
A. Cheikhyoussef
Food Science and Technology, Southern Yangtze University
N. Pogori
Laboratory of Brewing Microbiology and Applied Enzymology,
School of Biotechnology, Key Laboratory of Industrial Biotechnology, Ministry of Education,
School of Biotechnology, Southem Yangtze University,
170 Huihe Rd, Wuxi 214036, Jiangsu, Peoples Republic of China
H. Zhang
Food Science and Technology, Southern Yangtze University
International Journal of Dairy Science
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 116-125







ABSTRACT
The antimicrobial effects of the supernatants obtained from four strains of bifidobacteria were tested against two types of indicator strains, Bacillus cereus and three strains of Escherichia coli. E. coli AS 1.543 was the most sensitive strain among the E. coli strains whereby B. bifidum showed the highest antimicrobial activity against indicator strains followed by B. infantis then B. longum and finally B. adolescentis. There was a significant reduction in the growth of the indicator strains, whereby all E. coli strains were inhibited more than 75% of their log CFU in monoculture, of which B. infantis reduced 95% of E. coli AS 1.543. On the other hand only more than 60% of the log CFU of B. cereus was reduced by all bifidobacterial supernatants in particular B. longum (75%). It was observed that substances or factors other than organic acids may contribute to the antimicrobial activity of the supernatants (CFS) from the bifidobacteria studied. Neutralization of the CFS from B. adolescentis and B. bifidum significantly reduced the antimicrobial activity while that from B. infantis and B. longum continued to inhibit growth of the indicator strain as observed by a clear defined zone uncharacteristic of acid production by the agar diffusion method. The inactivation of the antimicrobial activity of CFS from B. infantis and B. longum by proteolytic enzymes confirms the proteinaceous nature of the antimicrobial compound present. In addition their heat and pH stability further proves the presence of a proteinaceous antimicrobial compound which could be designated as bacteriocins or bacteriocin-like compounds.
PDF Abstract XML References
How to cite this article
A. Cheikhyoussef, N. Pogori and H. Zhang, 2007. Study of the Inhibition Effects of Bifidobacterium Supernatants Towards Growth of Bacillus cereus and Escherichia coli. International Journal of Dairy Science, 2: 116-125.
DOI: 10.3923/ijds.2007.116.125
URL: https://scialert.net/abstract/?doi=ijds.2007.116.125
DOI: 10.3923/ijds.2007.116.125
URL: https://scialert.net/abstract/?doi=ijds.2007.116.125
INTRODUCTION
Bifidobacteria are dominant in the human colon of healthy humans and constitute the main microbial population in breast-fed infants (Trejo et al., 2006). Bifidobacteria benefit the health of the host by balancing the intestinal microbiota, by fermenting oligosaccharides that are not digested neither absorbed in the upper gastrointestinal tract and by displaying adverse effects on other bacteria (De Vuyst et al., 2004).
Several mechanisms have been suggested for the inhibitory action of bifidobacteria towards pathogens, including a decrease of the local pH by the production of organic acids, the inhibitory action of undissociated organic acid molecules, the competition for nutrients, the competition for adhesion sites, the stimulation of the host’s immunity and the production of specific antibacterial substances like bacteriocins (Fuller, 1989; Ballongue, 1998; De Vuyst et al., 2004).
Organic acids, in particular acetic acid and lactic acid, have a strong inhibitory effect against pathogenic bacteria especially Gram-negative bacteria. Some authors suggest that the production of organic acids is the sole factor responsible for the antagonistic activity of bifidobacteria (Ibrahim and Bezkorovainy, 1993; Bruno and Shah, 2002), while other reports have been attributed it to the production of proteinaceous agents in addition to the pH-reducing effects (Geis et al., 1983; Anand et al., 1985; Meghrous et al., 1990; Gibson and Wang, 1994; Yildirim and Johnson, 1998; Collado et al., 2005).
Bacillus cereus is an important food pathogen. This organism is often isolated from food samples, such as raw milk, dairy products and rice products, as a contaminant (Larsen and Jørgensen, 1997). There are two principal types of food poisoning caused by B. cereus, namely diarrheal and emetic types of food poisoning. Diarrheal type of enterotoxin is commonly associated with dairy products while some emetic outbreaks have been transferred via cooked rice (Kramer and Gilbert, 1989).
Raw milk can also be a source of infections caused by Escherichia coli serotype O157:H7 (E. coli O157). The ability of E. coli O157 to survive in fermented dairy products made from raw milk is of major concern because the consumption of such products has also led to infections in humans (Maher et al., 2001).
Bifidobacteria have the ability to suppress the growth of pathogenic bacteria by producing organic acids (Ibrahim and Bezkorovainy, 1993) and therefore lowering the pH of the surrounding environment. But there are some reports which mention the tolerance of pathogenic E. coli strains for low pH (Glass et al., 1992), survival in foods of low pH (Zhao and Doyle, 1994) and resistance to the lethal effects of very low pH. Since bifidobacteria have been found to have the ability to produce antimicrobial compounds such as bacteriocins (Gibson and Wang, 1994; Yildirim and Johnson, 1998; Ibrahim and Salameh, 2001), these bacteria can thus serve to inhibit these acid-tolerant pathogenic bacteria.
The objective of this study was to examine four strains of bifidobacteria (B. adolescentis, B. bifidum, B. infantis and B. longum) for their antimicrobial activity toward harmful bacteria such as Bacillus cereus and three Escherichia coli strains (E. coli TG1, E. coli DH5α and E. coli AS 1.543) and to elucidate that the antimicrobial activity of these Bifidobacterium strains is not only related to the production of organic acids but also to proteinaceous agents which could be designated as bacteriocins or bacteriocin-like compounds.
MATERIALS AND METHODS
Bacterial Strains and Culture Media
Bacterial strains used in this study and their origin are listed in Table 1. The bifidobacterial strains were activated in MRS broth (De Man et al., 1960) obtained from Merck; (Darmstadt, Germany) supplemented with 0.05% (w/v) L-cysteine-hydrochloride (Cys-HCl, Sigma, St. Louis, MO, USA) and incubated under anaerobic conditions using an atmosphere generation system (GasPak system, Oxoid, Basingstoke, Hampshire, England) at 37°C. All strains were subcultured at least three times prior to the experiments.
The numbers of colony forming units (CFU mL-1) in cultures of bifidobacteria and indicator strains cultivated in their respective media as described in Table 1 were determined by the pour plate method at different time intervals during the 60 h time period. All cultures were inoculated with an inoculum of 1% (v/v). The changes in pH in the cultures of bifidobacteria and indicator strains were also measured every 12 h during the 60 h cultivation time.
| Table 1: | List of microorganisms, sources and their culture conditions |
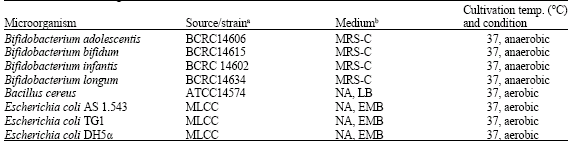 | |
| ATCC: American Type Culture Collection, USA, BCRC: Bioresource Collection and Research Center, Taiwan, MLCC: Microbiology Laboratory Culture Collection, Southern Yangtze University, Wuxi, Jiangsu, China, MRS-C: deMan Rogosa Sharpe agar supplemented with 0.05% (wt/vol) cysteine hydrochloride, NA: Nutrient Agar (pH = 7.2), EMB: Eosin Methylene Blue broth (pH = 6.8) , LB: Luria-Bertani broth (pH = 7.0) | |
Preparation of the Cell-free Supernatants (CFS) from Bifidobacteria
Cell-free supernatants (CFS) of bifidobacterial strains were obtained from 18 h old cultures in MRS broth. The culture broth was centrifuged at 8000 rpm for 20 min at 4°C. The resultant supernatants were then filter-sterilized using a 0.45 μm membrane filter (Millipore Corporation, Bedford MA, USA). Supernatants were then stored at -20°C until further use.
Measurement of Antimicrobial Activity Against Indicator Strains by Turbidimetric Method
The percent inhibition caused by the CFS towards the growth of the indicator strains was determined by measuring the growth (optical density at 600 nm) in monoculture and culture containing the 2% (v/v) CFS.
Measurement of Antimicrobial Activity Against Indicator Strains in Skim Milk Medium
The skim milk medium (12%) (Bright Dairy Company, Shanghai, PRC). The pH of the medium was adjusted to pH 6.60 and then autoclaved at 115°C for 15 min. The skim milk medium was inoculated simultaneously with indicator strain (109 CFU mL-1) and CFS at 1% (v/v). The cultures were then incubated for 48 h at 37°C under aerobic conditions. The numbers of colony forming units (CFU mL-1) were determined every 12 h within a 60 h time period using nutrient agar (Oxoid) supplemented with 10 g L-1 glucose.
Measurement of Antimicrobial Activity Against Indicator Strains by the Agar Diffusion Method
The agar diffusion method according to Tagg et al. (1976) was conducted for the detection of antimicrobial activity against E. coli AS 1.543 and B. cereus with the following modifications: 25 mL soft Nutrient agar (0.7% w/v ) (pH = 6.80) and 25 mL soft Luria-Bertani agar (0.7% v/w ) (pH = 7.00) were seeded with 1% (v/v) of an overnight cultures of E. coli and B. cereus, respectively, poured into sterile Petri plates and allowed to solidify at room temperature. The pH of the CFSs was neutralized by using 5 N NaOH to pH 6.60 (Ivanova et al., 2000). Wells (6mm) were cut in the solidified agar using a sterile metal cork borer and filled with 100 μL of neutralized CFS (pH 6.60). The plates were left at 4°C for 2 h to allow diffusion of the neutralized CFSs after which they were incubated under aerobic conditions at 37°C for 24 h. The presence of inhibition zones was recorded.
Effect of Enzymes and Detergents on the Antimicrobial Activity of the Bifidobacteria
The CFSs were neutralized to pH 6.60 to test only the antimicrobial activity of the inhibitory substance(s) and not that of the presence of organic acids. The CFS was treated with catalase to eliminate the effect of hydrogen peroxide towards the growth of the indicator strains.
The effect of various enzymes (alcalase, diastase, catalase, neutral proteinase, pepsin and trypsin) and detergents (EDTA, sodium dodecyl sulphate and Tween 80) on the antimicrobial activity of neutralized CFS obtained from bifidobacterial strains against E. coli MLS001 was studied in nutrient agar (Oxoid) supplemented with 10 g L-1 glucose at 37°C. The agar diffusion method described by Tagg et al. (1976) was used to test the antimicrobial activity.
To test the effect of enzymes the CFS were incubated for 2 h with 0.10 and 1.0 mg mL-1 final concentration of the following enzymes (Merck, Darmstadt, Germany): Alcalase, diastase, catalase, neutral proteinase, pepsin and trypsin. The control experiment contained no enzymes.
To test the effect of various detergents on the antimicrobial activity, the CFS was incubated for 6 h with 0.10 and 1.0% final concentration of the following detergents: Ethylene Diamine Tetraacetic Acid (EDTA), Sodium Dodecyl Sulphate (SDS) and Tween 80. The control experiment contained no detergent. All experiments were carried out in triplicate.
RESULTS AND DISCUSSION
Growth of Bifidobacteria in Skim Milk and MRS Media
Bifidobacterium strains and indicator strains and their culture conditions which were used in this study are listed in Table 1. The growth of bifidobacteria and indicator strains (CFU mL-1) varied clearly and was depended on the types of media and incubation conditions used (Table 2).
Bifidobacteria obtained highest CFU numbers in skim milk medium after 48 h (Table 2a) and after 36 h in MRS broth (Table 2b). On the other hand, the indicator strains grew faster and caused the spoilage of the skim milk (highest CFU number was detected after 24 h as shown in Table 2c). It was observed that bifidobacteria can decrease the pH (Data not shown) of their culture media more than the indicator strains and this is as a result of their production of organic acids. It is well established that the major metabolites of bifidobacteria are acetic acid and lactic acid in ratio 3:2 and this production is responsible for the associated drop in pH, which may be sufficient to antagonize many pathogenic bacteria belonging to both Gram-positive and Gram-negative bacteria (Rioradan and Fitzgerald, 1998; Makras and De Vuyst, 2006).
| Table 2a: | Growth of the bifidobacterial strains in monoculture in skim milk medium |
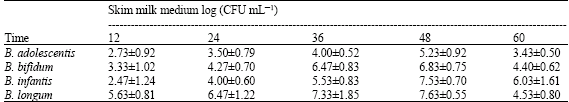 | |
| Table 2b: | Growth of the bifidobacterial strains in *MRS Broth |
 | |
| *MRS: Man Rogosa and Sharpe (De Man et al., 1960) | |
| Table 2c: | Growth of indicator strains in skim milk medium |
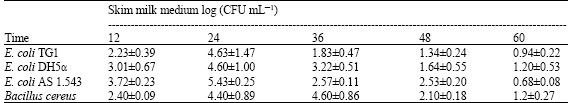 | |
Antimicrobial Activity of Bifidobacteria
The antimicrobial activity of bifidobacterial strains was detected by measuring the optical density of the culture with indicator strains in monoculture and in the presence of 2% of bifidobacterial CFSs in three different media (Table 3a and b).
E. coli AS 1.543 was the most sensitive strain among the E. coli strains whereby B. bifidum showed the highest antimicrobial activity against indicator strains followed by B. infantis then B. longum and finally B. adolescentis (Table 3a and b). These inhibitory effects varied among the different media used whereby E. coli strains were more sensitive in Nutrient Broth (NB) than Eosin Methylene Blue broth (EMB). On the other hand B. cereus was more sensitive in Luria-Bertani broth than in NB.
Antimicrobial Effects of Bifidobacteria Against the Indicator Strains in Skim Milk Medium
There was a significant reduction in the log CFU number of the indicator strains, whereby all E. coli strains were inhibited more than 75% of their log CFU in monoculture, of which B. infantis reduced 95% of E. coli AS 1.543.
| Table 3a: | The growth of indicator strains in monocultures and the percent inhibition of indicator strain populations in Nutrient Broth (NB) by Cell Free Supernatants (CFS) obtained from bifidobacterial cultures |
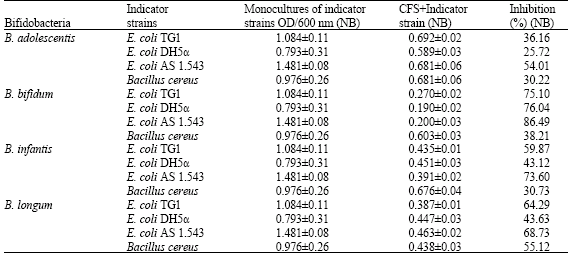 | |
| NB: Nutrient broth (pH = 7.2), Conditions of cultivation: Inoculum of E. coli and B. cereus was 109 CFU mL-1, Incubation at 37°C for 24 h, aerobic conditions | |
| Table 3b: | The growth of indicator strains in monocultures and the percent inhibition of indicator strain populations in EMB and LB media by cell free supernatants (CFS) obtained from bifidobacterial cultures |
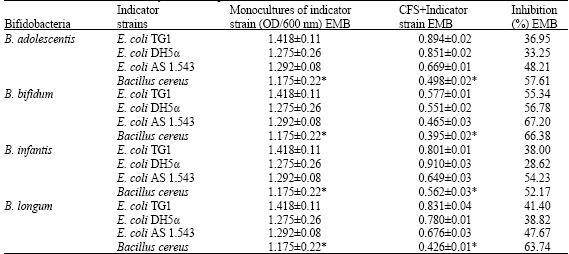 | |
| EMB: Eosin Methylene Blue broth (pH = 6.8); *Luria-Bertani broth (pH = 7.0), Conditions of cultivation: Inoculum of E. coli and B. cereus was 109 CFU mL-1, Incubation at 37°C for 24 h, aerobic conditions | |
| Table 4: | The growth of indicator strains in monocultures and the percent inhibition of indicator strain populations in skim milk medium by cell free supernatants (CFS) obtained from bifidobacterial cultures |
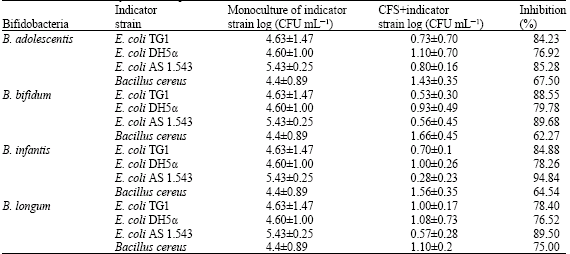 | |
| The skim milk medium was inoculated simultaneously with indicator strain (108 CFU mL-1) and CFS obtained from bifidobacterial cultures. The culture was then incubated for 24 h at 37°C under aerobic conditions | |
On the other hand only more than 60% of the log CFU of B. cereus was reduced by all bifidobacterial supernatants in particular B. longum (75%) (Table 4).
The activity of the CFS was tested also under conditions which eliminate the possible effect of organic acids (lactic and acetic acids). The antimicrobial activity of the supernatants from B. bifidum and B. adolescentis was severely reduced which may explain that their activity is mainly due to the presence of the organic acids. The activity of B. infantis and B. longum supernatants was not affected. In order to determine whether the inhibitory compounds produced by B. infantis and B. longum were other than organic acids such as the possible presence of bacteriocins or bacteriocin-like compounds, the agar well diffusion method was carried out using two indicator strains (E. coli AS 1.543 and B. cereus, Fig. 1 and 2). It was observed that substances or factors other than organic acids may contribute to the antimicrobial activity of the CFS studied. Neutralization of the CFS from B. adolescentis and B. bifidum significantly reduced the antimicrobial activity while that from B. infantis and B. longum continued to inhibit growth of the indicator strain as observed by a clear defined zone uncharacteristic of acid production by the agar diffusion method (Fig. 1 and 2). These results are in agreement with findings by Gibson and Wang (1994), who found that the inhibitory effects of B. infantis against E. coli or Clostridium perfringens were not necessarily related to acid production, but was not in agreement with data obtained by Ibrahim and Bezkorovainy (1993) who concluded that the inhibitory effects of spent bifidobacterial broths towards E. coli were based solely on the reduction in pH. In order to further investigate whether the antimicrobial effect was from the presence of an antimicrobial compound in the CFS, the effect of enzymes and detergents was studied.Effect of Enzymes and Detergents on the Antimicrobial Activity of the Bifidobacteria
The effects observed from different enzymes and detergents on the antimicrobial substances from B. infantis and B. longum were found useful for partially characterizing these substances. There was a partial inactivation of antimicrobial activity of the CFS from B. infantis after treatment with proteolytic enzymes. Complete inactivation or significant reduction in the activity of the CFS from B. longum was observed after treatment with neutral proteinase, alcalase, pepsin and trypsin which indicated the proteinaceous nature of the antimicrobial compound (Table 5). This is in agreement with findings by Collado et al. (2005) who studied the effects of different enzymes on the activity of six bifidobacterial strains of human origin and identified the proteinaceous structure of the antimicrobial compound. Diastase and lipase had no effect which confirmed that carbohydrate and lipid moieties were not required for the inhibitory activity.
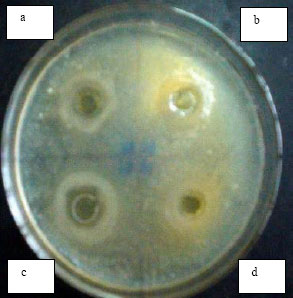 | |
| Fig. 1: | Inhibitory activity of CFS toward E. coli AS 1.543 in Nutrient agar (Agar diffusion method was carried out according to Tagg et al. (1976) whereby wells have been loaded with 100 μL CFS). Inhibitory activity of CFS from B. infantis before (a) and after neutralization (b). Inhibitory activity of CFS from B. longum before (c) and after neutralization (d) |
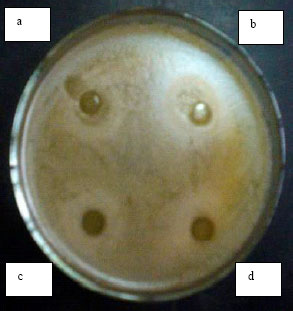 | |
| Fig. 2: | The effect of CFS toward Bacillus cereus in Luria-Bertani agar (Agar diffusion method was carried out according to Tagg et al. (1976) whereby wells have been loaded with 100 μL CFS). Inhibitory activity of CFS from B. infantis before (a) and after neutralization (b). Inhibitory activity of CFS from B. longum before (c) and after neutralization (d) |
This data is in agreement with findings by Yildirim and Johnson (1998) who found that lipids and carbohydrates moieties were absent and not responsible for the inhibitory action of bifidocin B from B. bifidum NCFB1454.
| Table 5: | Effect of enzymes and surfactants on the antimicrobial activity of cell free supernatants (CFS) obtained from Bifidobacterium infantis and Bifidobacterium longum cultures against the growth of Escherichia coli AS 1.543 |
 | |
| + : Complete inactivation, - : No effects, *Partial inactivation, The agar diffusion method described by Tagg et al. (1976) was the used to visualize the antimicrobial activity on nutrient agar | |
Complete inactivation was observed when supernatants from B. longum and B. infantis (Table 5) were treated with detergents (Tween 80, SDS and EDTA). Tween 80 only partially inactivated the activity in supernatants from B. infantis.
Catalase treatment did not affect the antimicrobial activity of B. infantis and B. longum supernatants toward the indicator strain, E. coli AS 1.543, which proved that hydrogen peroxide or organic acids were not the sole reason of the antimicrobial activity. This data is in agreement with findings by Rioradan and Fitzgerald (1998).
Heat treatment showed that antimicrobial compound from B. infantis is more stable than that of B. longum. The antimicrobial compound from B. infantis was found to be stable over a wide range of pH from 4-10 and that of B. longum had pH stability from pH 5-9 (Data not shown). These results are in agreement with findings by Meghrous et al. (1990) who studied the production of bacteriocins by 13 strains of bifidobacteria and found that the crude bacteriocin-like compounds produced were of a protein type, heat-stable and active at pH values ranging from 2 to 10.
The inactivation of the antimicrobial activity of CFS from B. infantis and B. longum proteolytic enzymes confirms the proteinaceous nature of the antimicrobial compound present. In addition their heat and pH stability further proves the presence of a proteinaceous antimicrobial compound. Therefore the compounds can be designated as bacteriocins or bacteriocin-like compounds (Tagg et al., 1976; Collado et al., 2005). Several reports have demonstrated that specific antimicrobial substances are elaborated by members of bifidobacteria, including bifidin, bifidocin and biflong produced by B. bifidum and B. longum strain respectively (Anand et al., 1985; Kang et al., 1989). In addition Meghrous et al. (1990) identified bacteriocin-producing bifidobacterial strains active against certain Gram positive species and Gibson and Wang (1994) provided additional evidence that acidity may not be the sole mechanism of inhibition.
CONCLUSIONS
In this work it was shown that the basic metabolites (acetic and lactic acids) from bifidobacteria are not the only factors affecting the inhibitory performance towards E. coli and Bacillus sp. and that there are compounds present that are of proteinaceous nature which can solely inhibit the indicator strains studied. The CFS (non-neutralized) from bifidobacteria efficiently inhibited the growth of E. coli and Bacillus strains in their optimum culture media including that in skim milk medium. Therefore, the antimicrobial compounds studied here have potential as food safety agents in the dairy products such yogurt and cheese.
ACKNOWLEDGMENTS
The research work was financed by the Key Laboratory of Lactic Acid Bacteria Fermentation Technology and Food Safety, Ministry of Education, Southern Yangtze University, Wuxi, P.R. China.
REFERENCES
- Bruno, F.A. and N.P. Shah, 2002. Inhibition of pathogenic and putrefactive microorganisms by Bifidobacterium sp. Milchwiss, 57: 617-621.
Direct Link - Collado, M.C., M. Hernandez and Y. Sanz, 2005. Production of bacteriocin-like inhibitory compounds by human fecal Bifidobacterium strains. J. Food Prot., 68: 1034-1040.
Direct Link - De Man, J.C., M. Rogosa and M.E. Sharpe, 1960. A medium for the cultivation of Lactobacilli. J. Applied Bacteriol., 23: 130-135.
CrossRefDirect Link - Fuller, R., 1989. Probiotics in man and animals. J. Applied Bacteriol., 66: 365-378.
CrossRefPubMedDirect Link - Geis, A., J. Singh and M. Tueber, 1983. Potential of lactic streptococci to produce bacteriocin. Appl. Envir. Microbiol., 45: 205-211.
Direct Link - Gibson, G.R. and X. Wang, 1994. Regulatory effects of bifidobacteria on the growth of other colonic bacteria. J. Applied Bacteriol., 77: 412-420.
CrossRefDirect Link - Glass, K.A., J.M. Loeffelholz, J.P. Ford and M.P. Doyle, 1992. Fate of Escherichia coli O157:H7 as affected by pH or sodium chloride and in fermented, dry sausage. Applied Environ. Microbiol., 58: 2513-2516.
Direct Link - Ibrahim, S.A. and M.M. Salameh, 2001. Simple and rapid method for screening antimicrobial activities of Bifidobacterium species of human isolates. Rapid Meth. Autom. Microbiol., 9: 52-63.
Direct Link - Ivanova, I., P. Kabadjova, A. Pantev, S. Danova and X. Dousset, 2000. Detection, purification and partial characterization of novel bacteriocin substance produced by Lactococcus lactis subsp. lactis B14 isolated from Boza-Bulgarian traditional cereal beverage. Biocatal. Fundam. Appl., 41: 47-53.
Direct Link - Larsen, H.D. and K. Jorgensen, 1997. . The occurrence of Bacillus cereus in danish pasteurized milk. Int. J. Food Microbiol., 347: 179-186.
PubMedDirect Link - Maher, M.M., K.N. Jordan, M.E. Upton and A. Coffey, 2001. Growth and survival of Escherichia coli O157:H7 during the manufacture and ripening of a smear-ripened cheese produced from raw milk. J. Applied Microbiol., 90: 201-207.
CrossRefDirect Link - Makras, L. and L. De Vuyst, 2006. The in vitro inhibition of Gram-negative pathogenic bacteria by bifidobacteria is caused by the production of organic acids. Int. Dairy J., 16: 1049-1057.
CrossRefDirect Link - Rioradan, K.O. and G.F. Fitzgerald, 1998. Evaluation of bifidobacteria for the production of antimicrobial compounds and assessment of performance in cottage cheese at refrigeration temperature. J. Applied Microbiol., 85: 103-114.
CrossRef - Tagg, J.R., A.S. Dajani and L.W. Wannamaker, 1976. Bacteriocins of gram-positive bacteria. Bacteriol. Rev., 40: 722-756.
CrossRefPubMedDirect Link - Trejo, F.M., J. Minnaarda, P.F. Pereza and G.L. De antoni, 2006. Inhibition of Clostridium difficile growth and adhesion to enterocytes by Bifidobacterium supernatants. Anaerobe, 12: 186-193.
CrossRef - Yildirim, Z. and M.G. Johnson, 1998. Characterization and antimicrobial spectrum of bifidocin B, a bacteriocin produced by Bifidobacterium bifidum NCFB 1454. J. Food Prot., 61: 47-51.
PubMedDirect Link