
G.R. Ghorbani
Department of Animal Science, Isfahan University of Technology, Isfahan, Iran
A. Jafari
Department of Animal Science, Isfahan University of Technology, Isfahan, Iran
A.H. Samie
Department of Animal Science, Isfahan University of Technology, Isfahan, Iran
A. Nikkhah
Department of Animal Science, University of Manitoba, Canada
International Journal of Dairy Science
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 79-84







ABSTRACT
Scarcity of data exists on potential use of exogenous enzymes for young calves. The objective of this study was to assess the effects of applying Exogenous Non-starch Polysaccharidases (ENP) to pre-weaning starter concentrate on starter intake, nutrient digestibility and calf growth. Eighteen neonatal Holstein calves (47.9±2.5 kg body weight) were monitored for 84 day. Treatments included pre-weaning calf starters with 1) no enzyme additives (C), 2) enzyme additive A (EA, 0.6 mL kg-1 starter), or 3) enzyme additive B (EB, 1.9 mL kg-1 DM of starter). The activity of endo-cellulase, exo-cellulase and xylanase was measured using carboxymethyl cellulose, filter paper test and oat spelt, as the respective substrates. The resulting activity (μ mol/mL/min) of exo-cellulase, endo-cellulase and xylanase were respectively 1437, 788 and 7476 for EA and 1446, 1350 and 5091 for EB. No enzymes were added to post-weaning diet. Weaning Criterion (WC) was defined as the calf age at a daily intake of 680 g starter and estimated with regression equations. Mixed models of repeated measures with best fitted covariance structures were used to analyze the data. The calves fed EA-supplemented starter had higher NDF apparent total tract digestibility at week-4, but not at week-8, compared to control and EB-fed calves. However, NDF digestibility was lower at week-12 in ENP-fed calves than in control calves. The ENP did not affect the average daily starter intake, growth rate and WC. Overall, applying ENP to pre-weaning calf starter did not improve calf performance in this study. Further studies preferably using a mixture of cell-content and cell-wall polysaccharidases in both pre- and post-weaning starters would be warranted.
PDF Abstract XML References
How to cite this article
G.R. Ghorbani, A. Jafari, A.H. Samie and A. Nikkhah, 2007. Effects of Applying Exogenous, Non-Starch Polysaccharidases to Pre Weaning Starter Concentrate on Performance of Holstein Calves. International Journal of Dairy Science, 2: 79-84.
DOI: 10.3923/ijds.2007.79.84
URL: https://scialert.net/abstract/?doi=ijds.2007.79.84
DOI: 10.3923/ijds.2007.79.84
URL: https://scialert.net/abstract/?doi=ijds.2007.79.84
INTRODUCTION
Despite their complex mechanisms of action and variable animal response, Exogenous Non-starch Polysaccharidases (ENP) have received much research interest in ruminant nutrition (Beauchemin et al., 2004). Improvements in milk yield in lactating cows (Beauchemin et al., 1998) and in Average Daily Gain (ADG) in beef steers (Beauchemin et al., 1995) after dietary application of ENP have been reported. Apparently, no or little inclusion of forage-fiber in pre-weaning calf diets has attracted much less interest to the use of ENP for young calves. From rumen development and health perspectives, however, little dietary forage would not explain overlooking the desperate need for greater utilization of non-forage polysaccharides by young dairy calves (Baldwin et al., 2004).
Pre-ruminant calves possess negligible activity of the enzymes degrading starch and cell-wall polysaccharides (Van Soest, 1994). Meanwhile, adequate supply of Volatile Fatty Acids (VFA) from microbial fermentation is crucial for the proliferating expansion of rumen epithelia and effective hepatic metabolism of VFA (Baldwin et al., 2004). Early establishment of fibrolytic capacity appears determining for such early development in the rumen and hepatic metabolism to occur (Van Soest, 1994). Universally, dairy calves are usually weaned between 5 to 12 weeks of age, depending on growth rate, body size and milk price. The milk price, nonetheless, may not play a major role at the expense of a desirable calf growth. Accordingly, early utilization of starter polysaccharides by the calf could hasten the reticulo-rumen maturity, lower the weaning age, save milk and reduce labor costs (NRC, 2001). Dairy industries with rather long commercial weaning age (>5-7 weeks) and a growing consumer demand for dairy products would benefit the most from such optimized nutrient utilization by dairy calves.
Documented research on the impact of applying ENP to calf starter diet on young calf performance is scarce. It is thus unknown if applying exogenous enzymes can enhance nutrient utilization and in turn ease lowering the weaning age. The objective of this study was hence to determine the effects of adding ENP to pre-weaning starter concentrate on starter intake, nutrient digestibility, growth and weaning criterion in Holstein calves.
MATERIALS AND METHODS
Experimental Design and Calves
Eighteen neonate Holstein calves (9 males and 9 females) with body weight (BW) of 47.9±2.5 (mean±SD) were used in a randomized complete block design with repeated measures. Calves were monitored for 84 day at Calf Raising Facilities of Foka Enterprise, the largest dairy herd in central Iran with approximately 2000 milking cows. Upon separation from dam shortly after calving, calves within each sex were assigned randomly to one of 3 treatments. Calves were then transferred into individual stalls bedded with clean wheat straw. Six calves including 3 males and 3 females were allocated to each treatment. One male calf on enzyme B was noticed unhealthy 3 week after beginning of the study and thus excluded from the trial. As all calves had entered the trial shortly after birth, the excluded calf could not be replaced. Calves were offered milk (10% of BW) twice daily at 0800 and 1500 h for 49 day. The daily offer of milk was halved from 49 to 55 day and ended at 56 day. Calves were offered ad lib a starter diet (Table 1) and fresh water for the entire experiment.
| Table 1: | Feed ingredients and chemical composition of the calf starter concentrate (DM basis) |
 | |
| 1Contained 250000 IU vit. A, 50000 IU vit. D, 1500 IU vit E, 2.25 g Mn, 7.7 g Zn, 20 g P, 20.5 g Mg, 186 g Na, 1.25 F, 3 g S, 14 mg Co, 1.25 g Cu, 56 mg I and 10 mg Se per kg supplement.2Calculated from NRC (2001).3Nonfiber carbohydrate = 100-(%NDF + %CP + %EE + %Ash) | |
Treatments and Enzymatic Activity
The experimental treatments included the starters supplemented with 1) no enzyme additives (C), 2) enzyme additive A or EA (Pro-Mote®, Biovans Technologies Inc., Omaha, NE; 0.6 mL kg-1 DM of starter) and 3) enzyme additive B or EB (Biozyme MT-4000 Finnfeeds International Ltd., Malborough, UK; 1.9 mL kg-1 DM of starter). The respective activity (μmol/mL/min) of exo-cellulase, endo-cellulase and xylanase were 1437, 788 and 7476 for EA; and 1446, 1350 and 5091 for EB. Each unit of enzyme activity was measured as μmol of reducing sugars released per min after adding 1 mL of the enzyme to a given substrate, Both EA and EB were analyzed for the enzymatic activity according to Mandel and Weber (1969). The activity of endo-cellulase, exo-cellulase and xylanase was measured using carboxymethyl cellulose (CMC), filter paper test and oat spelt as the respective substrates. Filter paper test and CMC were hydrolyzed by the cellulases for 1 h at 50°C under pH 4.8. The oat spelt was incubated with xylanase solution for 30 min at 50°C under pH 4.8. Both solutions were then added with 2 mL of 3,5-dinitrosalicylic acid and boiled for 15 min. The concentrations of reducing sugars liberated were read at 575 for glucose and 540 nm for xylose. Liquid enzyme supplements were diluted in distilled water (1:10 ratio of enzyme:water) before spraying onto the calf starter at 1 h before feeding. Control starter was similarly treated with water but without enzyme additives. The starter diet was then supplemented with ENP until 56 day of age and no enzyme additives were added to the post-weaning diet.
Calf Performance Measurements
Performance criteria were measured repeatedly for each calf in 4-week intervals at 28, 56 and 84 day of age. Calves were weighed at 0800 h for 2 consecutive day after feed was nocturnally withheld for 12 h at the end of each 4-week interval. Daily amount of starter offered and orts were recorded for each calf and their DM content was determined by oven-drying at 60°C for 48 h (AOAC, 1990). Feces were sampled by manual stimulation of anus at 1 h postfeeding for 2 consecutive days at each sampling. All feed and fecal samples were stored at -20°C until later analyses. The frozen fecal samples were thawed at room temperature and analyzed for DM (48 h at 60°C), ash (method No. 942.05), OM and ADF (method No. 973.18) according to AOAC (1990). All feed and fecal samples were analyzed for NDF using heat-resistant α-amylase (Van Soest et al., 1991). Acid Insoluble ash (AIA) was measured in feed and fecal samples and used as an internal marker to measure the apparent total tract nutrient digestibility (Van Keulen and Young, 1977).
Statistical Approaches
Data were analyzed as a linear mixed model for repeated measures (Wang and Goonewardene, 2004). The least square means estimation method was Restricted Maximum Likelihood and the method of calculating the denominator degrees of freedom was Kenward-Roger (SAS Institute, 1999). Fixed effects included treatment, sex, week and the interactions. The effect of calf nested within treatment by sex was considered random and used as the error term to test the effects of treatment, sex and treatmentxsex. Akaike's Information and Schwarz's Bayesian Criteria (Wang and Goonewardene, 2004) were used as the fit statistics to adopt the best fitted covariance structure in the final models. The growth rate for individual calves was additionally estimated by regressing the periodical BW values against time/calf age using linear regression equations. Polynomial equations were developed to regress the starter intake against time to estimate the weaning criterion or calf age at a daily intake of 680 g starter (NRC, 2001). Estimates for growth rates and weaning criterion were subsequently subjected to variance analysis with fixed effects of treatment, sex and treatment xsex. Least square means were compared using Tukey’s multiple range test (SAS, 1999) and significance levels were declared at p<0.05.
RESULTS
Daily starter intake of calves across three 4-week periods were not affected by applying the ENP to pre-weaning starter diet (p = 0.99, Table 2). Comparably, body weight (p = 0.73) and ADG (p = 0.51) of calves remained unchanged among treatments (Table 2).
| Table 2: | Effect of exogenous, non-starch polysaccharidases in pre-weaning calf starter on starter intake, feed efficiency, body weight, Average Daily Gain (ADG) and nutrient digestibility across pre- and post-weaning phases (84 days) in Holstein calves |
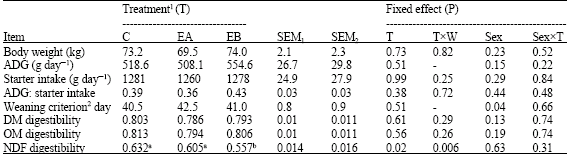 | |
| 1C = Control starter (no enzyme), EA = Enzyme A supplemented starter with respective activity of 1437, 788 and 7476 μmol/mL/min for exo-cellulase, end-ocellulase and xylanase. EB = Enzyme B supplemented starter with respective activity of 1446, 1350 and 5091 μmol/mL/min for exo-cellulase, endo-cellulase and xylanase. 2Calf age at a daily intake of 680 g starter (NRC, 2001). Male calves had significantly lower weaning criterion than did female calves (40.5±0.7 vs. 42.8±0.6 day). SEM1 = standard error of least square means for C and EA; SEM2 = Standard error of least square means for EB. T = Treatment, W = Week | |
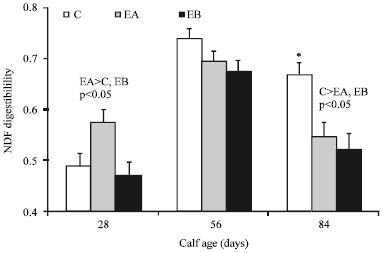 | |
| Fig. 1: | The apparent total tract NDF digestibility. C = Control starter with no enzymes, EA = starter with enzyme additive A, EB = Starter with enzyme additive B. The respective activity (μmol/mL/min) of exo-cellulase, endo-cellulase and xylanase were 1437, 788 and 7476 for EA and 1446, 1350 and 5091 for EB. No enzymes were added to the post-weaning starter |
Adding ENP to the calf starter did not influence the apparent total tract digestibility of Dry Matter (DM) and Organic Matter (OM) either in pre-weaning or in post-weaning calves (p>0.55, Table 2). Applying EA to the calf starter significantly enhanced (p<0.05) the apparent total tract NDF digestibility at week-4 but not at week-8, compared to EA-fed and control calves (Fig. 1). The apparent total tract NDF digestibility at week-12 was, however, lower (p<0.05) for calves fed ENP-treated, pre-weaning diet than for calves fed control, pre-weaning diet (Fig. 1). Such post-weaning decline in fiber digestibility led to an overall decline (p<0.05) in total tract NDF digestibility in response to EB; whereas EA-fed and control calves had comparable NDF digestibility.
As a result of similar starter intake and growth rate, the weaning criterion or the age at a daily intake of 680 g starter concentrate was equal among treatments (p = 0.51). Not unexpectedly, male calves had lower weaning criterion than female calves (p<0.05, Table 2). The between-sex difference in weaning criterion became apparent by fitting the polynomial regression equation for calf starter intake against calf age.
DISCUSSION
Unlike no impacts of ENP on starter intake across the 4-week periods in the current study (p>0.05; Table 2), recently Naserian et al. (2005) reported a reduction in DMI of post-weaning calves fed an enzyme supplemented, growing diet. The enzyme additive used by Naserian et al. (2005) was a mixture of cellulase, β-glucanase, α-amylase, protease, pectinase and phytase; whereas the enzyme additives used in our study contained only cell-wall carbohydrases. Additionally, unlike this study, Naserian et al. (2005) applied the exogenous enzymes to post-weaning diet and monitored the calf performance from 60 to 120 day of age. Such discrepancies in enzyme type and calf age may explain the different results between the two studies. Using beef cattle fed barley grain-based diets, Beauchemin et al. (1997) observed no effects of ENP on DMI. McAllister et al. (1999), on the other hand, reported a quadratic DMI response to ENP in backgrounding steers fed a barley silage-based diet. The non-linear response in DMI might indicate a more pronounced effect of ENP on less digestible, forage-based diets that in turn causes a faster ruminal outflow rate and greater feed intake response, as pointed out by Beauchemin et al. (2003) too. After all, the results of our study using young calves with those obtained using cattle already possessing a well-developed reticulo-rumen should be compared cautiously.
Similar to the present study, Naserian et al. (2005) found no effects of exogenous enzymes on post-weaning ADG of calves. The same ADG response could mean that the effect of ENP on nutrient utilization was not significant enough to cause a difference in pre-weaning calf growth. The early increase in fiber digestibility by EA may be due to the more transitory state of reticulo-rumen development at week-4 leading to a sharper reaction to ENP, when compared with week-8. The greater response in fiber digestibility to EA than to EB may also be attributable to the higher xylanase activity in EA than in EB (Materials and Methods). Introducing ENP into the rumen of sheep as compared to its prefeeding addition to barley silage reduced DM and NDF digestibility (McAllister et al., 1999). The depressed fiber digestibility might then suggest a contribution from competitive microbial interactions in the rumen. Noteworthy, we took spot fecal samples and used AIA as an internal marker to measure nutrient digestibility. Spot sampling may not realistically represent the continuous flow of excreta along the gastric tract. Hence, it would be suggested that future studies use other techniques preferably total fecal collection, if feasible experimentally. The equal BW and ADG of calves on control and ENP supplemented starters could indicate that the fiber amount utilized by 12-week-old calves was equally supportive to complement nutrient supply for growth among the treatments. Importantly, calves used in the present study trial achieved a daily starter intake of 680 g, recommended by NRC (2001), at approximately 41-42 days of age. Considering the conventional weaning age of 70-90 day in majority of dairy farms in Iran, results indicate that a lower weaning age could be considered where high-quality starch and protein sources are available.
CONCLUSIONS
Applying certain type of ENP (EA) to pre-weaning starter concentrate enhanced apparent total tract NDF digestibility at week-4 of age but not later. The NDF digestibility at week-12 was however lower in calves receiving ENB in pre-weaning starter than in calves fed EA-supplemented and control starters before weaning. Starter intake and growth rate were not affected by ENP neither pre-weaning nor post-weaning. As a result, the daily intake of 680 g starter diet was achieved at comparable ages among treatments. Further work preferably with a mixture of cell-wall and cell-content polysaccharidases is warranted if the commercial use of ENB in calf starter is to be decisively assessed.
ACKNOWLEDGMENTS
This study was funded by Isfahan University of Technology. The management board at Foka Dairy Farm is thanked for hosting the calf trial and Dr. Gary Crow (Department of Animal Science, University of Manitoba) is acknowledged for discussing the statistics.
REFERENCES
- AOAC, 1990. Official Methods of Analysis. 15th Edn., Association of Official Analytical Chemists, Washington, DC., USA., pp: 200-210.
Direct Link - Beauchemin, K.A., L.M. Rode and V.J.H. Sewalt, 1995. Fibrolytic enzymes increase fiber digestibility and growth rate of steers fed dry forages. Can. J. Anim. Sci., 75: 641-644.
CrossRefDirect Link - Beauchemin, K.A., L.M. Rode and D. Karren, 1999. Use of feed enzymes in feedlot finishing diets. Can. J. Anim. Sci., 79: 243-246.
Direct Link - Beauchemin, K.A., D. Colombatto, D.P. Morgavi and W.Z. Yang, 2003. Use of exogenous fibrolytic enzymes to improve feed utilization by ruminants. J. Anim. Sci., 81: E37-E47.
Direct Link - Beauchemin, K.A., D. Colombatto, D.P. Morgavi, W.Z. Yang and L.M. Rode, 2004. Mode of action of exogenous cell wall degrading enzymes for ruminants. Can. J. Anim. Sci., 84: 13-22.
Direct Link - McAllister, T.A., S.J. Oosting, J.D. Popp, Z. Mir and L.J. Yanke et al., 1999. Effect of exogenous enzymes on digestibility of barley silage and growth performance of feedlot cattle. Can. J. Anim. Sci., 79: 353-360.
Direct Link - NRC, 2001. Nutrient Requirements of Dairy Cattle. 7th Edn., National Academy Press, Washington, DC, ISBN: 978-0-309-06997-7, Pages: 405.
CrossRefDirect Link - Van Keulen, J. and B.A. Young, 1977. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci., 44: 282-287.
CrossRefDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Wang, Z. and L.A. Goonewardene, 2004. The use of mixed models in the analysis of animal experiments with repeated measures data. Can. J. Anim. Sci., 84: 1-11.
Direct Link