
Dalmasso Marion
Laboratoire de Microbiologie Alimentaire, ISARA-Lyon, Agrapole, 23, rue Jean Baldassini, 69364 Lyon Cedex 07, France
Prestoz Sylvie
Laboratoire de Microbiologie Alimentaire, ISARA-Lyon, Agrapole, 23, rue Jean Baldassini, 69364 Lyon Cedex 07, France
Rigobello Veronique
Laboratoire de Microbiologie Alimentaire, ISARA-Lyon, Agrapole, 23, rue Jean Baldassini, 69364 Lyon Cedex 07, France
Demarigny Yann
Laboratoire de Microbiologie Alimentaire, ISARA-Lyon, Agrapole, 23, rue Jean Baldassini, 69364 Lyon Cedex 07, France
International Journal of Dairy Science
Year: 2008 | Volume: 3 | Issue: 3 | Page No.: 117-130







ABSTRACT
This research was developed to track the microbial composition of raw milk obtained from twelve successive milking operations from the same farm with a special attention to lactococci, enterococci, lactobacilli and leuconostocs. Enterococci, lactobacilli and leuconostocs were detected at low levels. Lactococci represented the most abundant microflora. The dominance of Lc lactis subsp. cremoris from days 1 to 5 was followed by the dominance of Lc lactis subsp. lactis. Technological characterizations, Rep-PCR and whole-cell protein patterns analyses revealed the existence of two groups of lactococci. The first group designed as fundamental microflora included strongly acidifying strains. They were present day after day. The second group-transitory microflora-was observed occasionally and was less acidifying. A major influence of the biofilms of the milking equipment was postulated to explain the stability and the composition of the fundamental microflora. The implication of these observations on natural whey starters` composition and cheese particularity is discussed.
PDF Abstract XML References Citation
How to cite this article
Dalmasso Marion, Prestoz Sylvie, Rigobello Veronique and Demarigny Yann, 2008. Evolution of the Raw Cow Milk Microflora, Especially Lactococci,Enterococci, Leuconostocs and Lactobacilli over a Successive 12 Day Milking Regime. International Journal of Dairy Science, 3: 117-130.
DOI: 10.3923/ijds.2008.117.130
URL: https://scialert.net/abstract/?doi=ijds.2008.117.130
DOI: 10.3923/ijds.2008.117.130
URL: https://scialert.net/abstract/?doi=ijds.2008.117.130
INTRODUCTION
Cheese making implies two main successive processes. The milk is firstly clotted and the resulting curd acidified thereafter. Acidification results from the metabolism of Lactic Acid Bacteria (LAB), which transform lactose into lactic acid. This action is essential to allow the curd to reach the proper pH value and to remove the whey. LAB can originate from four different sources depending on the process: the raw milk, the cheese plant environment, commercial starters and the whey from the previous cheese making. This last practice called back slopping is still largely used in many countries and for a great variety of processes. The whey is recovered just after molding and incubated until the next cheese making process (Gatti et al., 2003). During this culture, LAB level increases dramatically as a consequence of the multiplicity of genus and species. The resulting whey starter-designed thereafter as Natural Whey Starter (NWS)-consists of complex microbial associations (Reinheimer et al., 1996). The LAB population originates from three main sources, (1) the previous cheese making, (2) the microbial environment of the cheese plant and (3) the raw milk microflora (Bertoni et al., 2001).
The role of LAB is therefore mainly limited to the first steps of cheesemaking. During the ripening process, starter LAB are less important than other flora such as, for instance, Non Starter Lactic Acid Bacteria (NSLAB). Nevertheless, they still contribute to the development of the typical cheese aroma and texture. LAB possess many enzymes-namely endo and exopeptidases-which can directly contribute to the appearance of sapid molecules, or ultimately generate aromatic compounds precursors (Mauriello et al., 2001; Herreros et al., 2003).
Natural Whey Starters (NWS) are particularly interesting for these two enzymatic properties. The diversity of strains which composed these bacterial communities is assumed to contribute to the enhancement of the typical flavor of many traditional cheeses and among them many products of Controlled Denomination of Origin (CDO).
If the microbial diversity of NWS is commonly described as positive, a major problem may arise in case of any technological problem. In particular, acidification defects are more and more reported by cheese makers using NWS. Such problems may originate from diverse causes and generate a great array of consequences among which, sudden stops or lengthening of the milk acidification curves frequently described. However, it is at the present time almost impossible to identify the origin of the problem and therefore to propose an appropriate solution because of the microbial complexity of NWS.
Among the four LAB sources which participate in the NWS diversity, raw milk is often described as a major, although fluctuating, source of LAB (Bachmann et al., 1996; Centeno et al., 1996; Manolopoulou et al., 2003; Duthoit et al., 2005). The influence of the raw milk microflora on the cheese characteristics has been demonstrated in the past (Herreros et al., 2007), even if most of the research was not specifically focussed on NWS. At the present time, no experiment has been published on the specific influence of raw milk LAB on the complexity of NWS ecosystems. Such work faces to two main difficulties. The raw milk LAB microflora is quantitatively rather low and its contribution difficult to assess. Moreover, the study of NWS implies a dynamic approach on several consecutive days and little is known about the evolution of LAB from day to day in raw milk of the same farm origin.
Consequently, the composition of raw cow milk samples coming from the same farm was followed over 12 consecutive days. The aim of this study was to evaluate LAB biodiversity, with a specific focus on lactococci, leuconostocs, lactobacilli and enterococci, from day to day. In a future work, we will study the influence of this LAB dynamic on the composition of natural whey starters.
MATERIALS AND METHODS
Milk Sampling and Colony Enumeration
Raw cow milk samples were collected from the same farm over twelve consecutive days in February 2006. Mesophilic aerobic microflora, leuconostocs, enterococci, yeasts and moulds, total and faecal coliforms, Pseudomonas were enumerated respectively on PCA, MSE, KF, GGC, VRBL and CFC agar as described by Desmasures et al. (1997). Lactococci were checked on Turner agar (Curk et al., 1994), facultative heterofermentative lactobacilli on FH-agar (Isolini et al., 1990); Micrococcaceæ on Mannitol Salt Agar (Demarigny, 1997) and corynebacteria on Cheese Ripening Bacterial Medium (Denis et al., 2001).
After enumeration, a maximum of four colonies were selected from Turner, MSE, KF and FH agar. After microscopic examination, cocci were purified on M17 agar (Biokar diagnostics, 60000 Beauvais, France) (30 °C, 24 h) and bacilli strains were purified on MRS agar (Biokar diagnostics) (anaerobiosis, 37 °C, 24 h). Each isolate was finally stored at -80 °C. One hundred and thirty six strains were collected and characterized from raw milks -40, 40, 40 and 16 isolates respectively from MSE, Turner, KF and FH agar.
Strains Characterisation
After thawing, lactococci, leuconostocs and enterococci were cultivated on M17 broth (Biokar diagnostics) for 24 h at 30 °C and lactobacilli on MRS broth (Biokar diagnostics) for 24 h at 30 °C. Cells were then harvested and washed twice in 0.09 g L-1 NaCl (Sigma) distilled water. Optical density at 600 nm was adjusted between 0.2 and 0.3. This final suspension was used to inoculate at 2% the characterization media. All the incubations were made at 30 °C.
Biochemical Characterization
Biochemical Characterization of Lactococci and Leuconostocs
Salt resistance, growth at 15 and 37 °C, sugars fermentation, arginine dihydrolase capability and citrate utilization were studied as described by Demarigny (1997).
Biochemical Characterization of Enterococci
Sugar fermentation was tested on API 20A basic medium (BioMérieux, 69280 Marcy l`Etoile, France). 0.5 mL of a saccharose, glycerol or L-arabinose solution (50 g L-1, Sigma) was added to 4.5 mL of medium. Growth at 45 °C and pH = 9.6 was performed on M17 broth. Citrate utilization was checked on KCA agar (Nickels and Leesment, 1964).
Biochemical Characterization of Lactobacilli
Catalase test, arginine dihydrolase capability, citrate utilization, growth at 15 and 45 °C, production of CO2 and sugar fermentation for the characterization of lactobacilli were performed as described by Demarigny (1997).
Technological Characterization
Isolates were all analysed for the following technological aptitudes: acidification abilities, proteolysis, autolysis and behaviour on lithmus milk as described by Demarigny et al. (2006).
Whole-Cell Protein Patterns
The procedure followed was described by Demarigny et al. (2006) to study the diversity of lactococci from natural whey starters.
Extraction of Total DNA
From a single culture broth incubated at 30 °C for 24 h, total DNA was extracted by using the Nucleospin tissue kit (Macherey-Nagel, 67722 Hoerdt, France).
PCR Amplification and Gel Electrophoresis
The strains were confirmed to belong to Enterococcus, Leuconostoc mesenteroides and Lactococcus lactis subsp. lactis and cremoris by means of PCR-based methods. Enterococcal DNA were amplified using primers E1 and E2 (Sigma), biding to positions 632-646 and 1353-1369, respectively and corresponding to positions in the E. coli 16S rRNA sequence, according to Deasy et al. (2000). 16S rDNA fragments of Ln. mesenteroides were amplified using primers Lnm1, corresponding to conserved E. coli 16S rRNA position 185 forward and Lnm2 (position 470 reverse) (Sigma) according to Cibik et al. (2000). Lactococcus lactis subsp. lactis or subsp. cremoris DNA amplification was performed using primers His1 and His2 (Sigma), biding to positions 671-688 and 1587-1604, respectively and corresponding to positions in Lc. lactis subsp. lactis NCDO 2118 numbering, according to Corroler et al. (1998). Inter-Repetitive Extragenic Palindromic sequences were amplified by means of two 18-mer primers in combination (Rep1R-Dt, Rep2-D) (Sigma) as described by Bouton et al. (2002) for REP-PCR. In all cases, amplification reactions were performed in a final volume of 25 μL containing 1x reaction PCR buffer (Sigma), 0.4 μM of each opposing primers, 1 mM MgCl2 (Sigma), 0.2 μM of each deoxynucleoside triphosphate (Sigma), 0.5U Tap DNA Polymerase (Sigma) and 5 μL of DNA. The primer sequences and the PCR amplification conditions applied are recapitulated on Table 1. Amplification cycles were performed with a Thermal Cycler (Biorad, 92430 Marne-la-Coquette, France). PCR products of 25 μL were electrophoresed in a 10 g L-1 Seakem GTG agarose gel (Sigma) in TBE (Tris - Borate - EDTA pH 8) at 100 V for 3 h. The 123-pb DNA ladder (Invitrogen, 95613 Cergy Pontoise, France) was used as a size standard. The DNA fragments were stained with ethidium bromide (Sigma), viewed under UV light (302 nm, Biorad) and photographed on a digital camera (Camedia C-5060, Olympus).
| Table 1: | Primer sequences, amplification and application of PCR reactions |
 | |
Computer Analyses of REP-PCR Profiles
The band patterns were normalized and processed according to the same procedure as whole-cell protein patterns. The similarity coefficient (80%) of the Rep-PCR technique was evaluated by studying two strains six fold.
Statistical Analyses
Correlations, Multiple Correspondence Factorial Analyses (MCFA) between isolates and technological abilities and Hierarchical Classifications (HC) were performed using the STATITCF software (5th version, 1995, Institut Techniques des Céréales et des Fourrages, Paris, France).
RESULTS
Enumeration Results
The mesophilic aerobic microflora level ranged between 3.2 ± 0.08 and 5.2 ± 0.08 log cfu mL-1. Corynebacteria and Micrococcaceæ were present systematically at constant levels, respectively 3.4 ± 0.12 and 2.2 ± 0.16 log cfu mL-1 (Fig. 1). Yeast counts were inferior to 1.0 ± 0.2 log cfu mL-1, except on days 2, 8, 11 and 12 (2-3 ± 0.2 log cfu mL-1). Moulds were detected irregularly in milk and at levels that barely exceeded 2.0 ± 0.66 log cfu mL-1, except on days 11 and 12 (over 3.0 ± 0.66 log cfu mL-1). Coliforms and Pseudomonas were temporarily found at levels ranging from 1.2 to 4.4 ± 0.4 log cfu mL-1.
The evolution of the four main LAB microflora was based on the use of selective-FH agar, KF-and elective-Turner, MSE-media. The selectivity of these media being erratic, it is admitted to refer to a specific population by adding the term presumed. In some cases however, the gap between presumed and real levels is too large to be relevant. As a consequence, even the additional use of presumed does not refer to any correct tendency. In this study, this difficulty was overcome by characterizing isolates from the four LAB media. 136 isolates were picked following the number of colonies enumerated each day: 16 from FH agar, 40 from each other media. These isolates were identified by means of phenotypic and PCR-based methods. The isolates from Turner and FH media were all classified as respectively Lactococcus and Lactobacillus. Thirteen isolates out 40 from MSE were identified as Leuconostoc, while the 27 remaining isolates were further characterized as lactococci.
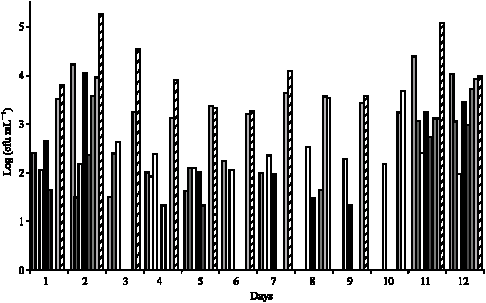 | |
| Fig. 1: | Evolution of the non-LAB microflora- ( |
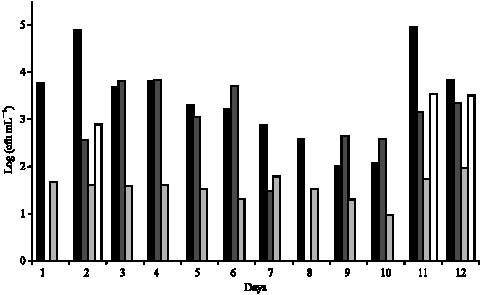 | |
| Fig. 2: | Evolution of the LAB population- ( |
Thirty-one isolates out of 40 from KF were referred to Enterococcus genus. It was not possible to clearly link the nine other isolates with an identified population. These results allowed to calculate the correct levels corresponding to each population. For instance on day 1, if two isolates from KF out of four were clearly identified as enterococci, the number deduced from plate counting was weighted by a coefficient equal to 2/4 (Fig. 2).
Lactococci level was relatively stable from day 1 to day 4 (between 3.0 and 5.0 ± 0.1 log cfu mL-1) and decreased thereafter to reach 2.0 ± 0.1 log cfu mL-1 on days 9 and 10. Then, the number of lactococci increased again. Enterococci were detected every day but never overstepped 2.0 ± 0.4 log cfu mL-1.
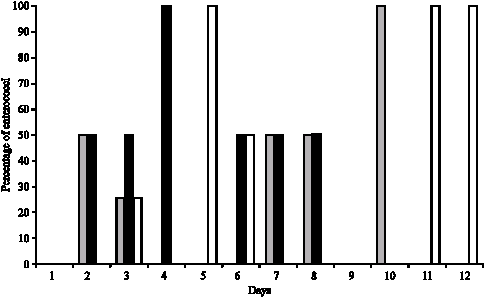 | |
| Fig. 3: | Main enterococcal groups- ( |
Lactobacilli were only detected on days 2, 11 and 12. Leuconostocs were detected everyday except on days 1 and 8. The levels of these two populations ranged between 2.4 and 4.0 ± 0.3 log cfu mL-1. A strong correlation was observed between leuconostocs and lactobacilli (r = 0.95).
It is noteworthy that lactococci, mesophilic aerobic microflora and total coliforms were strongly correlated (r>0.82), inferring a potential common origin for these three microflora.
Isolates Characterizations
Leuconostocs and Lactobacilli
The thirteen strains belonging to the Leuconostoc genus appeared to have the phenotypic characteristics of Ln mesenteroides subsp. mesenteroides. Leuconostocs mean acidifying and proteolytic activities were rather weak, respectively 0.20 ± 0.02 pH units and 9.2 ± 4 μmoles of glycine mL-1.
Lactobacilli were characterized by phenotypic means. They all belonged to the facultative heterofermentative lactobacilli group. Further identifications indicated that the isolates were classified as Lb plantarum or Lb paracasei subsp. paracasei.
Enterococci
Fourteen isolates were identified as Enterococcus fæcalis, ten as Ec durans and two as Ec fæcium. Five enterococci could not be accurately identified. Distribution of these three populations during the 12 days did not reveal any specific trend. Ec fæcalis strains were more commonly identified at the beginning of the period and Ec durans more common at the end. Despite this dual distribution, no correlation was observed between the three species.
Enterococci were divided into three groups-I, II, III-thanks to HC analysis (Fig. 3). The mean acidifying and proteolytic activities were equal to 0.66 ± 0.06 pH units and 3.6 ± 0.4 μmoles of glycine mL-1 for group I strains, 0.56 ± 0.08 pH units and 2.4 ± 0.7 μmoles of glycine mL-1 for group II strains and 0.26 ± 0.13 pH units and 1.7 ± 0.9 μmoles of glycine mL-1 for group III strains. A negative correlation was observed between III and II or III and I populations r = -0.68. Group III strains were generally present when the strains of the other groups were absent or at low rates.
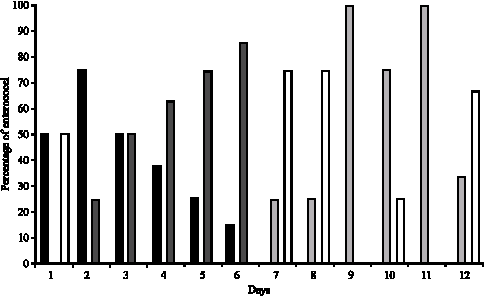 | |
| Fig. 4: | Main lactococcal groups- ( |
Lactococci
Forty lactococci were picked from Tuner agar and 27 from MSE agar. All these isolates displayed the phenotypic features of the subspecies Lactococcus lactis subsp. lactis. For example, they all showed positive arginine dihydrolase capability. They were, also, able to grow in a 40 g L-1 NaCl culture broth and to use maltose and ribose. Growth in litmus milk indicated that over 95% of the isolates were able to reduce, acidify and clot the milk in less than 24 h. None of the strains were able to use citrate to produce diacetyl.
The confirmation of the species identification was based on the identification of the histidine biosynthesis operon region, because of its Lc lactis species specificity. Based on its size, the resulting fragment was subspecies specific. Lc lactis subsp. cremoris strains have a DNA sequence of about 200 bp between the orf3 and hisC genes, which is not present in Lc lactis subsp. lactis strains. This work showed that all the strains belonged to Lc lactis species. However, 50% of the strains were classified in the cremoris subspecies. From day 1 to day 4, only Lc lactis subsp. cremoris was present in the milk. On day 5, a change was observed, the two subspecies being equally represented. Thereafter, Lc lactis subsp. cremoris disappeared to be replaced by Lc lactis subsp. lactis.
MCFA and HC analysis allowed to separate lactococci into four main groups, A, B, C and erratic strains (Fig. 4). If autolysis was roughly identical irrespective of the strains, some slight differences were observed on the basis of proteolysis and acidification aptitudes. The mean acidifying and proteolytic activities were equal to 1.70 ± 0.01 pH units and 6.7 ± 0.6 μmoles of glycine mL-1 for group A strains, 1.58 ± 0.23 pH units and 5.1 ± 1.6 μmoles of glycine mL-1 for group B strains and 1.74 ± 0.14 and 6.4 ± 1.1 μmoles of glycine mL-1 for group C strains. Strains which could not fall into one of these three groups were designed as erratic strains.
From days 1 to 6, the proportion of A strains decreased gradually in time. A strains represented 75% of the lactococci isolates on day 2 and only 15% on day 6. At the same time, group C became more and more important from day 2-25% of the isolates-to day 6-85% of the isolates. C strains suddenly disappeared thereafter, to be replaced by B strains. Erratic strains represented in some cases the dominant flora among lactococci isolates. Their presence was rather congruent with that of B strains.
 | |
| Fig. 5: | Dendrogramme of 58 lactococcal whole-cell protein patterns by SDS-PAGE. Each pattern is identified by a milk number indicating the origin of the strain, e.g., all the lactococcal strains from milk 1 are labelled Milk 1 on the dendrogramme. Clusters are materialized by bold horizontal lines and numbered from 1 to 11. The similarity coefficient of 90% is materialized by a bold vertical line |
The whole-cell protein patterns of the 67 lactococci were analyzed by SDS-PAGE. Nine strains providing unusable profiles were not kept. The 58 remaining patterns were divided into 11 strain clusters, on the basis of a similarity coefficient of 90% (Fig. 5). Clusters 6 and 8 grouped 21 and 19 profiles i.e., 69% of the lactococci strains. Among these two clusters, strains originated from milks 1 to 11, meaning that along this period, a fundamental microflora had settled durably. The 18 other patterns were shared among the nine remaining clusters. They corresponded with strains isolated from milks 3, 4, 6 and 8 to 12. Compared with the leading microflora represented by clusters 6 and 8, this secondary microflora could be designed as transitory.
It is interesting to note that these results partially confirmed the observations deduced from acidifying abilities and proteolysis analysis. C strains profiles were distributed among several clusters. A and B strains profiles were essentially concentrated in clusters 6 and 8. These strains could be linked with the fundamental microflora and C and erratic strains to the transitory microflora. The distribution of A, B and C strains among the clusters allowed to calculate the diversity coefficient H = Σ Pi log Pi (with Pi = number of strains in cluster i/total number of strains) (Shannon and Weaver, 1983). A, B and C populations diversity coefficients were respectively equal to 30, 54 and 71%. A population was the less diversified, strains being shared among clusters 6 and 8. C population included six different profiles.
 | |
| Fig. 6: | Example of a dendrogramme of lactococcal profiles by REP-PCR. Each pattern is identified by a milk number indicating the origin of the strain, e.g., all the lactococcal strains from milk 1 are labelled Milk 1 on the dendrogramme. Clusters are materialized by bold horizontal lines and numbered from 1 to 8. The similarity coefficient of 80% is materialized by a bold vertical line |
A second analysis was made by Rep-PCR. An example of the results obtained with this technique is displayed on Fig. 6. Eight clusters were obtained. 83% of the lactococci classified in the SDS-PAGE cluster 6 were again in the same cluster (3). This observation confirms the existence of some form of stable leading microflora in the milk from day to day. The other strains, in particular those from SDS-PAGE cluster 8, were not grouped into a specific cluster and were scattered among many clusters.
DISCUSSION
The origin of the raw milk microflora is still currently under study. The potential sources of contamination include the skin of the cow, feed dusts, milking equipment and finally the environment of the cheese plant. Among the different microflora which can be identified in raw milk, lactic acid bacteria seem particularly attractive for the cheese maker. These bacteria, namely lactococci and lactobacilli, are acknowledged to contribute to curd acidification. They are also suspected to bring interesting features-directly during the cheese making process or thereafter during the ripening stage- to the cheese. Such elements enable to give the cheese its original flavour typical of its geographical origin. These bacteria are mainly characterized by their strong rusticity. This quality leads them to colonize the curd at the end of the cheese making process, in spite of hard selective conditions: their level is at least 10000 times lower than the starter, some nutrients are rapidly falling in numbers-non protein nitrogen for instance-, acidification is very intense and in some cases, the technological temperature is far from their optimum.
This work was undertaken to monitor the evolution of the main LAB microflora during twelve consecutive days of milking, with a specific focus on the Lactococcus group. It allowed the study of the LAB microflora dynamic from day to day, which had never been done before at such an extensive scale. Three different methodologies were combined to study enterococci, leuconostoc, lactobacilli and lactococci: technological aptitudes, PCR methods and whole-cell protein patterns. Corsetti et al. (2003) followed a similar approach-wall-cell protein patterns, RAPD-to study sourdough lactic acid bacteria. They concluded to the effectiveness of the combination of two different methods to classify invasive microbial populations. In a recent study, we also underlined the necessity to cross two different microbial techniques to observe NWS, each method giving complementary results (Demarigny et al., 2006).
On a twelve-day period, the mean mesophilic aerobic flora was centered around 4.0 ± 0.08 log cfu mL-1, with some variations. These values are similar to the observations already made by many authors on raw milk samples. The evidence of a correlation between lactococci, coliforms and mesophilic aerobic flora emphasizes a potential common origin. Lactococci levels, in particular, are frequently close to mesophilic aerobic flora levels. Lactococci probably originate from biofilms present in the milking equipment. This assertion is based on the results obtained by Laithier et al. (2004), which showed that biofilms were mainly composed of LAB, the other flora being generally-yet not systematically-far less important. In a recent study, Kagkli et al. (2007) established that milk enterococci and lactobacilli also originated from milking equipment. In our case, however, no correlation was observed between the levels of these two populations.
Although frequently counted, enterocci did not reach high levels in milk. Isolates were identified as Ec faecium, Ec faecalis and Ec durans, three species commonly found in raw milks (Aquilanti et al., 2006). Their technological interest appeared limited because of low milk acidifying and proteolytic abilities, which is in accordance with Morandi et al. (2006). Some trends were observed following the acidifying proteolytic characteristics of the strains isolated. Lactobacilli were inconstant as observed by Bouton et al. (2005). These authors indicated that variations in the levels of fermentative lactobacilli could be partially explained by the type of feeding used by the dairy farmer, farm practices and general hygiene of the premises.
The specific enumeration of leuconostocs was particularly difficult to achieve. The MSE medium only allowed counting dextrane-producing strains of leuconostocs. This lack of selectivity was made worse by the pressure exerted by the other microorganisms able to grow on MSE (Corry et al., 2007). In spite of this problem, we only found small numbers of Leuconostoc mesenteroides subsp. mesenteroides. This observation reinforces the assertion that bacteria from the Leuconostoc genus are a minor microflora in milk (Zamfir et al., 2006).
Lactococci appeared to be the principal LAB population, their proportion ranging between 93 and 100% of the total LAB microflora. They, also, constituted the major part of the milk microflora. Their levels varied considerably, from 2.0 to 5.0 ± 0.1 log cfu mL-1. These results are congruent with former data. For instance, Corroler et al. (1998) indicated that lactococci varied between 2 and 3 log cfu mL-1, following the origin of the sample and the season.
All lactococci were confirmed to be part of the Lactococcus lactis species. Most of strains exhibited phenotypic characteristics of the subspecies lactis whereas they have been found to own the genetic profile of the subspecies cremoris. This observation is in complete agreement with the study completed by Ward et al. (1998) in which more than 70% of Lc lactis were found to have the phenotype of the subspecies lactis and the genotype of the subspecies cremoris. This ambiguous taxonomic assignment of Lc lactis strains can be directly compared to the works of Salama et al. (1995). Actually, these authors reported that phenotypic changes from lactis to cremoris subspecies had been observed before. It is particularly true for the phenotypic change from arginine negative to arginine positive capability, which is attributed to transducing phage. Consequently, several methods have to be used and new phenotypic characteristics have to be investigated in order to discriminate properly the two subspecies lactis and cremoris. For example, Nomura et al. (1999) proposed to test the presence of γ-aminobutyric acid, a by-product only produced by the subspecies lactis. We can wonder, however, if the phenotypic characterisation is still a relevant method to identify lactococci.
During the 12 day period, we observed two different behaviors. From day 1 to day 6, Lc lactis subsp. cremoris was the only subspecies identified, replaced later by Lc lactis subsp. lactis. If we suppose a major influence of biofilms on the LAB population, this shift appears surprising. Indeed, dairy farm practices were identical from day to day during the twelve consecutive days. Otherwise, we can argue with Salama et al. (1995) that Lc lactis subsp. cremoris acquired its lactis characteristics inside the biofilm by transducing phages. This hypothesis would be then fundamental. It would then imply that the release of Lactococcus strains from biofilms would be accompanied by the concurrent release of phages. If raw milk lactococci are intended to settle and even to dominate in NWS, the origin of acidification defects during cheese making would be possibly explained by these phages from biofilms.
Moreover, in a preceding study (Demarigny et al., 2004) NWS high levels of Lc lactis subsp. cremoris were observed in cheese. This population displayed the phenotypic features of the cremoris subspecies. Considering that Lc lactis subsp. cremoris are frequently added in commercial starter (Coppola et al., 2006), it would indicate that NWS dominant populations partly originate from the added starter.
The analysis of Lactococcus strains patterns indicated the occurrence of a fundamental microflora, exhibiting stable and interesting technological behaviors and a transitory microflora. This observation is always congruent with the biofilm hypothesis and the observations made by Laithier et al. (2005). The authors reported that lactococci and more generally LAB from biofilms could be classified among ten different classes following their abilities.
Our results were confirmed by using three methodological means at the same time. Unlike Casalta et al. (2005), the Rep-PCR characterization partially failed to disclose different patterns. This technique may therefore be inappropriate to study the lactococci diversity from the same ecosystem. Apart from the fundamental population, the presence of SDS-PAGE patterns corresponding to particular technological features-less important acidification ability-could indicate another source of lactococci, derived from the milk bulk and not from biofilm. The diversity coefficients calculation seemed to confirm that the transitory microflora was more diverse than the fundamental microflora. This could confirm a different origin of the transitory isolates.
CONCLUSION
The dynamics and the characteristics of the main LAB populations in raw cow milk were studied on a twelve-day period of milking. It is allowed to suppose that lactococci which dominated over the other LAB exerted an inhibition effect. It will be interesting to test in another study the ability of wild raw milk lactococci to produce antimicrobial compounds, active against the other LAB microflora.
According to preceding research, we considered biofilms, which settle in the milking equipment, as the main origin of lactococci. We postulated the influence of transducing phages, as indicated by Salama et al. (1995) to explain the stability of the lactococci characteristics. It would imply that the study of biofilms phages would be of a great interest to sort out cheese making defects. We can still wonder if the diversity of raw milk microflora contributing to the cheese particularity is not initially due to the exchanges of transducing phages.
Finally, it could be interesting to monitor the LAB microflora on many farms for many weeks. The LAB dynamics and specific farm practices have to be taken into account too.
ACKNOWLEDGMENTS
The authors would like to thank Marc Chareyron for his help in the writing of this paper and Fanny Moignard for her technical support.
REFERENCES
- Aquilanti, L., L. DellAquila, E. Zannini, A. Zocchetti and F. Clementi, 2006. Resident lactic acid bacteria in raw milk Canestrato Pugliese cheese. Lett. Applied Microbiol., 43: 161-167.
Direct Link - Bachmann, H.P., D.A. McNulty, P.L.H. McSweeney and M. Rüegg, 1996. Experimental designs for studying the influence of the raw milk flora on cheese characteristics: A review. J. Soc. Dairy Tech., 49: 53-56.
Direct Link - Bertoni, G., L. Calamari and M.G. Maianti, 2001. Producing specific milks for speciality cheeses. Proc. Nutr. Soc., 60: 231-246.
PubMed - Bouton, Y., P. Guyot, E. Beuvier, P. Tailliez and R. Grappin, 2002. Use of PCR-based methods and PFGE for typing and monitoring homofermentative lactobacilli during Comte cheese ripening. Int. J. Food Microbiol., 76: 27-28.
Direct Link - Casalta, E., J.M. Cachenaut, C. Aubert, F. Dufrene, Y. Noel and E. Beuvier, 2005. Application of specific starters for the manufacture of Venaco cheese. Lait, 85: 205-222.
Direct Link - Centeno, J.A., S. Menendez and J.L. Rodriguez-Otero, 1996. Main microflora present as natural starters in Cebreiro raw cow's-milk cheese (Northwest Spain). Int. J. Food Microbiol., 33: 307-313.
Direct Link - Cibik, R., E. Lepage and P. Tailliez, 2002. Molecular diversity of Leuconostoc mesenteroides and Leuconostoc citreum isolated from traditional french cheeses as revealed by RAPD fingerprinting, 16S rDNA sequencing and 16S rDNA fragment amplification. Syst. Applied Microbiol., 23: 267-278.
CrossRefPubMedDirect Link - Coppola, S., V. Fusco, R. Andolfi, M. Aponte, G. Blaiotta, D. Ercolini and G. Moschetti, 2006. Evaluation of microbial diversity during the manufacture of Fior di Latte di Agerola, a traditional raw milk pasta-filata cheese of the Naples area. J. Dairy Res., 73: 264-272.
Direct Link - Corroler, D., I. Mangin, N. Desmasures and M. Gueguen, 1998. An ecological study of lactococci isolated from raw milk in the camembert cheese registered designation of origin area. Applied Environ. Microbiol., 64: 4729-4735.
Direct Link - Corry, J.E.L., B. Jarvis, S. Passmore and A. Hedges, 2007. A critical review of measurement uncertainty in the enumeration of food micro-organisms. Food Microbiol., 24: 230-253.
Direct Link - Corsetti, A., M. De Angelis, F. Dellaglio, A. Paparella, P.F. Fox, L. Settanni and M. Gobbetti, 2003. Characterization of sourdough lactic acid bacteria based on genotypic and cell-wall protein analyses. J. Applied Microbiol., 94: 641-654.
Direct Link - Curk, M.C., J.M. Boeufgras, B. Decaris, F. Gavini, K. Kersters, J.P. Larpent, P. Le Bourgeois, P. Renault, H. de Roissart and C. Rouvier, 1994. Methods Used for the Identification of Lactic Acid Bacteria. Dans: Bactéries lactiques. In: Lactic Acid Bacteria, De Roissart, H., F.M. Luguet, H. Roissart and F.M. Luguet (Eds). Vol. 1. Uriage, Lorica, pp: 141-168.
- Deasy, B.M., M.C. Rea, G.F. Fitzgerald, T.M. Cogan and T.P. Beresford, 2000. A rapid PCR based method to distinguish between Lactococcus and Enterococcus. Syst. Applied Microbiol., 23: 510-522.
Direct Link - Demarigny, Y., 1997. Influence of raw milk microflora and technological parameters (salt contents and ripening temperature) on the evolution of the biochemical, microbiological and sensorial characteristics of semi-hard cheese. Ecole Nationale Supérieure de Biologie Appliquée à la Nutrition et à l'Alimentation. Poligny, France.
- Demarigny, Y., C. Sabatier, N. Laurent, S. Prestoz, V. Rigobello and M.J. Blachier, 2006. Microbiological diversity in natural whey starters used to make traditional Rocamadour goat cheese and possible relationships with its bitterness. Italian J. Food Sci., 18: 261-276.
Direct Link - Denis, C., M. Gueguen, E. Henry and D. Levert, 2001. New media for the enumeration of cheese surface bacteria. Lait, 81: 365-379.
Direct Link - Desmasures, N., F. Bazin and M. Gueguin, 1997. Microbiological composition of raw milk from selected farms in the Camembert region of Normandy. J. Applied Microbiol., 83: 53-58.
Direct Link - Duthoit, F., L. Tessier and M.C. Montel, 2005. Diversity, dynamics and activity of bacterial populations in registered designation of origin salers cheese by single-strand conformation polymorphism analysis of 16S rRNA genes. J. Applied Microbiol., 98: 1198-1208.
Direct Link - Gatti, M., C. Lazzi, L. Rossetti, G. Mucchetti and E. Neviani, 2003. Biodiversity in Lactobacillus helveticus strains present in natural whey starter c J. Applied Microbiol., 95: 463-470.
Direct Link - Herreros, M.A., J.M. Fresno, M.J. Gonzalez-Prieto and M.E. Tornadijo, 2003. Technological characterization of lactic acid bacteria isolated from Armada cheese (a Spanish goat's milk cheese). Int. Dairy J., 13: 469-479.
Direct Link - Herreros, M.A., R. Arenas, M.H. Sandoval, J.M. Castro, J.M. Fresno and M.E. Tornadijo, 2007. Effect of addition of native cultures on characteristics of Armada cheese manufactured with pasteurised milk: A preliminary study. Int. Dairy J., 17: 328-335.
Direct Link - Kagkli, D.M., M. Vancanneyt, C. Hill, P. Vandamme and T.M. Cogan, 2007. Enterococcus and Lactobacillus contamination of raw milk in a farm dairy environment. Int. J. Food Microbiol., 114: 243-251.
Direct Link - Laithier, C., Y.M. Chatelin, R. Talon, J. Barral, H. Tormo and Y. Lefrileux, 2005. Effectiveness in laboratory then in farms of cleaning/desinfection processes on positive selection of biofilms. Proceedings of the 12th Rencontres Recherche Ruminants Conference, December 7-8, 2005, Paris, France, pp: 367-370.
- Manolopoulou, E., P. Sarantinopoulos, E. Zoidou, A. Aktypis, E. Moschopoulou, I.G. Kandarakis and E.M. Anifantakis, 2003. Evolution of microbial population during traditional feta cheese manufacture and ripening. Int. J. Food Microbiol., 82: 153-161.
Direct Link - Mauriello, G., L. Moio, G. Moschetti, P. Piombino, F. Addeo and S. Coppola, 2001. Characterization of lactic acid bacteria strains on the basis of neutral volatile compounds produced in whey. J. Applied Microbiol., 90: 928-942.
Direct Link - Morandi, S., M. Brasca, C. Andrighetto, A. Lombardi and R. Lodi, 2006. Technological and molecular characterization of enterococci isolated from North-West Italian dairy products. Int. Dairy J., 16: 867-875.
Direct Link - Nomura, M., H. Kimoto, Y. Someya and I. Suzuki, 1999. Novel characteristic for distinguishing Lactococcus lactis subsp lactis from subsp cremoris. Int. J. Syst. Bacteriol., 49: 163-166.
Direct Link - Reinheimer, J.A., A. Quiberoni, P. Tailliez, A.G. Binetti and V.B. Suarez, 1996. The lactic acid microflora of natural whey starters used in Argentina for hard cheese production. Int. Dairy J., 6: 869-879.
CrossRefDirect Link - Salama, M.S., T. Musafija-Jeknic, W.E. Sandine and S.J. Giovannoni, 1995. An ecological study of lactic acid bacteria: Isolation of new strains of Lactococcus including Lactococcus lactis subspecies cremoris. J. Dairy Sci., 78: 1004-1017.
Direct Link - Shannon, G.E. and W.W. Weaver, 1963. The Mathematical Theory of Communities. 1st Edn., University of Illinois Press, Urbana, ISBN: 978-0252725463, pp: 111-125.
Direct Link - Ward, L.J.H., J.C.S. Brown and G.P. Davey, 1998. Two methods for the genetic differentiation of Lactococcus lactis ssp. lactis and cremoris based on differences in the 16S rRNA gene sequence. FEMS. Microbiol. Lett., 166: 15-20.
Direct Link