
Mostafa Sayed Khattab
Department of Dairy Science, National Research Centre, Dokki, Cairo, Egypt
S.A.H. Abo El-Nor
Department of Dairy Science, National Research Centre, Dokki, Cairo, Egypt
H.M.A. El-Sayed
Department of Animal Production, Faculty of Agriculture, Ain Shams University, Shoubra, Al-Kheima, Cairo, Egypt
N.E. El-Bordeny
Department of Animal Production, Faculty of Agriculture, Ain Shams University, Shoubra, Al-Kheima, Cairo, Egypt
M.M. Abdou
Department of Dairy Science, National Research Centre, Dokki, Cairo, Egypt
O.H. Matloup
Department of Dairy Science, National Research Centre, Dokki, Cairo, Egypt
International Journal of Dairy Science
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 95-102







ABSTRACT
The increase in bio-ethanol industry has created a need for alternative to corn for ruminants. In the other side there is increase in availability of glycerol, a primary co-product material of biodiesel production. The objective of this study was to investigate the effect of partial replacing of corn with glycerol in diets fed to lactating goats. Twelve lactating Nubian goats were fed a base diet (T1), diets containing 9% glycerol (on DM basis) (T2) and diet containing 9% glycerol plus commercial enzymes 4 g kg-1 DM (on DM basis) (T3) for 84 days. The experimental diets T2 and T3 decreased butyric acids concentration and acetate:propionate ratio in rumen liquor in relation to T1, the concentration of propionic acid was increased in T2 and T3 compared with T1. Replacing corn by glycerol (T2) decreased apparent nutrients digestion coefficients Dry Matter (DM), Organic Matter (OM), Crud Protein (CP), Neutral Detergent Fiber (NDF) and Acid Detergent Fiber (ADF) comparing with other treatments (T1 and T3). Milk production was 1581, 1174 and 1610±77.6 g h-1 day-1 and FCM was 1774, 1030 and 1648±115.9 g h-1 day-1 for T1, T2 and T3, respectively, Milk composition was not altered by glycerol feeding plus fibrinolytic enzymes (T3) except that milk total protein was decreased from 4.6 24 to 3.5%. While, replacing corn by glycerol (T2) decreased values of milk composition compared with control diets (T1). The results indicated that glycerol is a suitable replacement for corn grain with adding fibrinolytic enzymes in diets for lactating goats and that it may be included in rations to a level of at least 9% of dry matter without adverse effects on milk yield or milk composition.
PDF Abstract XML References Citation
Received: July 15, 2012;
Accepted: October 16, 2012;
Published: November 27, 2012
How to cite this article
Mostafa Sayed Khattab, S.A.H. Abo El-Nor, H.M.A. El-Sayed, N.E. El-Bordeny, M.M. Abdou and O.H. Matloup, 2012. The Effect of Replacing Corn with Glycerol and Fibrinolytic Enzymes on the Productive Performance of Lactating Goats. International Journal of Dairy Science, 7: 95-102.
DOI: 10.3923/ijds.2012.95.102
URL: https://scialert.net/abstract/?doi=ijds.2012.95.102
DOI: 10.3923/ijds.2012.95.102
URL: https://scialert.net/abstract/?doi=ijds.2012.95.102
INTRODUCTION
The high global demand for energy led to an increasing production and trade of biofuel, especially liquid fuels for transportation, in order to substitute fossil energy sources, to enhance energy security and to respond to greenhouse gas emissions (Walter et al., 2008; Heinimo and Junginger, 2009). This led to an increasing competition for raw materials for food, feed and fuel usage and the disposability of more by-products. For example, starch and vegetable oils are converted into bioethanol and biodiesel during the production process, one of the by-products of this process is glycerol. Glycerol is an essentially structure component of triglycerides and phospholipids and its glucogenic properties are well established (Cori and Shine, 1935). Glycerol enters the metabolic pathway to glucose at a different step than other glucogenic precursors (Leng, 1970). The glycerol component can be converted to glucose by the liver or kidneys (Krebs and Lund, 1966) to provide energy for cellular metabolism. Glycerol has been shown to be an effective treatment against ketosis in dairy cattle (Leng, 1970). Cows fed glycerol lost less body weight and remained in more positive energy balance (DeFrain et al., 2004), Several researchers (Schroder and Sudekum, 1999; Mach et al., 2009) estimated the energy value of glycerol in beef and they have concluded to be similar to that of corn grain. Therefore, glycerol could be used as an energetic ingredient in animal diets replacing cereals. The inclusion of glycerol in animal nutrition has been studied in several species and reports indicate that moderate inclusions do not compromise animal performance (Lammers et al., 2008; Mach et al., 2009).
Many in vitro and in vivo studies carried-out to investigate glycerol as feed on rumen fermentation and concluded that glycerol increase total volatile fatty acids, propionate and butyrate concentrations and decreases the molar ratio of acetate: propionate concentration (Schroder and Sudekum, 1999; Goff and Horst, 2001; Farkasova et al., 2008; Abo El-Nor et al., 2010). The objective of the present study is to find alternative source energy could be used in ruminant diets by evaluating the effects of substituting corn by glycerol with some biological additives (Fibrinolytic enzymes) on productive performance of lactating goats.
MATERIALS AND METHODS
This study was carried out to investigate the effect of replacing corn by glycerol alone or glycerol with a biological additive comparing with the control diet on the productive performance of lactating Nubian goats.
Digestibility and lactation trails: Twelve lactating Nubian goats ranging from 35-40 kg were assigned randomly into three experimental groups (four animals each) using Latin Square design. The experimental extended for 84 days after 7 days of parturition and consisted of three equal experimental periods (28 days each). The goats were fed individually according to nutrients requirement recommendations of NRC (1981). The dietary forage: concentrate ratio was 38:62. Forage source was Egyptian clover (Trifolium alexanderinum) (Table 1).
| Table 1: | Ingredient and nutrient composition of diets |
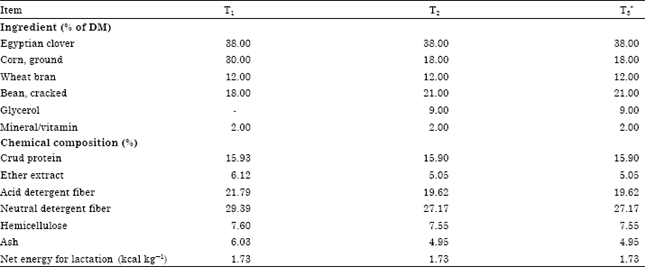 | |
| *Enzymes (15000 cellulase enzyme unit, 187500 protease enzyme unit, 300000 α-amylase enzyme unit, 15000 β-amylase enzyme unit, 75x108 CFU of Bacillus subtilis) add at rate of 4 g kg-1 DM | |
The experimental treatments were: control fed on 38% Egyptian clover and 62% Concentrate Feed Mixture (CFM) without glycerol (T1); glycerol group fed as control except replacing 30% corn with glycerol (T2) and the third group (T3) were fed as glycerol group with adding enzymes (4 g kg-1 DM). Diets were offered twice daily at 08:00 and 16:00 h. water was available continuously.
At the last day of each experimental period, rumen liquor samples were collected from each animal at zero, 3 and 6 h post-morning feeding by stomach tube. The samples were strained through two layers of cheese cloth and were immediately used for determination of ruminal pH. Rumen liquor samples were stored in glass bottles after adding metaphosphoric acid (15%) and stored at deep freeze (-18°C) for analysis of ammonia and volatile fatty acids fraction. Feces grab sample method was used and silica as internal marker was applied for determining the apparent digestibility. Feces grab samples were collected at 07:00 h for three successive days from each animal. Solution of 10% H2SO4 were added to the representative samples then dried in oven at 70°C for 24 h. The dried feces samples from each animal were mixed and stored at -18°C for chemical analysis of DM, OM, CP, NDF, ADF and ash. The animals were handily milked twice daily at 07:00 and 19:00 h during the last three days of each experimental period. After each milking, sample of each animal represents a mixed sample of constant percentage of the evening and morning yield and stored at (-18°C) for further analysis of milk constituents.
Chemical analysis
Feed and feces samples analysis: The dry matter contents of feed and faeces were determined by oven-drying for 4 h at 105°C according to AOAC (1990) method No. 930.15. Ash analysis was conducted at 550°C for 4 h based on the AOAC (1990) method No. 942.05. Nitrogen was measured using a mixed catalyst Kjeldahl method AOAC (1990); method No. 988.05). The crude protein content was calculated by multiplying nitrogen by 6.25. Ether extract was determined by the Soxhlet method with petroleum ether as a solvent following AOAC (1990) method No. 963.15. The TMR samples were also analyzed for ADF (method 973.18c; AOAC (1990) and NDF (Van Soest et al., 1991) using α-amylase (A3306; Sigma Chemical Co., St. Louis, MO) and sodium sulfite corrected for ash concentration adapted for an Ankom 200 fiber analyzer (Ankom Technology, Fairport, NY).
Rumen liquor samples analysis: Samples for VFA analyses were prepared and analyzed as described by Bush et al. (1979). Ammonia samples of rumen liquor were centrifuged at 2000X g at 4°C for 10 min and the supernatant was acidified with 0.5 mL of 0.1 N HCl then analyzed for NH3 by a Bio-Diagnostic kit (Bio-Diagnostic, Cairo, Egypt) using a spectrophotometer (T80 UV/Vis spectrophotometer, PG Instruments Ltd., England).
Milk samples analysis: Milk samples were analyzed for Total Solids (TS), Solids Not Fat (SNF), Total Protein (TP), fat and lactose using infrared spectroscopy (Bentley 150, Infrared Milk Analyzer, Bentley Instruments, USA).
Statistical analysis: Data were statistically analyzed using the ANOVA procedures of SAS according to procedures outlined by Snedecor and Cochran (1982), significant level was 0.05.
RESULTS
The ingredients and chemical composition of the experimental diets are presented in Table 1. Dietary ADF, NDF and ether extract content decreased as proportion of glycerol in diets. Dietary CP and net energy for lactation (NEL) content were similar among experimental diets.
Rumen fermentation parameters: Rumen pH values for the experimental treatments through different sampling times showed that replacing corn by glycerol tends to slightly decrease rumen pH comparing with control (T1) (Table 2). These results indicated that there were insignificant (p>0.05) differences among the overall mean of rumen pH in different treatments being 6.42, 6.43 and 6.31 for T1, T2 and T3, respectively. The molar proportion for acetate decreased (p<0.05) with feeding glycerol (T2) recorded the lowest value (44.56%) compared with T1 (46.96%), while T3 increased the value of acetic acid value (47.93%). The pattern of propionic acid concentration had showed that T2 recorded the highest value (33.1%) then T3 (28.43%) then T1 (26.33%). The acetate to propionate ratio decreased (p<0.05) with the T2 diet relative to the T1 and T3 diet. The molar proportions for butyrate decreased with feeding glycerol (T2 and T3) compared with T1, the recorded values for butyric acid concentration were 23.66, 21.33 and 22.23% for T1, T2 and T3, respectively. The overall means of NH3 concentration were lower significantly (p<0.05) in T2 compared to T1 and T3 being 12.33, 8.83 and 10.2 100 mg mL-1 for T1, T2 and T3, respectively.
| Table 2: | Effect of experimental diets on ruminal parameters |
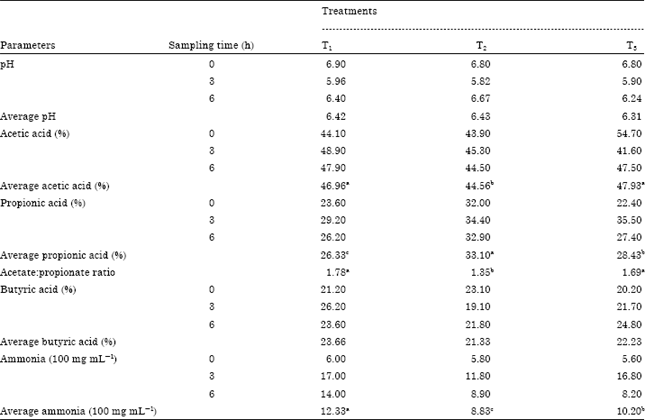 | |
| T1: Total mixed ration consist of 62% concentrate feed mixture (CFM) and 38% roughage (Egyptian clover), T2: Replacing 30% of diet corn with crude glycerol, T3: Replacing 30% of diet corn with crude glycerol plus 4 g kg-1 DM enzymes, Means in the same row with different superscripts are significantly differ at p<0.05 | |
| Table 3: | Effect of experimental diets on nutrients digestibility |
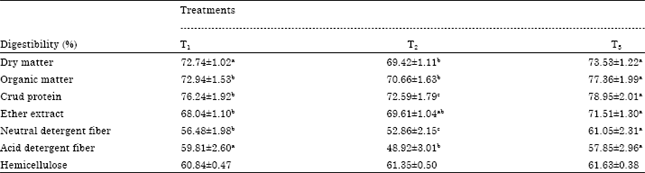 | |
| T1: Total mixed ration consist of 62% concentrate feed mixture (CFM) and 38% roughage (Egyptian clover), T2: Replacing 30% of diet corn with crude glycerol, T3: Replacing 30% of diet corn with crude glycerol plus 4 g kg-1 DM enzymes, Means in the same row with different superscripts are significantly differ at p<0.05 | |
| Table 4: | Effect of experimental diets on milk yield and composition |
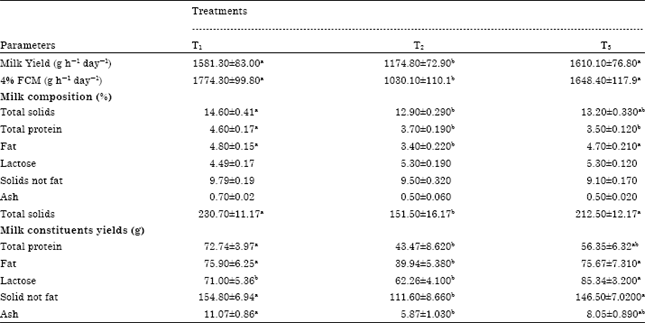 | |
| T1: Total mixed ration consist of 62% concentrate feed mixture (CFM) and 38% roughage (Egyptian clover), T2: Replacing 30% of diet corn with crude glycerol, T3: Replacing 30% of diet corn with crude glycerol plus 4 g kg-1 DM enzymes, Means in the same row with different superscripts are significantly differ at p<0.05 | |
Digestibility coefficients: The results of Table 3 showed a significant (p<0.05) decrease in digestion coefficients of DM, OM, CP, NDF and ADF of T2 compared with T1, while there was no significant differences (p<0.05) between T3 and T1 in DM, ADF and hemicellulose digestibility, while adding fibrinolytic enzymes to glycerol diet (T3) significantly (p<0.05) increased OM, CP, NDF and Ether Extract (EE) digestibility comparing with T1.
Milk yield and composition: Goats fed glycerol (T2) produced lower amount of milk and FCM (p<0.05) during the feeding period as goats fed a diet containing no glycerol (T1) or containing glycerol with fibrinolytic enzymes (T3) (Table 4). Milk composition was altered to be significantly reduced (p<0.05) in response to glycerol feeding except for lactose (p>0.05). Milk total solids, total protein, SNF and fat were decreased (p<0.05) in case T2 which replaced corn by glycerol compared with control (T1) while adding fibrinolytic enzymes has a significant effect on the milk composition of T3.
DISCUSSION
These results indicated that there were insignificant (p>0.05) differences among the overall mean of rumen pH in different treatments being 6.42, 6.43 and 6.31 for T1, T2 and T3, respectively. T2 recorded the lowest value through 3 hrs post-feeding which reflect the high fermentation potential of glycerol in the rumen and produced more propionate which contributes a decrease in pH value. Roger et al. (1992) reported that the addition of glycerol to the in vitro media greatly inhibited the growth and cellulolytic activity of rumen bacteria and fungi. Paggi et al. (2004) reported that the cellulolytic activity of ruminal extract was reduced as glycerol concentration in rumen cultures increased from 50 to 300 mmol. The reduction in the molar proportion of acetate and acetate to propionate ratio in T2 was consistent with the reduction in NDF digestibility. Studies that have reported reduction in NDF digestibility have also reported reductions in acetate concentration and acetate to propionate ratio (Ribeiro et al., 2005; Castillejos et al., 2006). The decrease in NH3 in the rumen of glycerol groups might be due to the reduction of proteolytic activity of rumen microorganisms (Paggi et al., 1999) who found that adding glycerol reduced proteolytic activity by 20%, the glycerol has three hydroxyl groups, but unlike VFA and to a lesser extent lactic acid, glycerol lacks a hydrophobic chain. This lack would make the medium where glycerol is dissolved less suitable for the activity of the proteolytic enzymes. This latter effect of the structure of glycerol would predominate over its effect on the solubility of the substrate rendering, as a balance, a depressive effect on the proteolytic activity of rumen microorganisms. Glycerol also reduced the proteolytic activity of bovine rumen fluid by about 20% when concentrations of glycerol in the medium (Paggi et al., 1999). Glycerol in excess of 1% in bacterial culture media (in vitro) inhibited growth and cellulolytic activity of Ruminococcus flavefaciens and Fibrobacter succinogenes Roger et al. (1992).
The tendency for a reduction in NDF digestibility 183 was matched by reductions in OM or DM digestibility. These data contrast with previous observations of Donkin et al. (2009) which reported a notable increase in DM, OM and nitrogen digestibility with addition of glycerol to lactating cow diets at 15% of DM.
The reduction of milk yield and FCM values might be due to replacing corn by glycerol and its negative effects on rumen microflora (as microbial protein) and activity and the reduction in DM, OM, CP, NDF and ADF digestion coefficients, while, in T3 adding fibrinolytic enzymes to glycerol enhance the animals' response which clears in the return of nutrients digestion value of DM, OM, CP, NDF, ADF and EE compared to the control diet (T1). The reduction of rumen micro-flora biomass and activity caused the significant reduction in nutrients digestion coefficients especially NDF, ADF and CP which lead the observed reduction in milk yield and milk constituents’ reduction mainly milk total solids, fat and protein. While the slight increase (p>0.05) of lactose percent in case of inclusion of glycerol in the diet might be due to propionic acid which is the main volatile fatty acid derived from glycerol, supporting the glycogenic role of glycerol in ruminants (Johns, 1953; Garton et al., 1961).
CONCLUSION
From the results of these experiments it is clear that replacing corn by glycerol at relatively high level (9% of DM) required addition of the fibrinolytic enzymes to avoid the negative effect of glycerol on the digestion and productive performance of lactating goats.
REFERENCES
- Bush, K.J., R.W. Russell and J.W. Young, 1979. Quantitative separation of volatile fatty acids by high-pressure liquid chromatography. J. Liquid Chromatogr., 2: 1367-1374.
CrossRef - Castillejos, L., S. Calsamiglia and A. Ferret, 2006. Effect of essential oil active compounds on rumen microbial fermentation and nutrient flow in in vitro systems. J. Dairy Sci., 89: 2649-2658.
CrossRefDirect Link - Cori, C.F. and W.M. Shine, 1935. The formation of carbohydrate from glycerophosphate in the liver of the rat. Science, 82: 134-135.
CrossRefDirect Link - DeFrain, J.M., A.R. Hippen, K.F. Kalscheur and P.W. Jardon, 2004. Feeding glycerol to transition dairy cows: Effects on blood metabolites and lactation performance. J. Dairy Sci., 87: 4195-4206.
CrossRefDirect Link - Donkin, S.S., S.L. Koser, H.M. White, P.H. Doane and M.J. Cecava, 2009. Feeding value of glycerol as a replacement for corn grain in rations fed to lactating dairy cows. J. Dairy Sci., 92: 5111-5119.
CrossRefDirect Link - Farkasova, Z., P. Reichel, C. Tothova, V. Hisira and G. Kovac, 2008. Effect of feeding glycerol on rumen fluid and the parameters of the energetic profile in dairy cows. Medycyna Wet., 64: 770-772.
Direct Link - Garton, G., A. Lough and E. Vioque, 1961. Glyceride hydrolysis and glycerol fermentation by sheep rumen contents. J. Gen. Microbiol., 25: 215-225.
PubMedDirect Link - Heinimo, J. and M. Junginger, 2009. Production and trading of biomass for energy-an overview of the global status. Biomass Bioenergy, 33: 1310-1320.
CrossRefDirect Link - Krebs, H.A. and P. Lund, 1966. Formation of glucose from hexoses, pentoses, polyols and related substances in kidney cortex. Biochem. J., 98: 210-214.
PubMedDirect Link - Lammers, P.J., B.J. Kerr, M.S. Honeyman, K. Stalder and W.A. Dozier et al., 2008. Nitrogen-corrected apparent metabolizable energy value of crude glycerol for laying hens. Poult. Sci., 87: 104-107.
CrossRefDirect Link - Mach, N., A. Bach and M. Devant, 2009. Effects of crude glycerin supplementation on performance and meat quality of Holstein bulls fed high-concentrate diets. J. Anim. Sci., 87: 632-638.
CrossRefPubMedDirect Link - Paggi, R.A., J.P. Fay and C. Faverin, 2004. In vitro ruminal digestibility of oat hay and cellulolytic activity in the presence of increasing concentrations of short-chain acids and glycerol. J. Agric. Sci., 142: 89-96.
CrossRefDirect Link - Paggi, R.A., J.P. Fay and H.M. Fernandez, 1999. Effect of short-chain acids and glycerol on the proteolytic activity of rumen fluid. Anim. Feed Sci. Technol., 78: 341-347.
CrossRefDirect Link - Ribeiro, C.V.D.M., S.K.R. Karnati and M.L. Eastridge, 2005. Biohydrogenation of fatty acids and digestibility of fresh alfalfa or alfalfa hay plus sucrose in continuous culture. J. Dairy Sci., 88: 4007-4017.
CrossRefDirect Link - Roger, V., G. Fonty, C. Andre and P. Gouet, 1992. Effects of glycerol on the growth, adhesion and cellulolytic activity of rumen cellulolytic bacteria and anaerobic fungi. Curr. Microbiol., 25: 197-201.
CrossRefDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Walter, A., F. Rosillo-Calle, P. Dolzan, E. Piacente and K. Borges da Cunha, 2008. Perspectives on fuel ethanol consumption and trade. Biomass Bioenergy, 32: 730-748.
CrossRefDirect Link