
H. Tormo
Universite de Toulouse, Ecole d`Ingenieurs de Purpan, 75, voie du TOEC, BP 57611, F-31076 Toulouse Cedex 03, France
A. Delacroix-Buchet
INRA (UMR 1319) and AgroParisTech, Institut Micalis, F-78352 Jouy-en-Josas, France
C. Lopez
Institut de l`Elevage, 149 rue de Bercy-75595 Paris Cedex 12, France
D. Ali Haimoud Lekhal
Universite de Toulouse, Ecole d`Ingenieurs de Purpan, 75, voie du TOEC, BP 57611, F-31076 Toulouse Cedex 03, France
C. Roques
Universite de Toulouse; UPS, LU 49, Adhesion bacterienne et formation de biofilms, 35 chemin des maraichers, F-31062 Toulouse cedex 9, France
International Journal of Dairy Science
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 29-43







ABSTRACT
Studies on cow’s milk have already underlined some relationships between management practices and the microbial quality of milk, but only a few have been carried out on goat’s milk. The aim of this study was: (1) to determine the main bacterial species present in goat’s milk and (2) to identify management practices that may have a potential influence on the bacterial species in the milk. The bacterial species present in samples of raw goat’s milk from 30 farms in different regions were characterized by PCR-Temporal Temperature Gradient gel Electrophoresis (TTGE) and PCR-Denaturing Gradient Gel Electrophoresis (DGGE). The predominant bacterial species belonged to the genera Staphylococcus, Arthrobacter and Serratia. Lactic acid bacteria (Lactococcus lactis and Enterococcus faecalis) were only detected in 25% of the milk samples. Brevibacterium linens were detected systematically in milk from zero-grazing herds. High standards of hygiene with regard to milking machine cleaning considerably reduced the number of dominant species in the milk; lactic acid bacteria, in particular were no longer detected. On the other hand, inferior hygiene increased the number of species present and E. faecalis was more commonly found. Herd health management was a determining factor in the microbial contamination of the milk samples studied. Inferior health conditions increased contamination of the milk with Serratia and Staphylococcus. Direct contact between the milking parlour and the goats’ housing appeared to enrich the milk in L. lactis. Present results underlined that it may be possible to influence the balance between the bacteria present in milk using certain practices.
PDF Abstract XML References Citation
Received: September 17, 2010;
Accepted: October 23, 2010;
Published: December 27, 2010
How to cite this article
H. Tormo, A. Delacroix-Buchet, C. Lopez, D. Ali Haimoud Lekhal and C. Roques, 2011. Farm Management Practices and Diversity of the Dominant Bacterial Species in Raw Goat's Milk. International Journal of Dairy Science, 6: 29-43.
DOI: 10.3923/ijds.2011.29.43
URL: https://scialert.net/abstract/?doi=ijds.2011.29.43
DOI: 10.3923/ijds.2011.29.43
URL: https://scialert.net/abstract/?doi=ijds.2011.29.43
INTRODUCTION
The paramount role of the microflora on the sensory characteristics of cheese has been demonstrated in many studies on various cheese technologies. Suppression of the microflora in raw milk (by microfiltration or pasteurization) causes a reduction in the flavour and aromatic differences of the resulting cheese (Beuvier et al., 1997; Bouton and Grappin, 1995; Buchin et al., 1998; Verdier-Metz et al., 2002). In addition to the value that certain bacterial populations present in raw milk can provide in terms of the sensory qualities of the cheese, they have also been shown to have an inhibitory effect on pathogenic microorganisms (El-Gazzar et al., 1992; Gay and Amgar, 2005). In France, payment according to milk quality stipulates low microbial burden as a measure of milk quality, the priority being to reduce the numbers of microorganisms that cause spoilage or are potentially pathogenic. This necessary reduction in the levels of unwanted microorganisms has undoubtedly modified the composition of the bacterial flora that is useful to cheese-making technology. A few studies, most of which were on cow’s milk, have shown that the balance between the spoilage or pathogenic bacterial flora and the bacterial flora of technological interest is influenced by farm management practices, particularly those concerning udder cleGayaning and disinfection and milking hygiene (Michel et al., 2001, 2005). Verdier-Metz et al. (2009) also showed that the diversity of bacterial species in raw milk declined significantly when standards of hygiene (from teat preparation to milk production) were high. Only coryneform bacteria were detected in these milk samples. This imbalance and in particular the absence of lactic acid bacteria, can decrease the sensorydiversity of the resulting cheese. Very few studies have been conducted on this subject in goats and they investigated only a small number of farms (Allonso-Calleja et al., 2002; Foschino et al., 2002; Callon et al., 2007). In addition, farm management practices, from the goats’ housing to milk production, have not been evoked to explain the variability of the bacterial characteristics of milk. The recent development of DNA fingerprinting techniques has enabled the use of another approach to analysing the bacterial communities present in a medium (Coppola and Ercolini, 2008). These methods are based on DNA extraction without prior microbial culture. This gives a comprehensive snapshot of the dominant bacterial communities present, without previously selecting certain microbial groups at the expense of others, as occurs in culture-dependent techniques. These culture-independent molecular techniques have been applied to the assessment of microbial diversity in milk. This is the case in Single Strand Conformation Polymorphism (SSCP) (Callon et al., 2007; Verdier-Metz et al., 2009), Temporal Temperature Gradient gel Electrophoresis (TTGE) and Denaturing Gradient Gel Electrophoresis (DGGE) techniques (Ogier et al., 2002, 2004; Lafarge et al., 2004; Ercolini et al., 2009; Giannino et al., 2009). The objectives of this study were to elucidate the profiles of the dominant bacterial communities in raw goat’s milk samples and the relationships between these bacterial communities and farm management practices (from goat housing to milk production). This study was carried out under real herd management conditions among 30 farmers of PDO (Protected Designation of Origin) farmhouse Pelardon and Rocamadour cheeses, respectively. TTGE and DGGE techniques were used to obtain a rapid, comprehensive view of the predominant bacterial species present in the samples of raw goat’s milk. The relationship between the bacterial species profile and the farm management practices was investigated by statistical analysis.
MATERIALS AND METHODS
Study description:The present work was conducted in the Languedoc Roussillon and Midi-Pyrenees regions among producers of PDO (Protected Designation of Origin) farmhouse Pelardon (11 farms) and Rocamadour cheeses (19 farms), respectively. This study was carried out under real herd management conditions. For the best representation of breeding and milking production practices, the farms were selected by the Rocamadour and Pelardon cheese producers’ trade unions. The monitoring period was May and June 2006, because there are great differences between farms in terms of management practices, for example the daily temperature ranged from 18 and 25°C and the lactation stage from 3 to 5 months for 70% or more of the goats in each flock.
Data collection: For each farm, a survey, observations and measurements were carried out by the investigators. Variables were categorized in seven families: (1) general characteristics of the farm including the region, (2) bedding management practices, (3) environmental conditions during and after milking, (4) cleaning practices and cleanness of the Milking Machine (MM), (5) MM handling and characteristics, (6) quality and cleaning of the teat and (7) cleaning practices of the dairy and milk tank.
Raw milk samples: For each period, three milk samples from the evening milking were collected from each farm. The 3 samples were collected on days 1, 4 and 8 from the tank, cooled to 10°C and stored at -25°C for 15 days. A total of 90 samples were analysed
Temporal Temperature Gradient gel Electrophoresis (TTGE) and Denaturing Gradient Gel Electrophoresis (DGGE)
DNA extraction: Sixty milliliter of raw milk samples were digested by addition of 50 mg of pronase (Roche Diagnostics, Meylan, France) and 100 μL of β-mercaptoethanol (Serva, Heidelberg, Germany) and were incubated for 3 h in a 52°C water bath. Bacterial pellets were obtained by centrifugation at 12000 g for 20 min at 4°C and were washed once with sterile water and once with 10 mL of TES buffer (25 Mm Tris-HCl, 0.1 M EDTA, 25% [wt/vol] saccharose; pH 8). Supernatants were discarded and bacterial pellets were stored at -20°C. Before experimentation, bacterial pellets were resuspended in 500 μL of TES and the bacteria were mechanically lysed with zirconium beads (diameter, 150 to 200 μm; Sigma, St. Louis, Mo.) by three cycles of 30 sec of vortexing with 1 min of storage in ice between each cycle. DNA purification was performed as described previously (Delos Reyes-Gavilan et al., 1992).
PCR amplification: The V3 region of the 16S RNA gene is the substrate for PCR amplification. PCR amplification was performed as described by Lafarge et al. (2004).
TTGE and DGGE analysis for identification of bacteria with AT-rich genomes (TTGE) and GC-rich genomes (DGGE): TTGE and DGGE analysis were performed as described previously (Lafarge et al., 2004).
Analysis of TTGE and DGGE gels: GelCompar software (Applied-Maths, Sint-Martens-Latem, Belgium) was used to analyse TTGE and DGGE gels. Standardization markers were included in all gels with a standard gel to adjust for migration difference between gels. Bacterial species isolated from dairy products were then identified by comparison with a reported reference dairy bacteria database (Ogier et al., 2002, 2004).
Statistical analysis of the data: The TTGE and DGGE profiles were encoded for analysis: all the bands (species) was recorded and scored as 1 if present or 0 if absent for each milk profile.
Descriptive analysis of the bacterial species present in the milk samples: A descriptive analysis of the milk samples was carried out: mean frequency of detection of the various species in the milk samples and number of farms where the species was detected in at least two of the three milk samples collected. Correlations between species were investigated.
Cluster analysis of the bacterial species present in the milk samples: Classes of milk bacterial species were defined. This was achieved though Principal Component Analysis (PCA), then Agglomerative Hierarchical Clustering (AHC) on the PCA factors, followed by classification on the basis of an optimal cut-off level. Bacterial species exhibiting little variability across milk samples from the various farms (i.e., where 90% of milk samples were positive or negative for the species in question) were introduced into the analyses as illustrative PCA variables. This analysis was carried out using SPAD software (version 5.5). In order to study the relationships between the bacterial species classes and farm management practices, it was necessary to assign farms to the class that corresponded to the majority of their milk samples (where two or three of the three milk samples from the farm belonged to the same class). Where the farm’s three milk samples belonged to three different classes, the data on this farm were not included in the following analysis.
Identification of farm management practices predictive of milk bacterial profiles: Two approaches were used to investigate the relationships between the presence of bacterial species and farm management practices.
| • | Identification of farm management practices predictive of bacterial species class Farm management practices were classified according to their ability to discriminate, in a χ2 test, between the farms’ classes of representative bacterial species. |
| • | Identification of farm management practices predictive of the presence of bacterial species |
The probability of the presence of bacterial species when certain farm management practices were used was described using a mixed logistic model (GLIMMIX procedure, SAS 9.1) adjusted for the farm effect. Logistic regression is a statistical technique that aims to produce a model from a set of observations that is capable of predicting the values of a categorical variable from a series of continuous and/or binary explanatory variables. In our case, the categorical variable is the microbial species and it has two possible values: no detection or detection. The explanatory variable, the farm management practices, is binary. The effect of farm management practices on the presence of the bacterial species was estimated using their odds ratio. The odds ratio is the ratio of the probability of occurrence of an event (in this case, detection of a species) to the probability of non-occurrence. The between-farm variance relative to the total variance of random effects (sum of the between-farm variances and within-farm variance) is estimated by the coefficient ρ (%). This descriptor is used to assess how much of the variability in the detection of a microbial species is due to between-farm variability and by subtraction, how much is due to within-farm variability. When within-farm variability is high, the effect of the factor in question is reduced.
RESULTS
Some samples could not be processed due to problems with extracting microbial DNA, despite repeated attempts. The results therefore pertain to 81 of the 90 milk samples taken at the 30 farms.
| Table 1: | Frequency of detection of bacterial species (in ascending order) in the milk samples and the number of farms where the various species were detected |
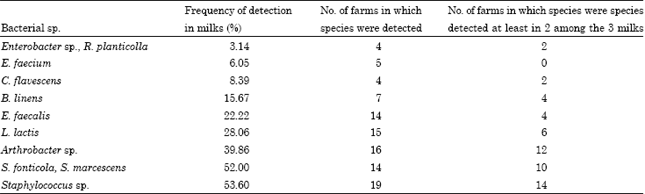 | |
Descriptive analysis of each species and correlation between bacterial species: Three milk samples were analysed for 21 of the farms and 2 milk samples were analysed for 9 of the farms. Analysis of the TTGE and DGGE profiles revealed the predominance of staphylococci in the milk samples analysed (Table 1). Staphylococci were detected in 54% of the samples and were found in samples from 19 farms, including 14 where they were detected in at least 2 of the 3 milk samples from the farm. Serratia fonticola and Serratia marcescens were also dominant species. They were found in 52% of the milk samples and in milk from 14 of the farms, including 10 where they were detected in at least 2 of the 3 milk samples analysed. The next most predominant type was Arthrobacter, followed by Lactococcus lactis and Enterococcus faecalis. Detection of the latter two species was rather sporadic for farms where they were detected in the milk, so although L. lactis was detected in milk from 15 farms, it was only present in at least 2 of the 3 milk samples from 6 of them. Similarly, E. faecalis was detected in milk from 14 farms, but was only present in at least 2 of the 3 milk samples from 4 of them. Other species were detected at low frequency (less than 20%).
The correlations between species (R values) were low. Significant positive correlations were found between: S. fonticola, S. marcescens and C. flavescens (R = 0.319, p = 0.004), S. fonticola, S. marcescens and Arthrobacter (R = 0.273, p = 0.014), E. faecalis and Enterobacter or Raoultella planticola (R = 0.364, p = 0.001). A negative correlation was found between L. lactis and Brevibacterium linens (R = -0.275, p = 0.013).
Description of the bacterial species classes: The variability of the distribution of the bacterial species in milk samples was assessed by performing a cluster analysis of the samples on the basis of the bacterial species detected.
The milk samples from the 30 farms studied fell into five bacterial species classes. These classes accounted for 56% of the total variability of the milk bacterial species. The species that contributed to the construction of the classes were: Staphylococcus spp., S. fonticola and S. marcescens, L. lactis and E. faecalis. The other species were introduced as supplementary variables.
The data on these classes are shown in Fig. 1 to 5.
| • | Class A (Fig. 1): Fifteen milk samples from 11 farms that contained only 1 to 3 of the 9 species detected. These milk samples therefore exhibited very low species diversity |
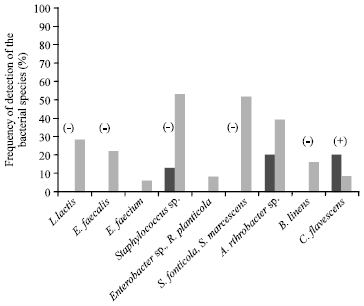 | |
| Fig. 1: | Mean frequency of detection of the bacterial species in class A milk samples and in all the samples. The signs between brackets indicate a significant difference (p<0.05) of the frequency of the species between class A and the total sample (+higher frequency, - lower frequency) |
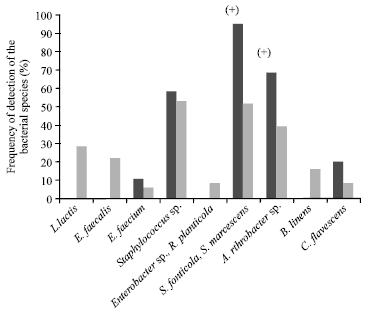 | |
| Fig. 2: | Mean frequency of detection of the bacterial species in class B milk samples and in all the samples. The signs between brackets indicate a significant difference (p<0.05) of the frequency of the species between class B and the total sample (+higher frequency, - lower frequency) |
| • | Class B (Fig. 2): Ninteen milk samples from 13 farms that predominantly contained S. fonticola and S. marcescens (95% of milk samples on average) and Arthrobacter (68%). No enterococci (E. faecalis and E. faecium) or L. lactis were detected in these samples |
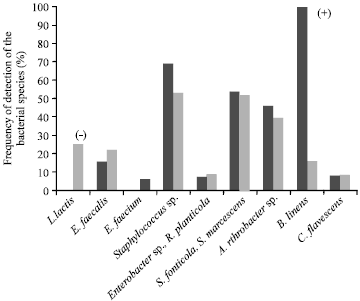
| • | Class C (Fig. 3): Eighteen milk samples from 11 farms, which all contained L. lactis. The other species were detected at a frequency identical to the total sample, except for E. faecalis and B. linens, which were not detected in this class |
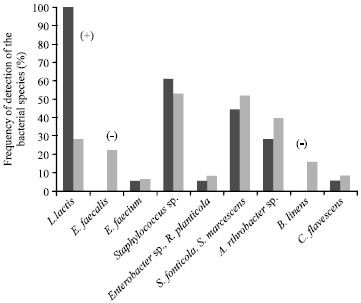 | |
| Fig. 3: | Mean frequency of detection of the bacterial species in class C milk samples and in all the samples. The signs between brackets indicate a significant difference (p<0.05) of the frequency of the species between class C and the total sample (+ higher frequency, - lower frequency) |
 | |
| Fig. 4: | Mean frequency of detection of the bacterial species in class D milk samples and in all the samples. The signs between brackets indicate a significant difference (p<0.05) of the frequency of the species between class D and the total sample (+ higher frequency, - lower frequency) |
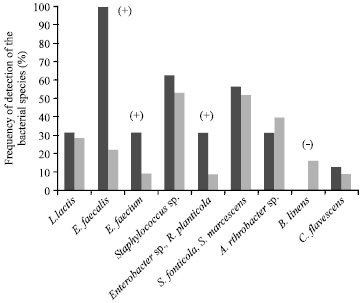
| • | Class D (Fig. 4): Thirteen milk samples from 7 farms. It was the most stable class in terms of the bacterial species profiles found on each farm, with 4 of the 7 farms where this class was found having 2-3 class D samples (i.e., 57% of farms where at least 2 samples were in the same class, compared to 23-36% for the other classes). These samples all contained B. linens, while the other classes did not. The frequency of detection of the other species was the same for all the milk samples analysed, except for L. lactis and E. faecium which were not detected |
 | |
| Fig. 5: | Mean frequency of detection of the bacterial species in class E milk samples and in all the samples. The signs between brackets indicate a significant difference (lower or higher frequency) between class E and the total sample (p<0.05) |
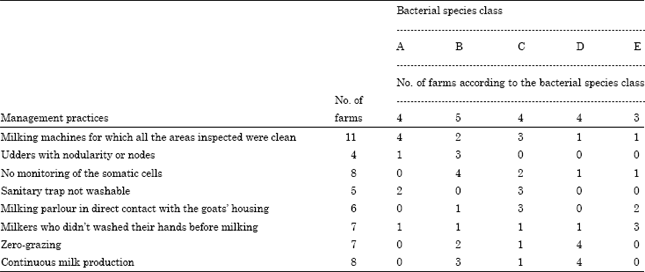
| Table 2: | Breakdown of the farms according to management practices and bacterial species class (the numbers in bold type are farms associated with an over- or under-represented microbial species class (p<0.05) |
 | |
| • | Class E (Fig. 5): Sixteen milk samples from 13 farms. This class was characterized by systematic detection of E. faecalis and higher than average detection of E. faecium, Enterobacter sp. and R. planticola. It is the class with the highest number of microbial species (8 of the 9 species detected) |
Identification of farm management practices predictive of bacterial species class: Twenty of the 30 farms were assigned to the bacterial species profile classes. It was not possible to assign the ten remaining farms to a predominant class because their three milk samples belonged to three different bacterial species classes.
Some specific associations were demonstrated between the farms in the various bacterial species classes and their farming practices (Table 2). Bacterial species class A contained 4 farms. This class, which was characterized by low bacterial species diversity, showed a significant association with farms where the milking machine was clean (12 inspected areas found to be clean).
Bacterial species class B, which was characterized by a higher rate of detection of S. fonticola, S. marcescens and Arthrobacter and no detection of lactic acid bacteria (L. lactis, Enterococcus spp.), was significantly associated with farms where udder trauma (eversion, nodules, enlarged lymph nodes) was found (3 of the 5 class B farms). The farmers in this class (4 of the 5 class B farms) did not monitor somatic cell counts in their herds.
Class C, characterized by the systematic presence of L. lactis, contained 4 farms. On 3 of the 4 farms, the milking parlours were in direct contact with the goat housing and had no washable sanitary trap. Three of the 4 class C farms had a clean milking machine.
Class D, in which milk systematically contained B. linens, contained 4 farms. These farms all used zero-grazing and did not halt production in Winter.
Class E, which exhibited the greatest species diversity and in which E. faecalis was systematically detected, was significantly associated with milkers not washing their hands before milking. Furthermore, as for class D, it was predominantly composed of farms where it was considered that milking machine cleaning was inadequate. Indeed, only 2 of the 7 farms in class D and E had milking machines that were considered clean.
The farms’ class assignment was independent of the geographical region in which they were located.
Identification of farm management practices predictive of the presence of bacterial species. The results of the statistical analysis using the mixed logistic model are summarized in Table 3. A significant association was found between detection of L. lactis and the location of the milking parlour. Indeed, the probability of detecting L. lactis (value of the odds ratio) was 4.2-fold higher when the milking parlour was in direct contact with the goats’ housing. This association was also found with the practices of the representative class C farms, where L. lactis was detected in all the milk samples.
| Table 3: | Farm management practices significantly associated with bacterial species |
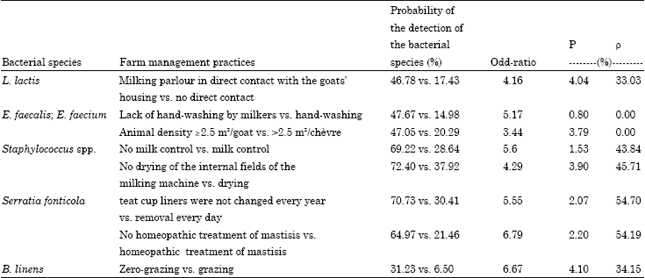 | |
The greatest association with the detection of enterococci was lack of hand-washing by milkers: the relative risk compared to milkers who washed their hands was 5.2. This association was also found with class D farms, the class in which enterococci were dominant.
The mean relative risk for the detection of staphylococci in milk on farms that did not use the milk testing system was 5.6 and 4.3 when milking machine tubes were not dried. The absence of homeopathic therapy at drying-off seemed to be associated with a high probability of detecting Serratia fonticola and Serratia marcescens in milk. Indeed, the probability was 6.8-fold higher when no homeopathic therapy was given. The probability of detecting Serratia species was also higher when teat cup liners were not changed every year (odds ratio: 5.5).
A higher frequency of detection of B. linens was related to the practice of zero-grazing (6.7-fold probability of having B. linens). Arthrobacter was more commonly detected in milk from farms where the storage facility was not considered clean and those that did not rinse the milking machine with water before it was cleaned (mean odds ratios of 4.4 and 18.7, respectively).
It is important to point out that between-farm variability for the lactic acid bacteria group was low and virtually nonexistent for enterococci (p<0.001%). The within-farm variability was therefore high, which minimizes the influence of farm management practices that affect the detection of the bacterial species in question.
For the other bacterial species, no farming practices were associated with a higher probability of detection.
The distribution of the microbial species in the milk samples was independent of the geographical region in which the farm was located.
DISCUSSION
The most commonly detected microflora in the samples of goat’s milk analysed were staphylococci (54% of the milk samples and 19 of the farms). These results corroborate those of Valle et al. (1991) and Kyozaire et al. (2005). The two Serratia species, S. fonticola and S. marcescens, were also predominant microflora (52% of the milk samples and 14 of the farms). To our knowledge, no studies have yet reported the dominance of the genus Serratia in goat’s milk. S. marcescens has been detected in cow’s milk however (Ercolini et al., 2009).
Coryneform bacteria (Arthrobacter and B. linens) were also detected fairly frequently in the goat’s milk samples, especially the genus Arthrobacter (nearly 40% of the milk samples and 16 of the farms). Arthrobacter sp. have already been detected frequently in goat’s milk from one farm (Callon et al., 2007) and in cow’s milk from 26 farms (Verdier-Metz et al., 2009). B. linens is also frequently detected in cow’s milk, as shown in studies by Verdier-Metz et al. (2009): 90% of milk samples from 67 farms contained this species. To our knowledge, no studies have reported the presence of this bacterium in goat’s milk. L. lactis and E. faecalis were the two species of lactic acid bacteria most commonly present in the milk samples analysed (28% of the milk samples and 15 of the farms for L. lactis and 22% of the samples and 14 of the farms for E. faecalis). Among the lactococci, L. lactis and E. faecalis are the two species most frequently isolated from cow’s milk (Corroler et al., 1998; Dalmasso et al., 2008; Rasolofo et al., 2010), from goat’s milk (Badis et al., 2004a, b; Callon et al., 2007) and sheep’s milk (Caridi et al., 2003). The low detection frequency of enterobacteria, E. faecium and C. flavescens corroborate previous studies by other authors on goat’s milk (Callon et al., 2007) and cow’s milk (Ercolini et al., 2009; Giannino et al., 2009; Verdier-Metz et al., 2009). Thus, the species most frequently detected in milk tend to be microflora of technological importance in cheese-making. Staphylococci and corynebacteria are ripening microflora that contribute to the flavour and appearance of the cheese (Bergere and Tourneur, 1992). L. lactis is the species most frequently cited for its acidifying capacity (Casalta et al., 1995; Lafarge et al., 2004) and its involvement in the production of flavour and aromas (Corroler et al., 1998).
However, in cheese-making technology, Serratia tends to be classified among the spoilage microflora (Coiffier, 1992).
The association between farm management practices and both milk bacterial classes and the presence of individual bacterial species reveals farm management practices that differ according to bacterial class or the bacterial species detected, independently of the region.
The cleanness of the milking machine is a determining factor in the diversity of the species present in the milk. Thus, the class A microbial species profile, characterized by low bacterial species diversity in the milk (at most 3 out of the 9 species detected) and the absence of lactic acid bacteria, was significantly associated with milking machines for which all the areas inspected were clean. This result is consistent with those of Verdier-Metz et al. (2009) in cows, in which the diversity index declined when milking hygiene was improved.
The presence of Serratia in milk was significantly associated with the health of the herd (udder health and monitoring of the herd’s health). It is highly probable that the presence of Serratia in milk is partially due to contamination of the teat surface or subclinical mastitis. Indeed, previous studies have shown that S. marcescens is an opportunistic pathogen of environmental origin that is sometimes involved in subclinical mastitis in ruminants (Todhunter et al., 1991; Bergonier et al., 2003). The probability of detecting Serratia was also higher when teat cup liners are not changed annually. Worn liners are often cracked, which could favour their colonization by microflora derived from the udders.
The frequency of detection of staphylococci in milk was also significantly associated with absence of herd monitoring by the milk testing system. Staphylococci and in particular coagulase-negative staphylococci are the bacteria most commonly responsible for subclinical mastitis in goats (Bergonier et al., 2003). Subclinical mastitis is more likely to develop in herds that are not subjected to health monitoring. However, this hypothesis remains to be verified because staphylococci belong to the normal skin flora of animals and humans.
The probability of detecting B. linens more frequently was higher in zero-grazing herds, a practice that is highly correlated with continuous milk production. To our knowledge, no studies have reported this result. B. linens is widespread in nature. However, its presence has been shown in very diverse environments: soil (Crombach, 1974), bedding (Schefferle, 1966; Mohan, 1981), plants (Keddie et al., 1966; Owens and Keddie, 1969), dairy products (Abd-El-Malek and Gibson, 1952; Jayne-Williams and Skerman, 1966; Crombach, 1974) and human skin (Pitcher and Noble, 1978). The higher frequency of detection of B. linens in herds that are kept indoors may be due to a higher microbial concentration of this ubiquitous species (permanent contact with bedding, potentially greater air contamination on account of the constant presence of the animals inside the buildings, etc.). Furthermore, the milking machine is in continual contact with milk (continuous milk production for zero-grazing herds). There may therefore be greater contamination of the milking machine with B. linens from milk.
The frequency of detection of enterococci and in particular E. faecalis was associated with milkers’ standards of hygiene and to a lesser extent with animal density. Warm-blooded animals, plants and insects are the normal hosts of enterococci (Facklam et al., 2002), so it stands to reason that they should be found frequently when standards of hygiene are poor.
As for L. lactis, it was most often present when the milking parlour was in direct contact with the goats’ housing. Klijn et al. (1995) isolated strains of L. lactis from dairy farm environments (from the cattle’s udders, skin and saliva, soil near the livestock housing, feed, silage and fresh grass, so these environments could be partially responsible for introducing L. lactis to the milk. Fodder in the bedding could also be a source of contamination with lactic acid bacteria, as has been demonstrated for certain lactobacilli (Bouton et al., 2005). Thus, during milking, permanent contact with the goats’ housing would tend to favour contamination of the milk with L. lactis.
CONCLUSIONS
The predominant bacterial flora in the milk from the farms studied belonged to species of the genera Staphylococcus, Arthrobacter and Serratia. The lactic acid bacteria species L. lactis and E.faecalis were detected in about 25% of the milk samples. B. linens was detected systematically in milk from farms using zero-grazing.
The association between bacterial species and farm management practices shows that it is eventually possible to identify factors in farm management that can be used to improve control over the microbiological quality of milk intended for cheese production, independently of the region studied. The milking machine and especially its cleanness were determining factors for bacterial species diversity in the milk samples. Herd health management was also a key factor for the balance of the bacterial flora. Inferior herd health conditions and infrequent monitoring favoured contamination with S. fonticola or Staphylococcus spp.
When hygiene standards are observed, particularly those applying to milkers, the fact that direct contact between milking parlours and the goats’ housing enriches the milk in L. lactis (a lactic acid bacteria of technological interest in cheese production) is an interesting result that merits further investigation. The latter conclusion needs to be examined in more detail through molecular analysis of the L. lactis strains present in the environments close to the herd (fodder, bedding, udders, air) to evaluate their similarity with the strains present in the raw goat’s milk, for greater understanding of the sources of L. lactis. This future work would make it possible to identify the main origin of the strains of L. lactis found in the milk.
ACKNOWLEDGMENTS
The authors would like to thank the technicians and farmers from the Pelardon and Rocamadour regions for their involvement in this study and Jean-Claude Ogier for his help in the microbial analysis. This study was supported by the Midi-Pyrénées Conseil Régional and the association GALA (Janzé, France).
REFERENCES
- Abd-El-Malek, Y. and T. Gibson, 1952. Studies in the bacteriology of milk. III. The corynebacteria of milk. J. Dairy Res., 19: 153-159.
CrossRef - Allonso-Calleja, C., J. Carballo, R. Capita, A. Bernardo and M.L. Garcia-Lopez, 2002. Comparison of the acidifying activity of Lactococcus lactis subsp. lactis strains isolated from goat's milk and Valdeteja cheese. Lett. Applied Microbiol., 34: 134-138.
PubMed - Badis, A., D. Guetarni, B. Moussa-Boudjema, D.E. Henni and M. Kihal, 2004. Identification and technological properties of lactic acid bacteria isolated from raw goat milk of four Algerian races. Food Microbiol., 21: 579-588.
CrossRefDirect Link - Badis, A., D. Guetarni, B. Moussa-Boudjema, D.E. Henni, M.E. Tornadijo and M. Kihal, 2004. Identification of cultivable lactic acid bacteria isolated from Algerian raw goat's milk and evaluation of their technological properties. Food Microbiol., 21: 343-349.
CrossRef - Bergonier, D., R. de Cremoux, R. Rupp, G. Lagriffoul and X. Berthelot, 2003. Mastitis of dairy small ruminants. Vet. Res., 34: 689-716.
CrossRefDirect Link - Beuvier, E., K. Berthaud, S. Cegarra, A. Dasen, S. Pochet, S.O. Buchin and G. Duboz, 1997. Ripening and quality of Swiss-type cheese made from raw, pasteurized or microfiltered milk. Int. Dairy J., 7: 311-323.
CrossRef - Bouton, Y. and R. Grappin, 1995. Comparison of the final quality of a swiss-type cheese made from raw or microfiltered milk. Lait, 75: 31-44.
CrossRef - Buchin, S., V. Delague, G. Duboz, J.L. Berdague, E. Beuvier, S. Pochet and R. Grappin, 1998. Influence of pasteurization and fat composition of milk on the volatile compounds and flavor characteristics of a semi-hard cheese. J. Dairy Sci., 81: 3097-3108.
CrossRef - Callon, C., F. Duthoit, C. Delbes, M. Ferrand, Y. Le Frileux, R. De Cremoux and M.C. Montel, 2007. Stability of microbial communities in goat milk during a lactation year: Molecular approaches. Syst. Applied Microbiol., 30: 547-560.
PubMed - Casalta, E., Y. Vassal, M.J. Desmazeaud and F. Casabianca, 1995. Comparison of the acidifying activity of lactococcus-lactis isolated from Corsican goat milk and cheese. Food Sci. Technol. Lebensmittel Wissenschaft Technol., 28: 291-299.
CrossRef - Corroler, D., I. Mangin, N. Desmasures and M. Gueguen, 1998. An ecological study of lactococci isolated from raw milk in the camembert cheese registered designation of origin area. Applied Environ. Microbiol., 64: 4729-4735.
Direct Link - Crombach, W.H., 1974. Relationships among coryneform bacteria from soil cheese and sea fish. Antonie Van Leeuwenhoek, 40: 347-359.
CrossRef - Dalmasso, M., S. Prestoz, V. Rigobello and Y. Demarigny, 2008. Behavior of Lactococcus lactis subsp lactis biovar. diacetylactis in a four Lactococcus strain starter during successive milk cultures. Food Sci. Technol. Int., 14: 469-477.
CrossRef - Delos Reyes-Gavilan, C.G., G.K.Y. Limsowtin, P. Tailliez, L. Sechaud and J.P. Accolas, 1992. A lactobacillus-helveticus-specific DNA probe detects restriction-fragment-length-polymorphisms in this species. Applied Environ. Microbiol., 58: 3429-3432.
Direct Link - El-Gazzar, F.E., H.F. Bohner and E.H. Marth, 1992. Antagonism between listeria-monocytogenes and lactococci during fermentation of products from ultrafiltered skim milk. J. Dairy Sci., 75: 43-50.
PubMed - Ercolini, D., F. Russo, I. Ferrocino and F. Villani, 2009. Molecular identification of mesophilic and psychrotrophic bacteria from raw cows milk. Food Microbiol., 26: 228-231.
CrossRef - Foschino, R., A. Invernizzi, R. Barucco and K. Stradiotto, 2002. Microbial composition, including the incidence of pathogens, of goat milk from the Bergamo region of Italy during a lactation year. J. Dairy Res., 69: 213-225.
Direct Link - Gay, M. and A. Amgar, 2005. Factors moderating Listeria monocytogenes growth in raw milk and in soft cheese made from raw milk. Lait, 85: 153-170.
CrossRefDirect Link - Giannino, M.L., M. Marzotto, F. Dellaglio and M. Feligini, 2009. Study of microbial diversity in raw milk and fresh curd used for Fontina cheese production by culture-independent methods. Int. J. Food Microbiol., 130: 188-195.
CrossRefPubMedDirect Link - Jayne-Williams, D.J. and T.M. Skerman, 1966. Comparative studies on coryneform bacteria from milk and dairy sources. J. Applied Bacteriol., 29: 72-92.
CrossRef - Keddie, R.M., B.G.S. Leask and J.M. Grainger, 1966. A comparison of coryneform bacteria from soil and herbage: Cell wall composition and nutrition. J. Applied Bacteriol., 29: 17-43.
CrossRef - Klijn, N., A.H. Weerkamp and W.M. de Vos, 1995. Detection and characterization of lactose-utilizing Lactococcus spp. in natural ecosystems. Applied Environ. Microbiol., 61: 788-792.
Direct Link - Kyozaire, J.K., C.M. Veary, I.M. Petzer and E.F. Donkin, 2005. Microbiological quality of goat's milk obtained under different production systems. J. S. Afr. Vet. Assoc., 76: 69-73.
PubMedDirect Link - Lafarge, V., J.C. Ogier, V. Girard, V. Maladen, J.Y. Leveau, A. Gruss and A. Delacroix-Buchet, 2004. Raw cow milk bacterial population shifts attributable to refrigeration. Applied Environ. Microbiol., 70: 5644-5650.
CrossRef - Ogier, J.C., O. Son, A. Gruss, P. Tailliez and A. Delacroix-Buchet, 2002. Identification of the bacterial microflora in dairy products by temporal temperature gradient gel electrophoresis. Applied Environ. Microbiol., 68: 3691-3701.
CrossRefDirect Link - Owens, J.D. and R.M. Keddie, 1969. The nitrogen nutrition of soil and herbage coryneform bacteria. J. Applied Bacteriol., 32: 338-347.
CrossRef - Rasolofo, E.A., D. St-Gelais, G. La Pointe and D. Roy, 2010. Molecular analysis of bacterial population structure and dynamics during cold storage of untreated and treated milk. Int. J. Food Microbiol., 138: 108-118.
CrossRef - Schefferle, H.E., 1966. Coryneform bacteria in poultry deep litter. J. Applied Bacteriol., 29: 147-160.
Direct Link - Todhunter, D.A., K.L. Smith and J.S. Hogan, 1991. Serratia species isolated from bovine intramammary infections. J. Dairy Sci., 74: 1860-1865.
PubMed - Valle, J., S. Piriz, R. De La Fuente and S. Vadillo, 1991. Staphylococci isolated from healthy goats. Zentralbl Veterinarmed B., 38: 81-89.
PubMed - Verdier-Metz, I., V. Michel, C. Delbes and M.C. Montel, 2009. Do milking practices influence the bacterial diversity of raw milk. Food Microbiol., 26: 305-310.
CrossRef - Caridi, A., P. Micari, P. Caparra, A. Cufari and V. Sarullo, 2003. Ripening and seasonal changes in microbial groups and in physico-chemical properties of the ewes cheese Pecorino del Poro. Int. Dairy J., 13: 191-200.
CrossRef