
A.F. Park
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
J.E. Shirley
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
E.C. Titgemeyer
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
R.C. Cochran
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
J.M. DeFrain
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
E.E. Wickersham
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
D.E. Johnson
Department of Statistics, Kansas State University, Manhattan, KS, USA
International Journal of Dairy Science
Year: 2010 | Volume: 5 | Issue: 4 | Page No.: 253-263







ABSTRACT
Present objective was to characterize plasma metabolites as cows transitioned from non-lactational to lactational state. Four multiparous, pregnant Holstein cows were used to achieve this goal. Plasma was obtained on day 79 before projected calving, weekly thereafter until parturition and on days 1, 3, 5, 7, 15, 20, 25, 30, 60 and 90 postpartum. Crude protein concentrations were 18.7, 11.5, 15.6 and 18.4% for late lactation (-79, -72, -65 days prepartum), far-off dry (-58, -51, -44, -37, -30 days prepartum), close-up dry (-23, -16, -9, -2 days prepartum) and early lactation diets. Albumin was above 3 g dL-1 during the last 3 weeks of lactation (days -79, -72, -65) and the first 3 weeks of the far-off dry period (days -58, -51, -44), decreased to less than 2 g dL-1 during the close-up dry period, then increased linearly after parturition reaching over 4 g dL-1 by day 25 postpartum. Plasma non-esterified fatty acids were relatively constant prepartum, increased at parturition peaking on day 15 and returned to prepartum levels by day 30 postpartum. Insulin and glucagon tended to decrease prior to calving, but glucagon increased during the first week postpartum, whereas insulin continued to decline. Insulin: glucagon was relatively constant prepartum and declined sharply through the first week after calving. Glucose and triacylglycerol decreased following parturition. Plasma urea varied with level of dietary protein and dry matter intake. Plasma total α-amino-nitrogen increased from parturition through day 20 of lactation. These data support the concept that plasma metabolites respond to changes in diet and lactational status.
PDF Abstract XML References Citation
Received: March 03, 2010;
Accepted: May 22, 2010;
Published: August 05, 2010
How to cite this article
A.F. Park, J.E. Shirley, E.C. Titgemeyer, R.C. Cochran, J.M. DeFrain, E.E. Wickersham and D.E. Johnson, 2010. Characterization of Plasma Metabolites in Holstein Dairy Cows during the Periparturient Period. International Journal of Dairy Science, 5: 253-263.
DOI: 10.3923/ijds.2010.253.263
URL: https://scialert.net/abstract/?doi=ijds.2010.253.263
DOI: 10.3923/ijds.2010.253.263
URL: https://scialert.net/abstract/?doi=ijds.2010.253.263
INTRODUCTION
The high producing dairy cow prepares for parturition and lactogenesis with surges in plasma insulin (Smith et al., 2008), non-esterified fatty acids (NEFA) and glucose as well as depressions in progesterone (Kunz et al., 1985), blood calcium and immune status (Goff and Horst, 1997). The trigger for these metabolic changes appears to be fluctuations between nutrient supply and demand. Initially, nutrient demand comprises only maintenance and fetal growth requirements (Moe and Tyrrell, 1972), but later it includes support for lactogenesis as the cow prepares for lactation (Capuco et al., 1997) in the face of a nutrient supply that typically diminishes during the last 3 weeks of gestation (Bertics et al., 1992). The degree to which metabolic changes occur in the dry period may influence the incidence of metabolic disease (Curtis et al., 1985). Excessive NEFA concentrations lead to fatty liver and ketosis (Bertics et al., 1992) and low levels of blood calcium can lead to milk fever (Goff and Horst, 1997). Thus, dry cow nutritional and managerial practices are necessary to control metabolic disease and optimize transition into lactation. For example, overfeeding cows in the late prepartum period increases insulin resistance and leads to greater NEFA mobilization, lower dry matter intake and greater incidences of fatty liver and ketosis (Holtenius et al., 2003).
Most dairy cows are allowed a 45 to 60-day dry or non-lactating period at the end of 305-day lactations (Jordan and Fourdraine, 1993). Bachman (2002) and Gulay et al. (2003) challenged this concept of a 60-day dry period by showing little advantage in the subsequent lactation to dry periods longer than 30 to 40 days. Others investigated eliminating the dry period, with few negative effects for mature Holstein cows (Annen et al., 2003; Rastani et al., 2005). Holcomb et al. (2001) and Winkelman et al. (2008) reported positive responses with limit-feeding dry cows nearing the end of their gestation. With these various management styles, it is necessary to understand how dairy cows adapt to nutritional and managerial changes, not only through the close-up period, but also from one lactation into the next, in order to evaluate dry-cow management strategies.
The purpose of this study was to characterize changes in selected endocrine and metabolic parameters over time as Holstein dairy cows fed a traditional two-group dry-cow feeding program moved from late gestation into early lactation. Changes in plasma metabolites were used to reflect metabolic shifts.
MATERIALS AND METHODS
Cows and Treatments
The Kansas State University Institutional Animal Care and Use Committee approved all practices in 2000. Cows were fed, housed and managed as described by Park et al. (2010). In brief, four multiparous, pregnant Holstein cows with similar parities, calving dates, body weights, body condition scores, previous 305-days mature equivalent milk yields and frame sizes and bred to bulls with similar calving ease scores were selected for this study. A late lactation diet was fed until 60 days before expected calving date. A far-off dry diet (day 60 through day 29 prepartum) and a close-up dry diet (day 28 prepartum until calving) were fed during the dry period. Cows were fed an early lactation diet from the first feeding after calving through 90 days in milk.
Sampling and Analysis
Blood samples were collected from the coccygeal vein weekly during late lactation, far-off dry and close-up dry periods and on days 1, 3, 5, 7, 15, 20, 25, 30, 60 and 90 postpartum. Prepartum samples were designated in relation to actual calving date (±2 days). Blood was sampled 3 h after feeding into EDTA-containing Vacutainer tubes (Becton, Dickinson and Company, Franklin Lakes, NJ), which were placed in an ice bath until processed. Tubes were centrifuged at 500 x g for 20 min, plasma was removed and 3-mL aliquots were stored frozen at -20°C until analysis.
Laboratory Analyses
Plasma albumin was measured using bromcresol green binding (Dumas et al., 1971) using a commercial kit (No. 631, Sigma Diagnostics, St. Louis, MO). Triacylglycerol (TAG) analysis (Sigma Diagnostics No. 343) utilized the procedure from Wako Chemicals (code No. 997-69801) with modifications from McGowan et al. (1983) and Fossati and Lorenzo (1982). Plasma glucose, urea-N (PUN) and total α-amino-nitrogen (TAAN) analyses used a Technicon auto analyzer II (Technicon Industrial Systems, Tarrytown, NY). The method of Gochman and Schmitz (1972) was used to determine plasma glucose. Measurement of PUN involved a diacetyl-monozime assay (Marsh et al., 1965). Determination of TAAN used the trinitrobenzenesulfonic acid assay (Palmer and Peters, 1969). Plasma NEFA were measured as described by Eisemann et al. (1988) (Wako Chemicals, procedure No. 994-75409E). Insulin was measured with a RIA kit (DSL-1600, Diagnostics Systems Laboratory, Webster, TX); the intraassay CV was 10.7% and the sensitivity was 0.0154 ng mL-1. Glucagon was measured with a RIA kit (Double Antibody Glucagon, Diagnostics Products Corporation, Los Angeles, CA); the intraassay CV was 8.9% and the sensitivity was 17.2 pg mL-1.
Statistical Analysis
The trial was a randomized design with six specified periods: late lactation (days -79, -72 and -65), far-off dry (days -58, -51, -44, -37 and -30), close-up dry (days -23, -16, -9 and -2), fresh cow (days 1, 3, 5 and 7), early lactation (days 15, 20, 25, 30) and peak lactation (days 60 and 90). Each of the 22 blood sampling dates was considered as a treatment. Data were analyzed by analysis of variance using the repeated option in the mixed procedure of SAS (SAS Inst. Inc., Cary, NC) with period and day within period in the model and with day as the repeated measure for each cow. The spatial powers option was used for the covariance structure. The covariance structure was based on the Akaike’s information criterion closest to zero. Periods were compared using pair-wise t-tests. Linear, quadratic and cubic contrasts were used to evaluate effects of time within each of the periods. One outlier was removed from the data set for NEFA during the late lactation period on day -72 leaving a mean of 0.23 mmol L-1; the original mean was 0.42 mmol L-1 with an quadratic effect of day within this period (p<0.05). Data are presented as least square means with pooled SEM.
RESULTS
Plasma glucose (Table 1) remained unchanged through late lactation and through the dry period, but decreased (p<0.05) during the first week after calving and then increased (p<0.05) between 15 and 30 days in milk. There was a numeric increase (linear effect in close-up dry period, p=0.15) in plasma glucose in the last sample collected prior to parturition.
Plasma insulin (Table 1) remained unchanged through late lactation and the dry period and exhibited a tendency (quadratic effect in close-up dry period, p=0.10) to decline at 2 days prior to calving. Plasma insulin concentrations were lower (p<0.05) after calving than before, as a result of a drop following calving as well as a tendency to decrease during the fresh cow period (linear, p=0.15) and remained lower (p<0.05) than in the dry period throughout the first 90 days in milk.
Plasma glucagon (Table 1) remained unchanged over late lactation and through the dry periods, but tended to decrease (linear, p=0.07) during the close-up dry period. Plasma glucagon increased linearly over the fresh cow period and then remained at concentrations greater than in the dry periods for the initial 90 days in milk.
As a result of the changes discussed for insulin and glucagon, molar ratios of insulin: glucagon in plasma were greater during the close-up dry period than during late lactation, but then decreased dramatically at parturition (p<0.05) and continued to decline further during the first week of lactation. Ratios of insulin:glucagon then remained low in plasma throughout the initial 90 days in milk.
| Table 1: | Plasma concentrations of glucose, insulin and glucagon in dairy cattle as impacted by period and day relative to parturition |
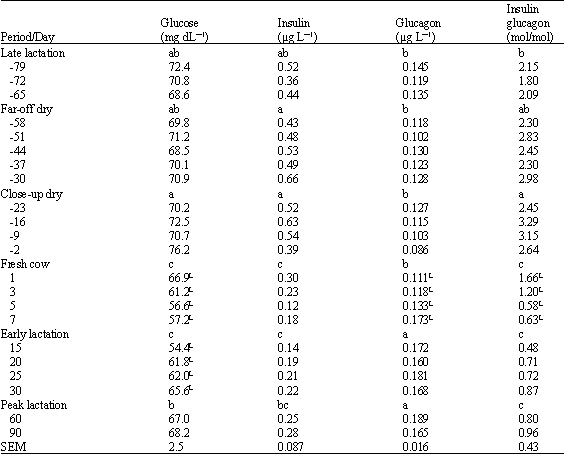 | |
| a,b,c: Periods without a common letter differ (p<0.05). L Linear effect of day within period (p<0.05) | |
Plasma NEFA concentrations (Table 2) remained unchanged over the late lactation and dry periods, but increased across parturition (p<0.05). Plasma NEFA reached their greatest concentration by day 15, then decreased linearly (p<0.05) between 15 and 30 days in milk and during peak lactation were similar to concentrations observed during the dry periods.
Plasma TAG concentrations remained relatively unchanged over the late lactation and dry periods, although some modest increases were observed 9 days prior to parturition (cubic effect in the close-up dry period, p<0.05). Concentrations of plasma TAG were decreased after calving and tended to remain lower than during the close-up dry period throughout the first 90 days in milk. During 15 to 30 days in milk, cubic effects of time were significant (slight increases on day 20 followed by a slight decrease on day 25).
Albumin concentrations (Table 2) decreased during the far-off dry period between 44 and 30 days prior to parturition (linear and quadratic, p<0.05) and remained lower throughout the close-up dry period. Following parturition, plasma albumin increased linearly during the first week of lactation and was greater during early and peak lactation than during the fresh cow period. Plasma albumin concentrations during early and peak lactation were somewhat greater than observed during late lactation (p<0.05).
Concentrations of TAAN were not different during the dry periods when compared to late lactation, although cubic effects of time were observed during the far-off dry period due to somewhat lower values observed on days -51 and -30. During the fresh cow period and during the early lactation period, plasma concentrations of TAAN exhibited increases such that concentrations observed during early lactation were greater than those observed during the dry periods or during the first week of lactation.
| Table 2: | Plasma concentrations of non-esterified fatty acids (NEFA), triacylglycerol, albumin, total α-amino N and urea in dairy cattle as impacted by period and day relative to parturition |
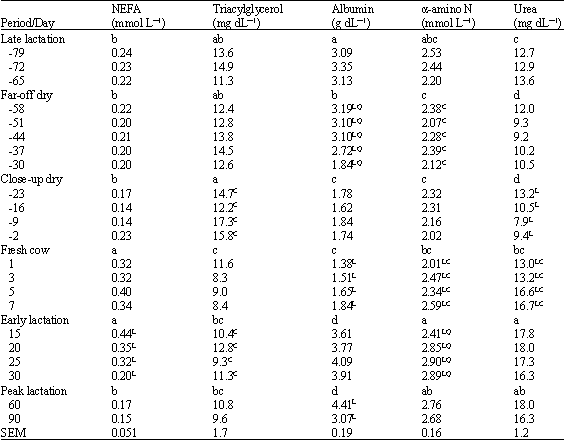 | |
| a,b,c,d: Periods without a common letter differ (p<0.05). L Linear, Q Quadratic, or CCubic effect of day within period (p<0.05) | |
Concentrations of TAAN during peak lactation were greater than observed during the dry periods.
Concentrations of PUN (Table 2) were lower during the far-off dry period than during late lactation and demonstrated linear (p<0.05) decreases during the close-up dry period. Concentrations of PUN were greater after calving than during the dry periods and increased between 3 and 5 days after parturition (linear and cubic effects during the fresh cow period; p<0.05). Additionally, PUN was greater during early lactation than during the fresh cow period.
DISCUSSION
Transition from Late Lactation to the Dry Period
There have been tremendous research efforts focused on the transition dairy cow, yet few studies have documented changes as cows move from late lactation into early lactation. In our study, few large changes in plasma metabolites occurred as cows moved from late lactation to the far-off period. Annen et al. (2003) and Rastani et al. (2005) suggested eliminating the dry period in an effort to alleviate stress on cows as they move from lactation to a non-lactating state. Stress events typically are characterized with a depression in insulin combined with an increase in glucagon that leads to higher plasma glucose, NEFA and TAG concentrations. However, our study showed that concentrations of insulin, glucagon, glucose, NEFA and TAG remained similar in late lactation, across dry-off and into the far-off period. These relatively stable plasma metabolite concentrations suggest that little stress occurred as the cows moved from late lactation into and through the far-off period.
Dann et al. (2006) touted the importance of far-off period nutrition on subsequent lactational performance. The relative stability of insulin, glucagon, glucose, NEFA and TAG from late lactation and into the far-off dry period, however, indicates that energy requirements were unlikely to be experiencing large changes through the far-off period.
Plasma albumin concentrations decreased during the far-off period, showing a large decline between day -37 and day -30. Lower albumin concentrations may reflect an increase in protein demand (Moorby et al., 2000). Concentrations of PUN were lower in the far-off dry period than during late lactation, reflecting the lower crude protein concentration of the far-off diet. Concentrations of TAAN also demonstrated some lower values during the far-off dry period (cubic effect, with TAAN being notably lower on day -51 and day -30), likely reflecting the lower crude protein content of the diet and lesser supplies of metabolizable protein to the cow.
Transition from Far-off Dry to Calving
Concentrations of glucose, insulin and glucagon were unaffected by the change from the far-off diet to the close-up diet. Concentrations of TAG on average were similar with the feeding of the close-up and far-off diets, although during the close-up period there was more variation among sampling days (cubic effect). Dietary fat level was similar between the two diets which matches the similarity in plasma TAG concentrations between the far-off and close-up dry periods, although the close-up diet contained a greater concentration of net energy for lactation and nonfiber carbohydrates. Greater concentrations of net energy for lactation and nonfiber carbohydrates are commonly included in close-up dairy diets to counter-balance the depression in intake typically observed in prepartum cows (Bertics et al., 1992) and the increase in nutrient requirements for fetal (Moe and Tyrrell, 1972) and mammary (Capuco et al., 1997) growth. The higher energy concentration in the close-up diet than in the far-off diet may have contributed to the numerically lower NEFA concentrations during the initial stages (day -23 through day -9) of the close-up period.
Plasma NEFA concentrations started to increase by day -2. Inadequate dietary nutrient supply during the periparturient period can cause tissue mobilization (Moe et al., 1971) or low lactational performance (Hart et al., 1979). Clearly, cows in our study began to mobilize tissue prepartum by 2 days prepartum, as indicated by numeric increases in NEFA and PUN by day -2. Cows with relatively high condition scores prepartum can experience a decreasing energy balance from approximately 4 weeks before parturition until parturition, possibly due to reduced intake (Park et al., 2000), but cows used in our study were in acceptable body condition score (2.75). A further understanding of how cow type (thin, fat, heifer, aged cow and cow carrying twins) and nutritional strategy might impact the initiation of body tissue mobilization as the cows approach parturition would be instrumental in determining which management and feeding strategies would be optimal.
Plasma glucose concentrations are controlled by insulin and glucagon concentrations and insulin concentrations started their lactation-associated decrease by day -2. Insulin resistance also may play a part in increased tissue mobilization leading to the rise in NEFA levels (Bell, 1995). Smith et al. (2008) investigated dietary nutritional modifications to alleviate the prepartum insulin resistance, but they were unable to show that feeding a diet with lower concentrations of nonfiber carbohydrates and fat affected the initiation or degree of prepartum insulin resistance.
The increase in PUN between day -30 (far-off diet) and day -23 (close-up diet) may be attributed to the greater dietary crude protein concentration for the close-up diet (Park et al., 2010). The declines in PUN over the close-up dry period probably reflect several changes occurring simultaneously. Plasma urea-N is a reflection of dietary protein concentration, dry matter intake and dietary carbohydrate level and availability, as well as protein mobilization from or deposition in body tissues (National Research Council, 2001). Initial declines in PUN may reflect reductions in dry matter intake (Park et al., 2010), whereas the slight increase between day -9 and -2 may reflect increased mobilization of body tissues in support of the initiation of lactation. Dry matter intake typically decreases during the last 2 to 3 weeks of gestation (Bertics et al., 1992), which in our study was related to the decrease in PUN between day -23 and -9. Plasma urea-N increased numerically on day -2 when intake was low, suggesting increased tissue protein mobilization (Butler, 1998). The timing of this increased tissue mobilization may be pivotal in allowing cows to have a good transition to lactation because most cows experience peak NEFA concentrations at parturition (Vazquez-Anon et al., 1994) when various aspects of the dairy cow’s life are changing (grouping, ration and social structure). Altering the timing of peak tissue mobilization might smooth transition into lactation.
Numeric decreases in TAAN during the close-up period (linear, p=0.16) likely relate to increases in protein (amino acid) demand to support mammary gland turnover that usually begins approximately 3 to 4 weeks prior to parturition (Capuco et al., 1997). In addition, this decrease could be due to fetal use of glucose, amino acids and glycerol (Bell, 1995), which increases linearly during late gestation (Bell et al., 1995). Others have observed similar results and concluded that the majority of metabolic adjustments prior to calving occur in the last 2 to 4 days of gestation (Athanasiou and Phillips, 1978; Bauman and Currie, 1980; Kunz et al., 1985).
Cows may have begun to mobilize lean tissue before day -2, as demonstrated by lower albumin concentrations by 30 days prior to calving. Moorby et al. (2000) reported that a lower albumin concentration during the last 3 to 4 weeks of gestation was due to a repartitioning of labile protein stores to the mammary gland in preparation for lactation, although decreased hepatic synthesis during this period could also contribute to the low plasma concentrations of albumin. Albumin concentrations remained low until parturition and for the first 7 days of lactation when they might have been used to provide amino acids for milk production when nutrient intake was insufficient to support lactation (Bell, 1995). More research on labile protein stores during this physiological state of the dairy cow is needed to substantiate these observations.
Tissue mobilization typically begins prepartum with the commonly observed intake depression during the last 3 weeks of gestation (Bertics et al., 1992) at the same time nutrient requirements increase (National Research Council, 2001). It has been hypothesized that cows with less condition will sustain intake longer and experience an abrupt drop in dry matter intake just prior to calving (Hayirli et al., 2002). This concept has been confirmed by those investigating limit feeding during the dry period (Holcomb et al., 2001). Body condition of cows in our study (average of 2.94) was less than recommended (Wildman et al., 1982), which may have led to tissue mobilization beginning near 2 days prepartum instead of earlier in the close-up period. Calculated energy balance data (Park et al., 2010) showed that cows were not in a negative energy balance prepartum, supporting this theory. A further understanding of how the nutritional strategy affects the timing of tissue mobilization would be valuable in optimizing nutritional strategies for different types of cows.
Transition into Lactation
Tissue mobilization is usually most active from 14 days prepartum to 35 days postpartum (Komaragiri et al., 1998) and Ben Salem and Bouraoui (2008) demonstrated the importance of appropriate nutrition during this period to minimize loss of body condition. Our cows were calculated (Park et al., 2010) to be in a negative energy balance postpartum until day 34 after calving. Concentrations of PUN in our cows help to confirm this tissue mobilization with PUN concentrations increasing during the first 7 days of lactation and plateauing by day 15. Lactating cows were fed a diet higher in crude protein than that fed prior to calving and they experienced greater intake on the lactation diet, which may explain some of the greater PUN concentrations. However, intake did not peak until day 81 of lactation (Park et al., 2010), whereas PUN values plateaued by day 15, possibly indicating that a portion of the increase was due to tissue mobilization (Butler, 1998). Similar to PUN, TAAN increased during the fresh cow period with concentrations plateauing near day 20. Traditionally, dairy cows have been thought to mobilize tissue until approximately 60 days in milk (McNamara et al., 1995). The low body condition of our cows may help explain the discrepancy, because cows fed prepartum for greater condition (3.7 vs. 2.7) experienced a negative energy balance longer into lactation (56 vs. 28 days; Agenas et al., 2003).
Plasma NEFA concentrations peaked on day 15 postpartum, which contrasts other studies where peaks were observed around 1 day postpartum (Bertics et al., 1992; Vazquez-Anon et al., 1994; Grum et al., 1996). During this same time frame, we observed an increase in glucagon and a numeric decrease in insulin resulting in linear decreases in insulin: glucagon. These changes should support increased lipolysis as demonstrated by maintenance of elevated plasma NEFA levels over this period. In addition, the increases in glucagon should support increased glycogenolysis and gluconeogensis; linear decreases in blood glucose levels in the face of ostensible increases in glucose supply likely reflect increased nutrient use by the mammary gland for milk production. High concentrations of plasma NEFA may lead to fat infiltration in the liver and subsequent metabolic diseases like fatty liver and ketosis, which can cause cows to reduce feed intake and milk production (National Research Council, 2001). Nazifi et al. (2008) found higher plasma NEFA in cows experiencing subclinical ketosis during the first 2 months of lactation. Between 6 and 20 days in milk, our cows demonstrated increases in intake from 16.4 to 20.5 kg day-1 and in milk production from 23.4 to 35.3 kg day-1. These responses suggest that a later than expected peak in plasma NEFA is not exceedingly detrimental to cow health. In support of this concept, plasma TAG concentrations decreased after calving, as expected, with TAG generally remaining low through early lactation. The low concentrations of TAG early in lactation likely was due to effective use of TAG for milk production.
Holcomb et al. (2001) observed that peak NEFA concentrations occurred later in lactation when cows (prepartum body condition score near 3.5) were restrictedly fed during the last 28 days of the dry period than when they were allowed ad libitum access to feed; the restrictedly fed cows with later NEFA peaks performed as well as those cows fed for ad libitum intake. This observation along with results from our study indicate that cows calving with less body condition may maintain intake longer prepartum and peak in NEFA concentration later in lactation (day 15 versus day 1), potentially leading to an improvement in cow health.
CONCLUSIONS
Dairy cows normally mobilize tissue as they enter lactation. Using a typical two-group dry period feeding strategy, few metabolic disturbances were noted as cows moved from late lactation into the far-off and finally the close-up period. A number of metabolite changes occurred near 2 days prepartum with increases in plasma NEFA and urea-N reflecting increases in tissue mobilization. Additional metabolite changes continued after parturition with decreases in insulin:glucagon and glucose and increases in PUN and TAAN through the first week of lactation. Further research is warranted on the impact of different feeding schemes during the dry period to prepare dairy cows for lactation.
ACKNOWLEDGMENTS
This study is contribution No. 09-274-J from the Kansas Agricultural Experiment Station, Manhattan, KS, USA. Partial funding for the experiment came from the Kansas Agricultural Experiment Station.
REFERENCES
- Agenas, S., E. Burstedt and K. Holtenius, 2003. Effects of feeding intensity during the dry period. 1. feed intake, body weight and milk production. J. Dairy Sci., 86: 870-882.
Direct Link - Athanasiou, V.N. and R.W. Phillips, 1978. Stability of plasma metabolites and hormones in parturient dairy cows. Am. J. Vet. Res., 39: 953-956.
PubMed - Bachman, K.C., 2002. Milk production of dairy cows treated with estrogen at the onset of a short dry period. J. Dairy Sci., 85: 797-803.
Direct Link - Bauman, D.E. and W.B. Currie, 1980. Partitioning of nutrients during pregnancy and lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci., 63: 1514-1529.
CrossRef - Bell, A.W., 1995. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci., 73: 2804-2819.
PubMedDirect Link - Bell, A.W., R. Slepetis and R.A. Ehrhardt, 1995. Growth and accretion of energy and protein in the gravid uterus during late pregnancy in Holstein cows. J. Dairy Sci., 78: 1954-1961.
CrossRef - Salem, M.B. and R. Bouraoui, 2008. Effects of calcium salts of palm fatty acids and protected methionine supplementation on milk production and composition and reproductive performances of early lactation dairy cows. Int. J. Dairy. Sci., 3: 187-193.
CrossRefDirect Link - Bertics, S.J., R.R. Grummer, C. Cadorniga-Valino and E.E. Stoddard, 1992. Effect of prepartum dry matter intake on liver triglyceride concentration and early lactation. J. Dairy Sci., 75: 1914-1922.
CrossRefPubMedDirect Link - Butler, W.R., 1998. Review: Effect of protein nutrition on ovarian and uterine physiology in dairy cattle. J. Dairy Sci., 81: 2533-2539.
CrossRefDirect Link - Capuco, A.V., R.M. Akers and J.J. Smith, 1997. Mammary growth in Holstein cows during the dry period: Quantification of nucleic acids and histology. J. Dairy Sci., 80: 477-487.
Direct Link - Curtis, C.R., H.N. Erb, C.J. Sniffen, R.D. Smith and D.S. Kronfeld, 1985. Path analysis of dry period nutrition, postpartum metabolic and reproductive disorders and mastitis in Holstein cows. J. Dairy Sci., 68: 2347-2360.
Direct Link - Dann, H.M., N.B. Litherland, J.P. Underwood, M. Bionaz, A. D`Angelo, J.W. McFadden and J.K. Drackley, 2006. Diets during the far-off and close-up dry periods affect periparturient metabolism and lactation in multiparous cows. J. Dairy Sci., 89: 3563-3577.
Direct Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Eisemann, J.H., G.B. Huntington and C.L. Ferrell, 1988. Effects of dietary clenbuterol on metabolism of the hindquarters in steers. J. Anim. Sci., 66: 342-353.
Direct Link - Fossati, P. and L. Prencipe, 1982. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem., 28: 2077-2080.
CrossRefPubMedDirect Link - Goff, J.P. and R.L. Horst, 1997. Physiological changes at parturition and their relationship to metabolic disorders. J. Dairy Sci., 80: 1260-1268.
CrossRefPubMedDirect Link - Gochman, N. and J. Schmitz, 1972. Application of a new peroxide indicator reaction to the specific, automated determination of glucose with glucose oxidase. Clin. Chem., 18: 943-950.
PubMed - Grum, D.E., J.K. Drackley, R.S. Younker, D.W. LaCount and J.J. Veenhuizen, 1996. Nutrition during the dry period and hepatic lipid metabolism of periparturient dairy cows. J. Dairy Sci., 79: 1850-1864.
CrossRefDirect Link - Gulay, M.S., M.J. Hayen, K.C. Bachman, T. Belloso, M. Liboni and H.H. Head, 2003. Milk production and feed intake of holstein cows given short (30-d) or normal (60-d) dry periods. J. Dairy Sci., 86: 2030-2038.
Direct Link - Hart, I.C., J.A. Bines and S.V. Morant, 1979. Endocrine control of energy metabolism in the cow: Correlations of hormones and metabolites in high and low yielding cows for stages of lactation. J. Dairy Sci., 62: 270-277.
PubMed - Hayirli, A., R.R. Grummer, E.V. Noedheim and P.M. Crump, 2002. Animal and dietary factors affecting feed intake during the prefresh transition period in Holsteins. J. Dairy Sci., 85: 3430-3443.
CrossRefDirect Link - Holcomb, C.S., H.H. van Horn, H.H. Head, M.B. Hall and C.J. Wilcox, 2001. Effects of prepartum dry matter intake and forage percentage on postpartum performance of lactating dairy cows. J. Dairy Sci., 84: 2051-2058.
Direct Link - Holtenius, K., S. Agenas, C. Delavaud and Y. Chilliard, 2003. Effects of Feeding intensity during the dry period. 2. Metabolic and hormonal responses. J. Dairy Sci., 86: 883-891.
Direct Link - Komaragiri, M.V., D.P. Casper and R.A. Erdman, 1998. Factors affecting body tissue mobilization in early lactation of body fat and protein. J. Dairy Sci., 81: 169-175.
Direct Link - Kunz, P.L., J.W. Blum, I.C. Hart, H. Bickel and J. Landis, 1985. Effects of different energy intakes before and after calving on food intake, performance and blood hormones and metabolites in dairy cows. Anim. Prod., 40: 219-231.
Direct Link - Marsh, W.H., B. Fingerhut and H. Miller, 1965. Automated and manual direct methods for the determination of blood urea. Clin. Chem., 11: 624-627.
Direct Link - McGowan, M.W., J.D. Artiss, D.R. Strandbergh and B. Zak, 1983. A peroxidase-coupled method for the colorimetric determination of serum triglycerides. Clin. Chem., 29: 538-542.
PubMedDirect Link - McNamara, J.P., J.H. Harrison, R.L. Kincaid and S.S. Waltner, 1995. Lipid metabolism in adipose tissue of cows fed high fat diets during lactation. J. Dairy Sci., 78: 2782-2796.
Direct Link - Moe, P.W., H.F. Tyrrell and W.P. Flatt, 1971. Energetics of body tissue mobilization. J. Dairy Sci., 54: 548-553.
Direct Link - Moe, P.W. and H.F. Tyrrell, 1972. Metabolizable energy requirements of pregnant dairy cows. J. Dairy Sci., 55: 480-483.
Direct Link - Moorby, J.M., R.J. Dewhurst, J.K.S. Tweed, M.S. Dhanoa and N.F.G. Beck, 2000. Effects of altering the energy and protein supply to dairy cows during the dry period. 2. Metabolic and hormonal responses. J. Dairy Sci., 83: 1795-1805.
Direct Link - NRC, 2001. Nutrient Requirements of Dairy Cattle. 7th Edn., National Academy Press, Washington, DC, ISBN: 978-0-309-06997-7, Pages: 405.
CrossRefDirect Link - Palmer, D.W. and T. Peters Jr., 1969. Automated determination of free amino groups in serum and plasma using 2,4,6-trinitrobenzene sulfonate. Clin. Chem., 15: 891-901.
PubMedDirect Link - Rastani, R.R., R.R. Grummer, S.J. Bertics, A. Gumen, M.C. Wiltbank, D.G. Mashek and M.C. Schwab, 2005. Reducing dry period length to simplify feeding transition cows: Milk production, energy balance and metabolic profiles. J. Dairy Sci., 88: 1004-1014.
Direct Link - Smith, K.L., M.R. Waldron, L.C. Ruzzi, J.K. Drackley, M.T. Socha and T.R. Overton, 2008. Metabolism of dairy cows as affected by prepartum dietary carbohydrate source and supplementation with chromium throughout the periparturient period. J. Dairy Sci., 91: 2011-2020.
CrossRefDirect Link - Vazquez-Anon, M., S. Bertics, M. Luck and R.R. Grummer, 1994. Peripartum liver triglyceride and plasma metabolites in dairy cows. J. Dairy Sci., 77: 1521-1528.
CrossRef - Wildman, E.E., G.M. Jones, P.E. Wagner, R.L. Boman, H.F. Troutt Jr. and T.N. Lesch, 1982. A dairy cow body condition scoring system and its relationship to selected production characteristics. J. Dairy Sci., 65: 495-501.
CrossRefDirect Link - Winkelman, L.A., T.H. Elsasser and C.K. Reynolds, 2008. Limit-feeding a high-energy diet to meet energy requirements in the dry period alters plasma metabolite concentrations but does not affect intake or milk production in early lactation. J. Dairy Sci., 91: 1067-1079.
CrossRef