1993).
Aspergillus flavus and Aspergillus parasiticus are the most famous fungi have the ability to produce toxic by-products called Aflatoxins (Harper, 2003). AFB1 is the most potent mycotoxin and has been shown to be a potent carcinogen. According to the International Agency Research on Cancer (IARC), AFB1 and AFM1 are categorized as class 1A and class 2B carcinogens, respectively (IARC, 1993; Maurice, 2002). There was a linear relationship between the quantity of AFM1 in milk and AFB1 in feed consumed by animals (Bakirci, 2001; Dragacci et al., 1995). AFB1 which originating feed is metabolized and excreted as AFM1 in the urine and thus in milk (Bakirci, 2001; Gurbay et al., 2006; Lopez et al., 2003). Without paying attention to that, exposure to aflatoxin can be through ingestion of contaminated milk containing AFM1.
The European Commission identified the limits for maximum levels of total aflatoxins (4 μg kg-1) and AFB1 (2 μg kg-1) allowed in groundnuts, nuts, dried fruit and their products. For nuts and dried fruit to be subjected to sorting or other physical treatment, before human consumption or use as an ingredient in foodstuffs the limits stand at 10 μg kg-1 (total aflatoxins) and 5 μg kg-1 (AFB1), 0.05 μg L-1 of AFM1 in milk, butter and cheese (Anonymous, 2001; EC, 2002).
Many microorganisms such as bacteria, yeasts and filamentous fungi play a vital role in the development of organoleptic characteristics and/or, on the contrary, may be the source of quality defects (Florez and Mayo, 2006; Callon et al., 2006; Gente et al., 2007).
Salicylic acid is used as a food preservative, as bactericidal and an antiseptic although its large quantities are toxic. For some people with Salicylate sensitivity even these small doses can be harmful. Ascorbate usually acts as an antioxidant. It typically reacts with oxidants of the reactive oxygen species, such as the hydroxyl radical formed from hydrogen peroxide. Such radicals are damaging to animals and plants at the molecular level due to their possible interaction with nucleic acids, proteins and lipids. Sometimes these radicals initiate chain reactions. Ascorbate can terminate these chain radical reactions by electron transfer (Masram and Jugade, 2013).
The goal of this study is determination of filamentous fungi on some types of cheese and their production of AFB1 to determine the severity of the growth of these organisms on cheese and how to control their growth as well as the production of the toxin on cheese by using Salicylic, Ascorbic and Citric acids and BSCAF.
MATERIALS AND METHODS
Sample collection: One hundred and twenty five of different cheese samples (local or imported, in Table 1) were collected from major department stores, supermarkets, stores and some small factories in Dakahlia Governorate-Egypt, during the months of May to September 2012.
| Table 1: | Types and No. of cheese samples |
| AD: Aged domiate, AR: Aged ras, B: Brick, C: Cheddar, E: Edam, F: Feta, FD: Fresh domiate, FR: Fresh ras, G: Gouda, P: Processed | |
All samples were collected under aseptic precautions and transferred to the laboratory as soon as possible to examine microbiologically for the presence of fungi and aflatoxins.
Aflatoxin B1 (AFB1) was obtained from Sigma (Deisenhofen-Germany). AFB1 was desolved in benzine: acetonitrle (98:2) to give a solution of final concentration containing 0.1 μg mL-1.
Salicylic, Ascorbic and Citric acids were obtained from CID Pharmaceuticals CO. Salicylic acid was dissolved in boiled water to get concentrations (1, 3 and 6 mmol) , Ascorbic and Citric acids were diluted in distilled water to reach the concentrations required (5,10 and 15 mmol).
Commercial bread yeast was used as a source for the isolation of Saccharomyces cerevisiae. The most appropriate medium for this purpose was the modified Saborou medium (Savova and Nikolova, 2002). The isolated strain was identified as S. cerevisiae according to the procedures described by Barnett et al. (1990) and in consultant with Department of Microbiology, Soils, Water and Environment Research Institute, ARC, Giza, Egypt.
Partial extraction of S. cerevisiae (Meyen ex E.C Hansen) was prepared by growing S. cerevisiae on plates of malt extract agar medium for 4 days at 28°C. The resulted cells were scraped gently with Pasteur pipet using 10 mL of sterilized water. The cell suspension was adjusted to 1011 cell Ml-1 and kept under freezing for 48 h to allow partial disruption of the cell walls. After melting, the obtained disrupted cells were considered as 100% concentration of yeast extract, abbreviated (BSCAF).
Ras Cheese was manufactured according to Abou-Donia (2002). Separately salicylic, Ascorbic and Citric acid S. cerevisiae at 50 and 100% concentrations were mixed with 5% Arabic gum and used to cover the surface of manufactured Ras cheese to study the controlling natural toxogenic fungi through 4 months of ripening period. During that period, the fungi which grew and AFB1 were determined after 30, 60, 90 and 120 days.
Isolation and identification of fungi: Czapek Doxs Agar medium (Oxoid, 1982) and potato dextrose Agar were used to isolate, purified and identify of fungi. The inoculated media with examined samples were incubated at 35°C for 5 days, then, all fungi isolates subjected to purification. Finally, isolates were identified in mycological laboratory-Botany department, faculty of Science, Mansoura University. Six isolates only were chosen for aflatoxin production through two months interval of incubation at room temperature.
Production of aflatoxins on synthetic media:
| • | Modified Czapek’s medium for aflatoxins production, Sucrose 200 g, NaNO3 2.0 g, KCl 0.5 g, K2HPO4 1.0 g, MgSO4 7H2O 0.5 g, Yeast extract 20 g, FeSO4. 7H2O 0.001 g, ZnSO4 0.001 g, MnCl2 0.001 g, distilled water 1000 mL |
| • | Surface growth on liquid medium: Fermented bottles (1 L) containing 100 mL modified czapek’s. Liquid medium were inoculated with spore suspensions of the isolated strains of fungi, 2 weeks old culture grown on agar medium and incubated at 30°C for 9 days |
| • | Extraction of aflatoxins from liquid synthetic medium culture were filtered and aflatoxins were extracted from filtrates by Refluxing for 1-2 h with chloroform. When high yields of toxin were expected, the filtrated media was shaken vigorously with 100 mL chloroform in reparatory funnels the lower chloroform layer were recovered and concentrated on a steam bath to just dryness |
Inoculum: The mold was grown on PDA slants for 14 days at room temperature (c.a. 25°C). Spores were harvested by adding 10 mL of sterile phosphate buffer (pH 7.0) to the slant and gently brushing the conidiophores with a sterile Inoculating loop.
Thin layer chromatography:
| • | Preparation of plates: Plates were prepared as mentioned in AOAC (1995) |
| • | Standard aflatoxin as above mentioned. A standard of AFB1 was spotted on TLC plate pre-coated with silica gel and activated; the plate was developed with solvent acetone: chloroform (1:9). When the solvent reached to 15 cm, then the plate was removed from the jar, air dried and identified by U.V. light and the Rf values were recorded for each of the six standards. Standard curve was measured at 365 nm by using thermo-Spectronic (Spectrophotometer) |
Column clean up chromatography: After evaporation of the remaining hexane by a slightly stream of nitrogen at 40°C the extracts were applied to strong anion exchange (C 18) Cartridge (100) mg 1 mL-1, phenomenex, Aschffenburg, Germany) which has been conditioned twice with 1 mL of aqua distilled water, following 1 mL of acetonitryle (ACN). One milliliter from extracted sample was transferred to clean up then, the cartridge was washed twice with 1 mL of aqua dist. water and 1 mL 10% ACN in aqua dist. water after that, column was washed and rinsed by 1 mL Aqua dist. water and dried by the use of vacuum for 2 min. Aflatoxin was eluted with 0.5 mL 30% ACN/aqua. dist. water for TLC.
Hard and soft cheeses, 15 g were blended. Cheese was cut in to small pieces, with 1 mL of salt solution, 5 g diatomaceous earth and 100 mL CHCl3 60 seconds in a 1 L blender Jar. The mixture was filtered through whatman (1) paper into 125 mL Erlenmeyer. Extract was treated as for fluid milk. Maximum filtrate was obtained from cheese extract by closing filter paper top and compressing entire paper and contents against funnel.
Antagonism of antioxidants against fungi: The disc system procedure was conducted to study that and Salicylic, Ascorbic and Citric acid and S. cerevisiae were used to study inhibition isolated fungi.
Salicylic, Ascorbic and Citric acid and S. cerevisiae were tested in vitro on the linear growth of the pathogenic fungi. Different concentrations were added to 10 mL of sterilized PDA before solidification and then poured in sterile Petri-dishes. After solidification, the plates were inoculated with fungal disc (5 mm) in the center of the plate and incubated at 27±1°C. Three plates for each particular treatment for each fungus were used as replicates; three plates were prepared to serve as control for each fungus. Linear growth was observed daily and diameter of fungal colonies were recorded when plates of any treatment were filled with the fungal growth.
RESULTS AND DISCUSSION
A hundred twenty five of different cheese samples were collected intentionally. All collected samples were examined for containing fungi. Our results evidenced a great phylogenetic diversity in fungi able to grow on different types of cheese and, in terms of ecology, a high specialization for cheese in some species but not in others that are generalist fungi. Having made an accurate species delimitation of cheese taxa, we were then able to propose molecular targets for their identification. Shown results in Table 2 appears the types of fungi that were found in collected cheese samples and the proportion of their presence in collected samples.
Bacteria, fungi and yeasts which have an important and necessary role during the ripening phase of cheese making can cohabit in the cheese because of cheese is a substrate of a unique heterogeneous physicochemical composition with high levels of protein and fat, as it matures, supports a complex ecosystem (Irlinger and Mounier, 2011), many products liberated by other microorganisms have benefits, must be exploited. Interaction among coexisting microbes may be through simple trophic or nutritional relationships or the microbes may interact directly through physical contact or via signaling molecules (Sieuwerts et al., 2008).
Five species of isolated fungi belonging to two genera (Aspergillus and fusarium) and other four genera (Penicillium spp., Rhizopus spp., Mucor spp. and Trichoderma spp.) Were detected in all examined samples, these fungal species are belonging to three classes Eurotiomycetes, Mucormycotina and Sordariomycetes (according to relative frequencies).
The Eurotiomycetes comprise five species related to two genera namely Aspergillus and Penicillium that include four species for Aspergillus and Penicillium spp. The class Sordariomycetes was represented by Trichoderma spp. and F. monliform. Mucormycotina also was represented by Mucor spp. and Rhizopus spp.
Aspergillus spp. recorded the greatest ratio among all the isolated fungi, Aspergillus spp. percents were 18.9, 13.4, 8.6 and 5.3% for A. flavas, A.niger, A. ocnrecious and A. oryzae, respectively. Rhizopus spp. came in the second group of fungi in percent 22.1% followed by Penicillium spp. 16.8%, Mucor spp. 11.5% and Trichoderma spp. 2.4%, whilst Fusarium moniliforme recorded the lowest percent 0.5%. These results fully agreed with Houbraken and Samson (2011) who reported that most Eurotiales includes many well-known and common fungi, particularly Penicillium and Aspergillus, recently grouped into the Aspergillaceae family. These fungi, which include Lipolytic and Proteolytic species El-Diasty and Salem (2007). Noteworthy observation, that 44.9% of examined samples were contaminated with toxic Aspergillus. This diversity of isolated fungi shows that many types of cheese are rich media for the growth of several fungi; that may be due to contamination of ripening rooms, which is the perfect environment for fungal growth or because of unhealthy trading of these cheeses. Surely, this corresponded with reaching low pH-values appropriate for enhancing the growth of these organisms, which naturally occur in air.
These findings agreed with Abdel All et al. (2008), they detected eight toxic Aspergillus strains (A. Parasiticus, A. Flavus, A. niger, A. versicolor, A. terrus. A. ochraceus, A. candidus and A. nidulans) and two of toxic Penicillium strains in cheese samples included Ras, White, Process, Tilsit and Cheddar cheese.
| Table 2: | Isolated fungi from cheese samples and their percentage |
| F.m: F. monliforme, Penici: Penicillium spp., Rhizop: Rhizopus spp., Tricho: Trichoderma spp. | |
Twenty-three strains were identified as P. camemberti, 22 as Penicillium biforme and 2 as Penicillium fuscoglaucum, two species considered as cheese contaminants. In this study, filamentous fungi ascomycetes in investigated cheese samples are members of two classes, the Eurotiomycetes and the Sordariomycetes. Within Eurotiomycetes, three genera are able to develop on cheeses, Penicillium within the Eurotiales and Sporendonema/Sphaerosporium within the Onygenales. Eurotiales includes many well-known and common fungi, particularly Penicillium and Aspergillus, recently grouped into the Aspergillaceae family (Houbraken and Samson, 2011). Most Eurotiales fungi are saprotrophic and represent some of the most catabolically and anabolically diverse microorganisms. They are among the most important food microorganisms (Geiser et al., 2006). Often fungi considered as food spoilers, however some are used as starter culture for cheese and meat products. P. camemberti and P. roqueforti are inoculated during cheese making process but closely related species of both are able to develop on this substrate as well but are considered as cheese contaminants (i.e., P. biforme, P. fuscoglaucum, P. palitans, P. solitum, P. nordicum and P. crustosumas relatives of P. camemberti; P. paneum as a close relative of P. roqueforti). (Frisvad and Samson, 2004).
Many fungi contaminate a vast array of food and agricultural commodities includes Aspergillus and Penicillium species which capable of growing on a variety of substrates and under a variety of environmental conditions as well as capable of producing aflatoxins Therefore, most foods are susceptible to aflatoxigenic fungi at some stage of production, processing, transportation and storage. In this investigation, AFB1 was determined in all cheese samples after collecting, data presented in Table 3 refer to that, two types of cheese (Gouda and Roquefort) were free of AFB1, most Penicillium species are known to produce mycotoxins under laboratory conditions even among inoculated species such as P. roqueforti and P. camemberti (Frisvad et al., 2004). Toxin of P. roqueforti showed instability in cheese (Chang et al., 1993). Although there is no evidence of toxicity to humans has been reported for cyclopiazonic acid produced by P. camemberti, absence of toxin in cheeses cannot be taken for granted (Pitt, 2002). P. camemberti produces several secondary metabolites that are toxic but it appears that they represent a low public health risk, even with the daily consumption of blue-veined cheeses (Teubler and Engels, 1983).
| Table 3: | Minimum and maximum of aflatoxin B1 content in different cheese samples, which were collected during the months of May to September 2012 |
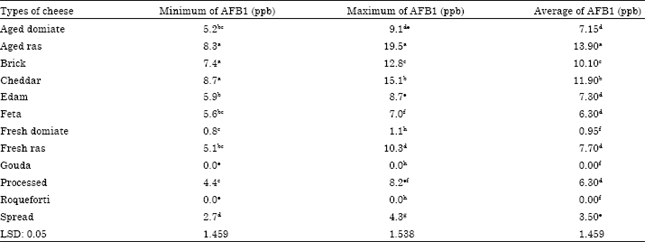 | |
On the contrary, other samples had different levels of toxin, statistical analysis LSD at (p>0.05) appeared, Aged Ras cheese was the highest contamination with AFB1 from 8.3a to 19.5a ppb average 13.9a ppb followed by Cheddar, Brick (fresh Ras and Edam and Aged Domiate and feta and Processed) and Spread. Fresh Domiate cheese recorded the lowest contaminate with toxin with average 0.95f ppb. These results were similar with findings by Abdel All et al. (2008) they found many cheese samples were contaminated with six types of Aflatoxins AFB1, AFB2, AFG1, AFG2, AFM1 and AFM2, ranged between 1.0 and 13 ppb, they also revealed that AFB1 was the highest level of toxin among other Aflatoxins, it ranged between 3.1 and 13 ppb. AFM1 was found in 16.6% of the cheeses tested, 31.3% of these were sheep-goat cheeses, 27.2% cow, 16.7% goat and 12.8% sheep cheeses, with no significant differences (p>0.05), all the samples of buffalo milk cheese were consistently negative (Montagna et al., 2008). Ashraf (2012) reported that AFM1was detected in several dairy products at different levels. It was noticeable that the level of contamination associated the age of cheese and the type or the method of industry, Roquefort cheese samples were free of AFB1 this probably due to the dominion of P. roqueforti that is used in the manufacture of that type of cheese.
Six isolated fungi were chosen to study production of aflatoxin B1 during incubation for two months in selective broth medium. Results in Table 4 refers to after 30 days under optimal conditions and medium, A. flavus produced the greatest quantity of AFB1 (9.2a ppb) followed by A. niger, A. oryzae, then Penicillium spp. Unlike A. ochraceus and Fusarium moniliforme failed completely to produce any quantity of AFB1 or maybe the quantity produced is not sensible. It is noted also the most quantities of AFB1were produced in the first thirty days whereas after 60 days of incubation in broth medium, the increase of AFB1 was insignificant, during the second month, A. flavus came at the first fungus among all tested fungi but Penicillium spp. failed to produce more than 0.6c ppb of AFB1.
To study the effect of Ascorbic acid, Citric acid, Salicylic acid or BSCAF on isolated fungi from cheese samples, a laboratory experimental was designed. Sterilized selective medium was plated into sterile dishes, ten dishes for each fungus and antifungal substance were used, one of them as a control and three for concentration.
Generally, data presented in Table 5 illustrated that all concentrations of additives appeared a significant antagonism effect towered all isolated fungi, BSCAF had the greatest effect on tested fungi followed by Ascorbic acid, Salicylic acid then citric acid. The most important observation in these results that second concentration of each antifungal substances is the best to inhibit tested fungi. Inhibition of fungi by using biotic or abiotic agents is a promising method; Galal et al. (1997) reported that the possibility of obtaining Surface-fungal growth control with antioxidants.
Most of the tested antioxidants revealed toxic effect to Fusarium spp., as well as other genera. These results suggested that the sensitivity of the some fungi to antioxidants was different from one species to another.
| Table 4: | Effect of different isolated fungi on aflatoxin B1 production after 30 or 60 days of incubation in specific broth media |
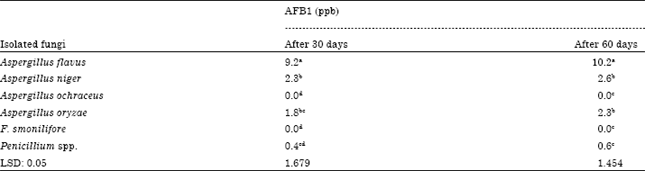 | |
| Table 5: | The antagonism effect of some additives on isolated fungi from several cheese samples |
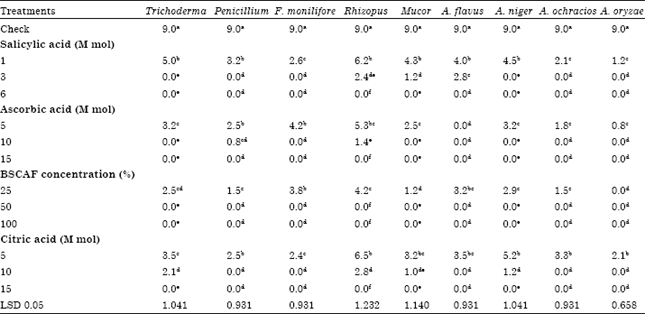 | |
However, the inhibitory effect of antioxidants increased with increasing the concentration. It was also reported that not only the mycelial growth of the fungi was influenced by antioxidants but also the percentage of sporulation was sharply decreased. Similar conclusion was reported on Fusarium spp. (Galal and Abdou, 1996). Elwakil and El-Metwally (2000) found that, the most potent antioxidant on the linear growth of F. moniliforme, was hydroquinone. Citric acid reduced the linear growth of some fungi however; salicylic acid inhibited its growth at 15 mM. The full inhibition of F. moniliforme, was achieved by all used antioxidants at 10-15 mM (Yousef, 2001).
Giridhar and Reddy (2001) explained the antioxidant activity based on reduction of oxygen tension in media that subsequently alter the membrane permeability and lead to growth and mycotoxin inhibition. The partial effect of antioxidants was improved by the addition of compounds used in the food industry to prevent fatty acid oxidation and deterioration of products (Sims and Fioriti, 1978). Some antioxidants are food grade. Therefore, they are safe for use from an antifungal point of view.
Curcumin may be employed successfully to control of toxigenic fungal growth on food, feed and subsequent contamination as well aflatoxins (Jahanshiri et al., 2012).
On the other hand, A. flavus was higher resistant than other fungi at 3 m mol of Salicylic acid followed by Rhizopus spp. then Mucor spp. Rhizopus spp. was the highest resistant at 10 m mL of Ascorbic acid then Penicillium spp. which recorded growth 0.8 cm around the disk. Lastly, citric acid failed to inhibit completely four fungi, Rhizopus spp., Trichoderma spp., A. niger and Mucor spp. at 10 M mL concentration. The yeast exhibited inhibit completely towards all tested fungi at 50 or 100% concentration this may be due to the availability of antioxidants in it.
Another important note, all isolated species of Aspergillus spp. were inhibited completely by second concentration of the four antifungal substances except A. flavus and A. niger in the presence of Salicylic acid and Citric acid, respectively. In addition, F. moniliforme was inhibited completely by the second concentration of all antifungal substances. The third concentration of all antifungal substances inhibited completely all tested isolates.
| Table 6: | Incidence of some fungi in ras cheese after treating superficially with some additives during storage for 120 days |
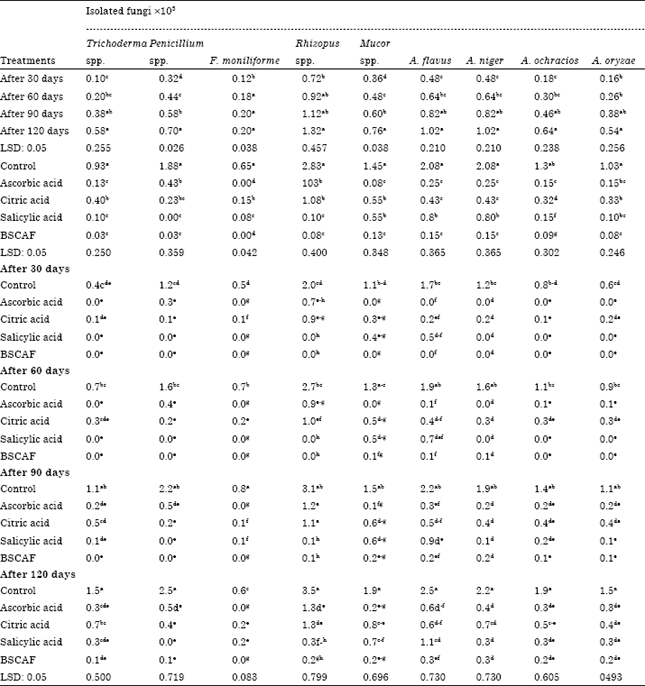 | |
From previous results, it was found that Ras cheese was the highest contaminated with AFB1. Therefore, Ras cheese was manufactured and treated with mentioned antifungal and stored in ripening room, then fungi and AFB1 were determined after intervals (30,60,90 and 120 days). Data presented in Table 6, by studying each fungus individually reveal that there were significant differences in numbers of A. flavus, A. niger, A. ochraceus and Trichoderma spp. between (the third and forth month) and (the first month) but no significant differences were observed between the first, second, third and forth month during storage period.
| Table 7: | The levels of AFB1 (ppb) in treated Ras cheese during storage for 120 days |
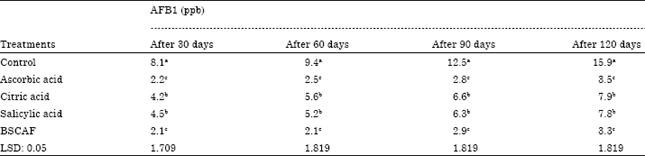 | |
In connection with, the impact of the anti fungi on tested fungi, there were significant differences between control and all tested anti fungi during ripening period, despite the differences between the impacts of these anti fungi. Saleem (2008) found that citric acid and some other food preservatives have a dampening effect on fungi such as A. niger, F. oxysporum and Nectria haematococca.
After the first month, significant differences were found between the effect of all anti fungi and the control. These differences confirmed significantly with increasing storage period. All tested anti fungi affected significantly on the growth of fungi after the first, second, third and forth month. Although, the impact of anti fungi were different on fungal species.
Lastly, AFB1 was determined in manufactured treated Ras cheese by mentioned anti-fungi after 30, 60, 90 and 120 days during storage period. Data published in Table 7 highlighted that control sample (without treatment) was greater contamination than all treated samples. On the other hand, significant differences were found between the effects of all used anti fungi and control on quantities of AFB1. Furthermore, that it is possible to divide the effects of used anti fungi on determined quantities of AFB1 to two sets, the strongest group included (Ascorbic acid and Yeast) followed by the weak group included (Salicylic acid and Citric acid). Yeast was the idealist substrate on decreasing AFB1 in treated Ras cheese whilst recorded the lowest levels 2.1c, 2.1c, 2.9c and 3.3c ppb of AFB1 after 30, 60, 90 and 120 days of storage period at ripening cheese room but citric acid was the lowest effect on reduction levels of AFB1.
CONCLUSION
In this study, many fungi were isolated; levels of AFB1 were detected in cheese samples too. Some antioxidants and BSCAF decreased the fungi number and levels of AFB1. Hence, to control fungi and AFB1 in ripening cheese, it can be recommend using antioxidants private BSCAF, Ascorbic and Salicylic acids at mentioned concentrations in this study.
REFERENCES
- Ashraf, M.W., 2012. Determination of aflatoxin levels in some dairy food products and dry nuts consumed in Saudi Arabia. Food Public Health, 2: 39-42.
CrossRef - Bakirci, I., 2001. A study on the occurrence of aflatoxin M1 in milk and milk products produced in Van province of Turkey. Food Control, 12: 47-51.
CrossRefDirect Link - Beresford, T.P., N.A. Fitzsimons, N.L. Brennan and T.M. Cogan, 2001. Recent advances in cheese microbiology. Int. Dairy J., 11: 259-274.
CrossRefDirect Link - Callon, C., C. Delbes, F. Duthoit and M. Montel, 2006. Application of SSCP-PCR fingerprinting to profile the yeast community in raw milk Salers cheeses. Syst. Appl. Microbiol., 29: 172-180.
CrossRefDirect Link - Chang, S.C., K.L. Lu and S.F. Yeh, 1993. Secondary metabolites resulting from degradation of PR toxin by Penicillium roqueforti. Applied Environ. Microbiol., 59: 981-986.
PubMedDirect Link - EC, 2002. Commission regulation (EC) No 257/2002 of 12 February 2002 amending regulation (EC) No 194/97 setting maximum levels for certain contaminants in foodstuffs and regulation (EC) No 466/2001 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Communities, L41: 12-15.
Direct Link - Dragacci, S., E. Gleizes, J.M. Fremy and A.A.G. Candlish, 1995. Use of immunoaffinity chromatography as a purification step for the determination of aflatoxin M1 in cheeses. Food Additives Contaminants, 12: 59-65.
CrossRefPubMedDirect Link - El-Diasty, E.M. and R.M. Salem, 2007. Incidence of lipolytic and proteolytic fungi in some milk products and their public health significance. J. Applied Sci. Res., 3: 1684-1688.
Direct Link - Elwakil, M.A. and M.A. El-Metwally, 2000. Hydroquinone, a promising antioxidant for managing seed-borne pathogenic fungi of peanut. Pak. J. Biol. Sci., 3: 374-375.
CrossRefDirect Link - Florez, A.B. and B. Mayo, 2006. Microbial diversity and succession during the manufacture and ripening of traditional, Spanish, blue-veined Cabrales cheese, as determined by PCR-DGGE. Int. J. Food Microbiol., 10: 165-171.
PubMedDirect Link - Frisvad, J.C. and R.A. Samson, 2004. Polyphasic taxonomy of Penicillium Subgenus Penicillium-a guide to identification food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol., 49: 1-174.
Direct Link - Frisvad, J.C., J. Smedsgaard, T.O. Larsen and R.A. Samson, 2004. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol., 49: 201-241.
Direct Link - Geiser, D.M., C. Gueidan, J. Miadlikowska, F. Lutzoni and F. Kauff et al., 2006. Eurotiomycetes: Eurotiomycetidae and chaetothyriomycetidae. Mycologia, 98: 1053-1064.
PubMedDirect Link - Gente, S., S. Larpin, O. Cholet, M. Gueguen, J.P. Vernoux and N. Desmasures, 2007. Development of primers for detecting dominant yeasts in smear-ripened cheeses. J. Dairy Res., 74: 137-145.
PubMedDirect Link - Giridhar, P. and S.M. Reddy, 2001. Phenolic antioxidants for the control of some mycotoxigenic fungi. J. Food Sci. Tech. (Mysore), 38: 397-399.
Direct Link - Houbraken, J. and R.A. Samson, 2011. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol., 70: 1-51.
CrossRefDirect Link - Irlinger, F. and J. Mounier, 2011. Microbial interactions in cheese: implications for cheese quality and safety. Curr. Opin. Biotechnol., 20: 142-148.
CrossRefDirect Link - Jahanshiri, Z., M. Shams-Ghahfarokhi, A. Allameh and M. Razzaghi-Abyaneh, 2012. Effect of curcumin on Aspergillus parasiticus growth and expression of major genes involved in the early and late stages of aflatoxin biosynthesis. Iran. J. Public Health, 41: 72-79.
Direct Link - Lopez, C.E., L.L. Ramos, S.S. Ramadan and L.C. Bulacio, 2003. Presence of aflatoxin M1 in milk for human consumption in Argentina. Food Control, 14: 31-34.
CrossRefDirect Link - Lund, F., O. Filtenborg and J.C. Frisvad, 1995. Associated mycoflora of cheese. Food Microbiol., 12: 173-180.
CrossRefDirect Link - Lund, F., A.B. Nielsen and P. Skouboe, 2003. Distribution of Penicillium commune isolates in cheese dairies mapped using secondary metabolite profiles, morphotypes, RAPD and AFLP fingerprinting. Food Microbiol., 20: 725-734.
CrossRefDirect Link - Maurice, O.M., 2002. Risk assessment for aflatoxins in foodstuffs. Int. Biodeterioration Biodegradation, 50: 137-142.
CrossRefDirect Link - Montagna, M.T., C. Napoli, O. De Giglio, R. Iatta and G. Barbuti, 2008. Occurrence of aflatoxin M1 in dairy products in Southern Italy. Int. J. Mol. Sci., 9: 2614-2621.
CrossRef - Saleem, A.R., 2008. Effect of some food preservatives on the lipolytic activity of beef luncheon fungi. Mycobiology, 36: 167-172.
CrossRefPubMedDirect Link - Savova, I. and M. Nikolova, 2002. Isolation and taxonomic study of yeast strains from Bulgarian dairy products. J. Cult. Collections, 3: 59-65.
Direct Link - Sieuwerts, S., F.A. de Bok, J. Hugenholtz and J.E. van Hylckama Vlieg, 2008. Unraveling microbial interactions in food fermentations: from classical to genomics approaches. Applied Environ. Microbiol., 74: 4997-5007.
CrossRefPubMedDirect Link - Masram, C.B. and R.M. Jugade, 2013. Polarographic studies of ascorbic acid and estimation in pharmaceutical formulations and fruit juices. Int. J. Pharm. Sci. Rev. Res., 22: 285-287.
Direct Link