
Naheed Mojgani
Depa1tment of Biotechnology, Razi Vaccine and Serum Research Institute
Cyrus Amirnia
National Research Institute of Animal Sciences, Islamic Republic of Iran
International Journal of Dairy Science
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 1-12







ABSTRACT
L. casei RN 78 isolated from a dairy sample was shown to produce a proteinaceous substance inhibitory towards a number of gram positive bacteria including an important food-borne pathogen L. monocytogenes. The inhibitory substance produced by this strain appeared not related to organic acids and hydrogen peroxide and was found resistant to catalase, lipase and lysozyme. The proteinaceous antagonistic nature of the agent appeared to be related to a bacteriocin and was termed Lactocin RN 78. The stability of the crude Lactocin RN 78 in the presence of different physical and chemical factors was evaluated which indicated its stability at wide temperature and pH range. The spectrum of inhibition of this bacteriocin was further enhanced to the growth of E. coli, Ps. aeruginosa, and S. typhi in the presence of 1% chelating agents like SDS and EDTA. Lactocin RN 78 appeared stable in the presence of high concentration of NaCl and resisted 40% of this salt during 48 h of incubation at room temperature. The kinetics of Lactocin RN 78 production by L. acidophilus RN 78 as a function of cellular condition was evaluated, which indicated a direct relationship between the growth rate (cfu mL-1) and the amount of bacteriocin produced. The synthesis and inhibitory activity started in the early logarithmic phase and continued to the end of exponential phase. The growth became static after entering the stationary phase within 10 h of incubation at 35°C and the antibacterial titer remained constant thereafter. The influence of variable physical and chemical agents on the growth metabolism and Lactocin RN 78 synthesis together was investigated. The optimal temperature and pH favoring maximum growth and production in the producer strain was 35°C and 6.0, respectively. The producer strain was able to grow and produce Lactocin RN 78 in the presence of 3% NaCl and 0.3% bile salts. The growth medium most effective was MRS broth medium supplemented with a carbon source.
PDF Abstract XML References
How to cite this article
Naheed Mojgani and Cyrus Amirnia, 2007. Kinetics of Growth and Bacteriocin Production in L. casei RN 78 Isolated from a Dairy Sample in IR Iran. International Journal of Dairy Science, 2: 1-12.
DOI: 10.3923/ijds.2007.1.12
URL: https://scialert.net/abstract/?doi=ijds.2007.1.12
DOI: 10.3923/ijds.2007.1.12
URL: https://scialert.net/abstract/?doi=ijds.2007.1.12
INTRODUCTION
Lactic Acid Bacteria (LAB) have traditionally been used in food processing because of their characteristic flavor changes but mainly because of their ability to lower the pH and to produce antimicrobial agents (Delves-Broughton, 1990; Brink et al., 1994; Minahk et al., 2004). The antagonistic activity possessed by these bacteria can be due to organic acids, diacetyl, acetic acid, hydrogen peroxide and carbon dioxide production alone or in combination and bacteriocins (Tagg et al., 1976; Piard and Desmazeaud, 1992; Casla et al., 1996; Coventry et al., 1996). In the last decades a vast interest has been developed in research on bacteriocins especially for exploiting them in dairy and other edible products as a natural biological preservative due to their important role of inhibiting pathogenic or spoilage bacteria through many mechanisms (Piard and Desmazeaud, 1991, 1992). In addition, the use of chemical preservatives has fallen into disfavor with consumers, who are requesting fresh, natural food products that are mild and light, with less acid, sugar, salt, or fat (Gould 1996; Lucke, 2000).
Bacteriocins are bioactive peptides produced by certain gram positive and gram negative bacteria which are inhibitory towards other closely related bacteria (Lewus and Montville, 1991; Itoh et al., 1995; Cleveland et al., 2001). A number of bacteriocins from Lactic acid bacteria have been studied In detail including the ones produced by L. casei, L. acidophilus, L. rhamnosus, L. reuteri, L. salivarius, L. plantarum, L. bulgaricus etc (Muriana and Klaenhammer, 1991; Itoh et al., 1995, Nam et al., 2002). The determination of optimum parameters for both enhanced production and purification of bacteriocins is amongst the prerequisites for their use in the food, veterinary and pharmaceutical industries. Thus, before these bacteria and their products could find their way into the commercial market a number of stress factors are to be studied on these bacteria so as to determine their tolerance and survival after being ingested by the consumer. In the present study we investigated the effect of different pH, temperature, salts and media composition on the growth and Lactocin RN 78 synthesis in L. acidophilus. To determine the extent of survival and tolerance of the bacteriocin produced a growth curve was plotted by using biomass (OD 655 nm and cfu mL-1) and bacteriocin production (inhibitory activity in mm) as objective variables.
MATERIALS AND METHODS
Bacterial Strains and Culture Conditions
The bacteriocin producing Lactobacillus casei RN 78 strain isolated from a dairy sample (Mojgani et al., 2005)), was propagated and cultured in MRS broth (De Man Rogosa and Sharpe, Oxoid), at 35-37°C, anaerobically in Oxoid anaerobic jars. The indicator organism used was Listeria monocytogenes ATCC 1315, that was propagated and maintained in Brain heart infusion broth (BHI, Difco), at 37°C for 24 h. Agar plates were prepared by adding 1.6% agar while soft agar was prepared by the addition of 0.8% agar to the respective broth medium.
The producer culture was maintained as frozen stock (-20°C) in MRS broth medium supplemented with 20% glycerol. The frozen stock cultures were thawed and propagated twice at 35°C in MRS broth supplemented with 5% horse serum.
Detection of Inhibitory Activity
To determine the antimicrobial activity of the bacteriocin produced by L.casei RN 78, agar-well diffusion, Spot-on lawn assay and paper disc diffusion methods were used with slight modifications (Toba et al., 1991; Apella et al., 1992). The MRS agar plates were overlaid with 5 mL of BHI soft agar having 10 μL of overnight indicator culture. Overnight producer cultures were centrifuged at 12,00 rpm, 4°C for 20 min and cell-free supernatant fluids were neutralized by 2 M NaOH. Wells of 5 mm were punched into the overlaid plates and 100 μL of the neutralized supernatant broth of the producer strain were added to each well. For spot-on lawn assay, 10 μL of the neutralized supernatant fluid, while in paper disc assay the supernatant impregnated paper discs were placed gently on the top of indicator lawn. All the plates were incubated at 4°C for 2-4 h prior to incubation at 37°C for 18-24 h. The zones of inhibition formed around the wells were measured in millimeter. The results were recorded as average of three experiments.
Physico-Chemical Characterization of Antimicrobial Substance
The effect of different enzymes, temperature, pH, NaCl, detergents and organic solvents were investigated on the antimicrobial activity of the biomolecule produced by the producer strain, by agar-well diffusion assay.
Effect of Enzymes
To study the effect of trypsin, proteinase K, catalase, lipase and lysozyme (Sigma) on the antagonistic activity, the supernatant fluid was incubated with enzyme solutions prepared to a final concentration of 1 mg mL-1 in phosphate buffer saline (pH 7.0). The activity was determined after 1, 4 and 6 h by agar-well diffusion assay.
Effect of Temperature
The stability of the crude bacteriocin produced by L. casei RN 78 was investigated by incubating the neutralized cell free extract of the producer strain at room (37°C), refrigerated (4°C) and freezing (-20°C) temperatures. After every 15 days the sample were taken and the activity recorded by agar-well diffusion assay.
Effect of pH
Influence of pH on the stability of the antagonistic molecule was investigated by adjusting the pH of the supernatant fluid from 1-11 with 3 M NaOH or 4 M HCl. After variable time intervals the remaining activity was recorded by the mentioned method.
Effect of NaCl
Increasing concentrations of NaCl ranging from 10 to 40% w/v were added to bacterial extracts and incubated at 35°C. Activity was recorded after 4, 12, 24 and 48 h by the method described earlier. Negative control consisted of 40% NaCl in phosphate buffer saline (pH 7.0).
Effect of Detergents
Neutralized cell free fluids were incubated with SDS and EDTA at a final concentration of 1% w/v and the activity determined after 4, 12, 24 and 48 h as earlier. Negative control consisted of 1% of the respective detergent in phosphate buffer saline (pH 7.0)
Effect of Organic Solvents
The effect of 10, 20, 30, 40 and 50% v/v of methanol, ethanol, propanol and butanol was studied on the bacteriocin extract. Results were recorded after every 4, 12, 24 and 48 h as before. As negative control 50% of the respective solvents in PBS (pH 7.0) were used.
Growth Kinetics of L. acidophilus RN 78 and Lactocin RN 78 Synthesis
The effect of various physical and chemical parameters was studied on the growth and lactocin production of L. acidophilus RN 78 which are as follows:
Influence of Temperature
The effect of four different temperatures 4, 22, 35 and 42°C were tested on the producer strain L. casei RN 78. A fixed amount of producer strain (10 μL) was added to MRS broth (5 mL, pH 5.5) at OD (655 nm) equal to 0.2 and incubated at the said temperatures anaerobically. Samples were drawn aseptically after every half an hour and monitored for absorbency at 655 nm and total viable count (cfu mL-1). The bacteriocin activity was determined by agar-well diffusion method described earlier and a growth curve plotted against OD or cfu mL-11 and antimicrobial activity (mm).
Influence of pH
The pH of the MRS broth was adjusted to 3, 5, 7, 9 and 11 prior to inoculation of the bacterial strain, by addition of 3 M NaOH or 2 M HCl solution. After seeding the broth medium with the test culture the flasks were incubated at 35°C and samples were monitored for viable count and antibacterial activity as above.
Influence of NaCl
Different concentrations of sodium chloride (Sigma) ranging from 0 to 5% w/v were added to MRS broth medium (pH 5.5) and the test culture added. Similarly the growth rate and bacteriocin activity was followed as before.
Influence of Bile Salt
Different concentrations of bile salt (Oxgall) i.e., 0.1, 0.2, 0.3 and 0.5% w/v was added to the broth medium (pH 5.5) before inoculation of the test strain. After 24 h of incubation at 35°C all the tests for determining the growth rate and activity were performed.
Influence of Media Composition
The effect of different media including MRS (HiMedia, India), MRS supplemented with 0.5% lactose, Triptic Soya broth (TSB, Oxoid) and Skim milk (Oxoid) was studied on the growth and production of lactocin RN 78 at 35°C in anaerobic conditions. All the samples were drawn aseptically and tested for their viable count and bacteriocin activity.
RESULTS
The inhibitory action of the cell free supernatant fluid of L. casei RN 78 was studied against a number of gram positive bacteria of which L. monocytogenes was the most strain and was thus used through out the studies as sensitive indicator strain. Compared to spot-on lawn and paper disc assay, the inhibitory activity demonstrated by the producer strain was best studied by the agar-well diffusion assay and was thus selected as the method of choice during the study (Fig. 1).
The chemical nature of the antagonistic molecule, different from lactic acid or hydrogen peroxide was proved because neither the cell free supernatant at pH 7.0 nor the catalase treatment reduced the antibacterial effect observed for the non-treated supernatant. However, the antimicrobial substance was inactivated within two h of incubation in the presence of 1 mg mL-1 of proteolytic enzymes. Lipolytic and glycolytic enzymes appeared to be ineffective at the same concentrations (Table 1).
The stability study of crude Lactocin RN 78 in different physico-chemical conditions indicated the resistance of this agent. The antimicrobial peptide in study was able to resist most of these factors to which it was exposed even during prolong incubation period. Figure 2 depicts the stability of this bacteriocin at different temperatures. Lactocin RN 78 was most stable at freezing temperatures and was able to retain its activity for 2 years without any decrease. At 4°C the bacteriocin lost its activity completely after 11 months of incubation and at 37°C it was active for only a week.
The bioactivity of Lactocin RN 78 was stable at a wide pH range of 3 to 10 when incubated in these conditions at optimum temperatures. Maximum activity of this bacteriocin was recorded at alkaline pH values exceeding pH 8.0.
In the presence of 1 to 40% NaCl, the bacteriocin retained its activity during 48 h of incubation at 37°C. However during prolonged incubations with the said concentrations of the salt Lactocin RN 78 lost its activity significantly. Complete loss of activity of the bacteriocin was observed after 45 days of incubation in the presence of 10% NaCl (Fig. 3).
The antibacterial activity of lactocin RN 78 was greatly enhanced in the presence of variable concentrations of the organic solvents in study. The bacteriocin was tolerant to butanol and resisted 50% of the solvent during the given time period. However, it was comparatively less resistant to methanol, ethanol and propanol and complete loss of activity in the presence of 20, 30 and 50% of these solvents was recorded.
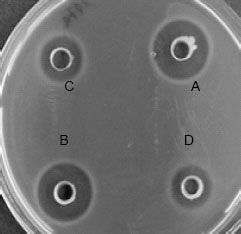
Figure 4, the chelating agents used in the study were able to enhance the antimicrobial activity of the bacteriocin against the indicator organism. Lactocin RN 78 was not only able to resist the detergents for more than 48 h but also enhanced the inhibitory action against E. coli, P. aeroginosa and S. typhi to which it was previously ineffective.
The growth metabolism and the synthesis of bacteriocin in L. casei RN 78 were controlled by plotting a growth curve. The time course required for the growth and bacteriocin synthesis in is shown in Fig. 5. The growth curve depicts the correspondence of cell density to lactocin RN 78 production in the producer strain and higher the growth greater the synthesis of antagonistic compound seen.
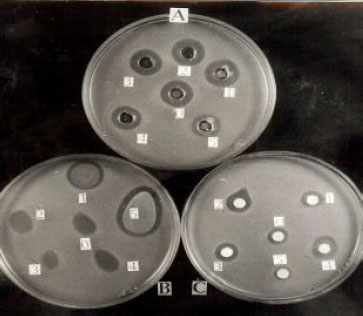 | |
| Fig. 1: | Antagonistic activity demonstrated by Lactocin RN 78 determined by three different methods. A: Agar-well diffusion assay; B: Spot-on lawn assay: C: Paper disc assay |
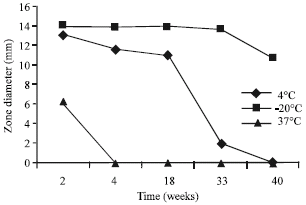 | |
| Fig. 2: | Stability of Lactocin RN 78 during prolong incubation at different temperatures |
 | |
| Fig. 3: | Effect of different percentage of NaCl on the bioactivity of Lactocin RN 78 during 1 week of incubation at room temperature |
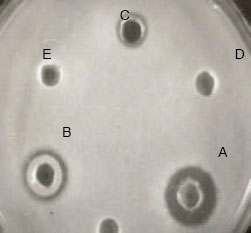 | |
| Fig. 4: | Inhibitory activity of the Lactocin RN 78 against S. typhi in the presence of 1% SDS and EDTA. The activity was checked after 4(A), 12(B) and 24(C) h. D and E are negative control |
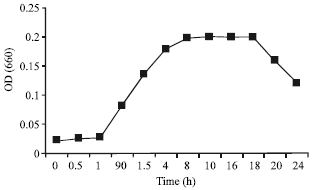 | |
| Fig. 5: | Growth metabolism of L. casei RN 78 at optimum temperature and pH conditions |
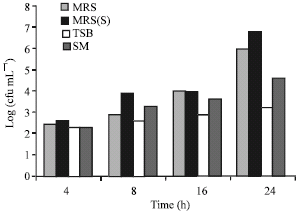 | |
| Fig. 6: | Effect of four different growth media (MRS, MRSs supplemented with lactose, TSB and Skim milk media) on the growth and production of producer strain |
| Table 1: | Effect of enzymes on the antagonistic activity of Lactocin RN78 |
 | |
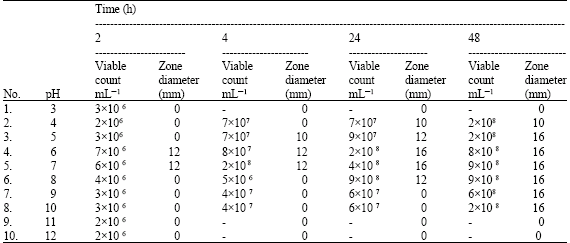
| Table 2: | Effect of pH on growth and Lactocin synthesis in L.casei RN 78 strain |
 | |
It may be noted that maximum growth of L. casei was observed after 4 h of anaerobic incubation at 35°C. The growth reached at maximum and a peak was reached within 12 h of incubation at the optimum temperature. The stationary phase was achieved after this time period where the growth and production remained constant for a period of time before declining. Both the parameters of growth and production was adversely effected at temperature above 40°C and a prolonged lag and a delayed log phase was observed at these temperatures. Similarly, minimum growth and ultimately minimum activity was recorded at low temperatures of 4°C and lower.
As Table 2 shows the bacteria in study was able to grow and produce lactocin RN 78 at pH values ranging from 4.0 to 9.0, while maximum growth and production was at pH values of 5.0 and 6.0 only. A delay in the log phase was observed at highly acidic or alkaline pH values i.e., at pH values lower than 4.0 and above 7.0. The growth at these pH values were recorded late after 18 h of incubation. An important observation made was the minimum rate of growth of the test strain during the initial incubation period (prior to 18 h) at pH 9.0, however after relapse of this time period there was an sudden drift in the growth rate and it accelerated significantly reaching its maximum within 24 h. During the shift, a sharp decline in the pH from 9.0 to 6.0, was recorded which might be the reasons for the growth accelerations.
A pronounce effect of NaCl and bile salts was observed on the growth and Lactocin RN 78 production. L. casei RN 78 growth was stable in the presence of 3% NaCl and 0.3% bile salts in 24 h. However above concentrations of these salts adversely effected the growth and production and delayed the natural lag phase from 4 to 8 h. Significant amount of growth and maximum production was observed after 15 h of incubation in the presence of above concentrations.
Figure 6 shows the effect of media on the criteria tested. The growth media favoring maximum growth and production of lactocin RN 78 was MRS broth media supplemented with 0.5% lactose. Skim milk favored only the growth of the test strain while had an adverse effect on the production ability and no inhibitory activity was observed in any of the growth phases.
DISCUSSION
The physico-chemical characterization of the antimicrobial substance produced by L. casei RN 78 revealed it to be proteinaceous and unrelated to acids and hydrogen peroxide or bacteriophages (Mojgani et al., 2005). Like other known bacteriocins of lactobacilli, the inhibitory activity exhibited by this isolate was inactivated by treatment with proteolytic enzymes. These observations suggested the proteinaceous nature of the inhibitor in study. Due to all these characteristics this substance was considered to be a bacteriocin or a bacteriocin like substance (Tagg et al., 1976; Piard and Desmazeaud, 1992; Jack et al., 1995; Audisio and Apella, 2006).
As Hardy (1975) had mentioned bacteriocins may be either simple protein or protein linked to lipid and carbohydrate. During the course of study it was observed that carbohydrate and lipid moiety were not critical for the activity in L. acidophilus RN 78 and they appeared simple protein molecule rather than conjugated. These results coincide with those of other authors who have reported bacteriocin production by strains of L. acidophilus (Barefoot and Klaenhammer, 1983; Muriana and Klaenhammer, 1991; Avonts et al., 2004).
An important factor that is to be considered while selecting a suitable bacteriocin for use as a preservative is its stable nature. Thus it is considered essential to study temperature stability, pH resistance, salt tolerance, heavy metals, organic solvents and effect of other food components, on the bacteriocin (Larsen et al., 1993). In our studies, Lactocin RN 78 appeared to be relatively a stable protein being active for 11 months at refrigerated temperature and more than two years during freezing temperatures. This property of lactocin RN 78 is a highly desirable factor especially for its future exploitation in food industries as a biological preservative.
An interesting fact revealed during the studies was the alkaline pH tolerance of the bacteriocin in study. These results were not coherent with the previous reports made by Bhunia et al. (1991) and Yang et al. (1992), who had indicated the tolerance of bacteriocins to acidic pH rather than alkaline. However, similar to Lactocin RN 78, listeriocin and perfringens, two bacteriocins produced by L. monocytogenes and C. perfringens, respectively, have been reported to be alkaline pH tolerant (Tagg et al., 1976).
Sodium chloride, an important constituent of food has been reported to affect the efficacy of bacteriocin against target microorganisms. Harris et al. (1989) had provided an evidence for this statement, as he noticed a decrease in survival rate of L. monocytogenes in the presence of nisin with sodium chloride. We here observed the stability of Lactocin RN 78 to this salt at concentrations as high as 40% w/v, which could be considered an advantage for its probable use in vegetable and vegetable juice fermentation. Moreover, the salt tolerance of lactocin RN 78 has additional advantage especially in food industries where NaCl is used as a chemical preservative.
Since the spectrum of activity of some of the LAB bacteriocins are considerably enhanced by combination with chelating agents, there is considerable interest in using bacteriocins in current and potential applications in the veterinary and pharmaceutical areas (Delves-Broughton et al., 1996). Some potential medical use of these bacteriocins in the presence of a chelator has been reported for nisin and pediocin, a bacteriocin produced by L. lactis and P. acidilactic. These bacteriocins in combination with EDTA showed good potential as a therapeutic agent in treatment of human ulcer disease caused by H. pylori, Bovine mastitis caused by S. aureus and in other systemic infections (Blackburn et al., 1989; Howell et al., 1993; Severina et al., 1998). Coherent results were obtained during these investigations and lactocin RN 78 showed enhanced inhibitory activity after treatment with 1% SDS and EDTA. After exposure the bacteriocin was able to inhibit the growth of some gram-negative organisms to which they were previously unaffected. As reported by others, the enhanced inhibitory action of bacteriocin after treatment with EDTA and SDS could be due to the ability of these agents to disperse the proteinaceous complex from its present form (large subunit) into smaller more active lethal units.
Similar to plantaricin LC 74, a bacteriocin produced by L. plantaricin LC74, lactocin RN 78 was found to be stable after treatment with different concentrations of organic solvents like butanol, ethanol and methanol. The stable nature of these bacteriocins in the presence of organic solvents also indicates their proteinaceous and soluble nature (Rekhif et al., 1994).
Among the factors well documented to effect the growth and production are the growth medium, temperature, pH, EDTA and sodium chloride, etc (Larsen et al., 1993).
The growth and bacteriocin-producing ability of L. casei RN 78 was characterized under the influence of some of these factors. The main criteria used were temperature, pH, salt, detergents and organic solvents. Lactocin RN 78 synthesis appeared similar to amylovorin L471, a bacteriocin produced by L. amylovorous (De Vuyst et al., 1994) as the growth rate was directly co-related to bacteriocin synthesis in these strains. Maximum production of Lactocin RN 78 was achieved in phases when the growth was at its peak. Contrasting results has been reported in salivacin 140 where the synthesis was not related to its growth (Arihara et al., 1996). However a sharp decline in the growth rate and antibacterial activity was observed after 48 h of incubation which could be due to the exhaustion of some essential micronutrients (vitamins and minerals) or amino acids in the media.
Optimal temperature for the growth of L. casei RN 78 was 35°C while minimum growth and synthesis was recorded at temperatures above 42°C and below 4°C. These results are in contrast with findings of Larsen et al. (1993) where maximum growth and production of bavaricin A was reported during the growth of L. bavaricus M1401 at temperature as low as 4°C.
The effect of pH on the bacteriocin production of a number of bacteria is well documented. According to Parente and Hill (1992), pH has the greatest influence on bacteriocin production and the more the synthesis being pH tolerant the better are the chances for its selection in the food system. Here we observed the growth and production of L. casei RN 78 at a wide pH range of 4.0 to 9.0. This property of lactocin RN 78 appears similar to pediococcin AcH and lactacin B produced by Pediococcus acidilactiae and Lactococcuc lactis B (Barefoot and Klaenhammer, 1983; Joerger and Klaenhammer 1986), which have reported the wide pH stability of the bacteriocins in study. An interesting observation was seen at alkaline pH values exceeding 8.0, where growth and production in L.casei RN 78 was evident after prolong incubation period. This delay in growth and activity could be possibly due to slow acidification of medium during the initial h of growth and acid produced by the test organism. Once the optimum pH was reached the growth and production accelerated. Similar phenomenon was also recorded for nisin and pediocin production, which were effected by acidification rate (Cabo et al., 2001; Guerra and Pastrana, 2002). An increase in acidity of the hydrolyzed medium enhanced the synthesis of both nisin and pediocin by L. lactis and P. acidilactiae, respectively.
When the effect of the salt was premeditated on the growth and bacteriocin production of this potentially important strain, a salt tolerance of 3% was exhibited. However it was unable to resist salt concentrations higher than 3% and the growth was completely inhibited at these concentrations. As reported earlier (Thomas et al., 1996; Rozes and Peres, 1996; Verluyten et al., 2004), high concentrations of salt might be responsible for uncoupling of growth and production or due to its action on the intracellular enzymes leading to cell death.
It was interesting to record production of Lactocin RN 78 in the presence of different concentrations of bile salts. The ability of the test organism to grow and synthesize in the presence of 0.3% bile salt indicates its potential application in probiotic formula especially in chickens against pathogen infection by listeria strains.
Previously, a number of authors have concluded that complex media are essential for bacteriocin production (Geis et al., 1983; Parente and Hill, 1992). The results presented by these authors were observed when L. casei RN 78 was grown in different growth media. A good correlation between the growth metabolism and bacteriocin production of the test strain especially in a media supplemented with lactose was recorded. Such findings have also been reported for other bacteriocins like pediocin AcH and salivaricin CRL 1384 production where the authors concluded that glucose and lactose are the best carbon source in the media for the maximum production of these bacteriocins (Parente and Hill, 1992; Audisio et al., 1999, 2001; Audisio and Apella, 2006).
The stability of bacteriocins under various stress conditions indicated the proteinaceous substance tolerant of variable stress factors. All these factors indicate the possible exploitation of this strain in different food and probiotic products. Nevertheless, before final assessments it is essential to evaluate the stability of this bacteriocin in in situ conditions in order to further refine these criteria and get deeper knowledge of the mechanism of their action. All these factors might help us to achieve improved and highly desirable biological preservatives and active probiotic product in near future.
REFERENCES
- Arihara, K., S. Ogihara, T. Mukai, M. Itoh and Y. Kondo, 1996. Salivacin 140, a novel bacteriocin from Lactobacillus salivarius subsp. salicinius T140 active against pathogenic bacteria. Lett. Applied Microbiol., 22: 420-424.
CrossRefDirect Link - Carina Audisio, M. and M.C. Apella, 2006. Bacteriocin-like substance produced by Lactobacillus salivarius subsp. salivarius CRL1384 with anti-Listeria and anti-Salmonella effects. Res. J. Microbiol., 1: 61-69.
CrossRefDirect Link - Avonts, L., E. Van Uytven and L. De Vuyst, 2004. Cell growth and bacteriocin production of probiotic Lactobacillus strains in different media. Int. Dairy J., 14: 947-955.
Direct Link - Barefoot, S.F. and T.R. Klaenhammer, 1983. Detection and activity of lactacin B, a bacteriocin produced by Lactobacillus acidophilus. Applied Environ. Microbiol., 45: 1808-1815.
Direct Link - Bhunia, A.K., M.C. Johnson, B. Ray and N. Kalchayanand, 1991. Mode of action of pediocin AcH from Pediococcus acidilactis H on sensitive bacterial strains. J. Applied Bacteriol., 70: 25-33.
Direct Link - Brink, B.T., M. Minekus, J.M.B.M. van der Vossen, R.J. Leer and J.H.J.H. Intveld, 1994. Antimicrobial activity of lactobacilli: Preliminary characterization and optimization of production of acidocin B, a novel bacteriocin produced by Lactobacillus acidophilus M46. J. Applied Bacteriol., 77: 140-148.
CrossRefPubMedDirect Link - Cabo, M.L., M.A. Murado, M.P. Gonzalez and L. Pastoriza, 2001. Effects of aeration and pH gradient on nisin production. A mathematical model. Enzyme Microbiol. Technol., 29: 264-273.
Direct Link - Casla, D., T. Requena and R. Gomez, 1996. Antimicrobial activity of lactic acid bacteria isolated from goat's milk and artisanal cheeses: Characteristics of a bacteriocin produced by Lactobacillus curvatus IFPL105. J. Applied Bacteriol., 81: 35-41.
CrossRefDirect Link - Cleveland, J., T.J. Montville, I.F. Nes and M.L. Chikindas, 2001. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol., 71: 1-20.
CrossRefPubMedDirect Link - Coventry, M.J., J.B. Gordon, M. Alexander, M.W. Hickey and J. Wan, 1996. A food-grade process for isolation and partial purification of bacteriocins of lactic acid bacteria that uses diatomite calcium silicate. Applied Environ. Microbiol., 62: 1764-1769.
Direct Link - Delves-Brougthon, J., 1990. Nisin and its uses as a food preservative. Food Technol., 44: 100-112.
Direct Link - Delves-Broughton, J., P. Blackburn, R.J. Evans and J. Hugenholtz, 1996. Applications of the bacteriocin, nisin. Antonie van Leeuwenhock, 69: 193-202.
CrossRefDirect Link - De Vuyst, L. and E.J. Vandamme, 1994. Nisin, a Lantibiotic Produced by Lactococcus lactis Subsp. lactis: Properties, Biosynthesis, Fermentation and Applications. In: Bacteriocins of Lactic Acid Bacteria: Microbiology, Genetics and Applications, De Vuyst, L. and E.J. Vandamme (Eds.). Blackie Academic and Professional, London, England, pp: 151-221.
- Geis, A., J. Singh and M. Tueber, 1983. Potential of lactic streptococci to produce bacteriocin. Appl. Envir. Microbiol., 45: 205-211.
Direct Link - Guerra, N.P. and L. Pastrana, 2002. Production of bacteriocin from Lactococcus lactis subsp lactis CECT 539 and Pediococcus acidilactici NRRL B-5627 using mussel-processing wastes. Biotechnol. Applied Biochem., 36: 119-125.
Direct Link - Harris, L.J., M.A. Daeschel, M.E. Stiles and T.R. Klaenhammer, 1989. Antimicrobial activity of lactic acid bacteria against Listeria monocytogenes. J. Food Protect., 52: 384-387.
Direct Link - Howell, T.H., J.P. Fiorellini, P. Blackburn S.J. Projan, J. De La Harpe and R.C. Williams, 1993. The effect of a mouthrinse based on nisin, a bacteriocin, on developing plague and gingivitis in beagle dogs. J. Clin. Periodont., 20: 335-339.
PubMedDirect Link - Jack, R.W., J.R. Tagg and B. Ray, 1995. Bacteriocins of gram-positive bacteria. Microbiol. Rev., 59: 171-200.
Direct Link - Joerger, M.C. and T.R. Klaenhammer, 1986. Characterization and purification of helveticin J and evidence for a chromosomally determined bacteriocin produced by Lactobacillus helveticus 481. J. Bacteriol., 167: 439-446.
Direct Link - Klaenhammer, T.R., 1993. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev., 12: 39-85.
CrossRefPubMedDirect Link - Lewus, C.B. and T.J. Montiville, 1991. Detection of bacteriocins produced by lactic acid bacteria. J. Microbiol. Methods, 13: 145-150.
CrossRef - Lucke, F.K., 2000. Utilization of microbes to process and preserve meat. Meat Sci., 56: 105-115.
CrossRefDirect Link - Meaney, W.J., D.P. Twomey, J. Flynn, C. Hill and R.P. Ross, 2001. The use of a bismuth-based teat seal and the bacteriocin Lacticin 3147 to prevent dry period mastitis in dairy cows. Proceedings of the British Mastitis Conference, Garstang, (BM'01), Institute for Animal Health/Milk Development Council, pp: 24-32.
- Minahk, C.J., F. Dupuy and R.D. Morero, 2004. Enhancement of antibiotic activity by sub-lethal concentrations of enterocin CRL 35. J. Antimicrob. Ther., 53: 240-246.
Direct Link - Muriana, P.M. and T.R. Klaenhammer, 1991. Cloning, phenotypic expression and DNA sequence of the gene for lactacin F, a bacteriocin produced by L. acidophilus. J. Bacteriol., 173: 1779-1788.
Direct Link - Nam, H., M. Ha, O. Bae and Y. Lee, 2002. Effect of Weissella confusa strain PL9001 on the adherence and growth of helicobacter pylori. Applied Environ. Microbiol., 68: 4642-4645.
Direct Link - Piard, J.C. and M. Desmazeaud, 1991. Inhibiting factors produced by lactic acid bacteria part 1. Oxygen metabolites and catabolism end-products. Lait, 71: 525-541.
CrossRef - Piard, J.C. and M. Desmazeaud, 1992. Inhibiting factors produced by lactic acid bacteria. 2. Bacteriocins and other antibacterial substances. Lait, 72: 113-142.
CrossRefDirect Link - Rekhif, N., A. Atrih and G. Lefebvre, 1994. Characterization and partial purification of plantaricin LC 74, a bacteriocin produced by L. plantarum LC 74. Biotechnol. Lett., 16: 771-776.
Direct Link - Rozes, N. and C. Peres, 1996. Effect of oleuropein and sodium chloride on viability and metabolism of Lactobacillus plantarum. Applied Microbiol. Biotechnol., 45: 839-843.
Direct Link - Severina, E., A. Severin and A. Tomasz, 1998. Antibacterial efficacy of nisin against multidrug-resistant Gram-positive pathogens. J. Antimicrob. Chemother., 41: 341-347.
Direct Link - Tagg, J.R., A.S. Dajani and L.W. Wannamaker, 1976. Bacteriocins of gram-positive bacteria. Bacteriol. Rev., 40: 722-756.
CrossRefPubMedDirect Link - Thomas, L.V. and J.W.T. Wimpenny, 1996. Investigation of the effect of combined variations in temperature, pH and NaCl concentrations on nisin inhibition of Listeria monocytogenes and Staphylococcus aureus. Applied Environ. Microbiol., 62: 2006-2012.
Direct Link - Verluyten, J., W. Messens and L. De Vuyst, 2004. Sodium chloride reduces production of curvacin A, a bacteriocin produced by Lactobacillus curvatus strain LTH 1174, originating from fermented sausage. Applied Environ. Microbiol., 70: 2271-2278.
Direct Link - Yang, R., M.C. Johnson and B. Ray, 1992. Novel method to extract large amounts of bacteriocin from lactic acid bacteria. Appl. Environ. Micrbiol., 58: 3355-3359.
CrossRefDirect Link - Itoh, T., Y. Fujimoto, Y. Kawai, T. Toba and T. Saito, 1995. Inhibition of food-borne pathogenic bacteria by bacteriocins from Lactobacillus gasseri. Lett. Applied Microbiol., 21: 137-141.
CrossRefDirect Link