
R. Subashkumar
Department of Environmental Sciences, Bharathiar University, Coimbatore-641 (146, India
Tha. Thayumanavan
Department of Environmental Sciences, Bharathiar University, Coimbatore-641 (146, India
C. Thilagavathy G. Vivekanandhan
Department of Environmental Sciences, Bharathiar University, Coimbatore-641 (146, India
K. Savithamani
Department of Microbiology, KSR College of Alts and Science, Tiruchengode, 690 204, India
P. Lakshmanaperumalsamy
Department of Biotechnology, RVS College of Alts and Science,
Coimbatore-641 402, India
International Journal of Dairy Science
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 70-83







ABSTRACT
A total of 105 milk samples were analysed for Aeromonas hydrophila and 17.14% of the milk samples were found to be contaminated with A. hydrophila. Out of the milk samples, 30.5% were found to be of poor quality. The A. hydrophila strains were resistant to bacitracin, cefazolin and novobiocin and sensitive to gentamycin, kanamycin, neomycin and polymyxin-B. About 94.4% of strains were found to be haemolysin producers. Proteolytic activity was exhibited by all the isolates. The environmental factors such as NaCl and pH were influencing the growth of the A. hydrophila isolates. None of the RAPD and ERIC profiles were found identical. This clearly revealed the wide genetic heterogeneity among the strains tested.
PDF Abstract XML References
How to cite this article
R. Subashkumar, Tha. Thayumanavan, C. Thilagavathy G. Vivekanandhan, K. Savithamani and P. Lakshmanaperumalsamy, 2006. Typing of Haemolytic and Antibiotic Resistant Aeromonas hydrophila Isolated from Raw Milk of Coimbatore, South India. International Journal of Dairy Science, 1: 70-83.
DOI: 10.3923/ijds.2006.70.83
URL: https://scialert.net/abstract/?doi=ijds.2006.70.83
DOI: 10.3923/ijds.2006.70.83
URL: https://scialert.net/abstract/?doi=ijds.2006.70.83
INTRODUCTION
Raw milk could be one of the sources of food-borne pathogens. The consumption of raw milk may result in sporadic disease outbreaks. Pasteurization is designed to destroy all bacterial pathogens common to raw milk, excluding spore-forming bacteria and possibly Mycobacterium paratuberculosis, but still some people are fond of drinking raw milk, believing it as safe. Current methods for assessing the bacteriological quality of raw milk, such as aerobic plate counts, are not usually designed to detect specific pathogens, such as the species of Mycobacterium, Escherichia, Clostridium, Pseudomonas, Bacillus, Aeromonas, Staphylococcus, Streptococcus, Yersinia, Listeria, Campylobacter which are responsible for gasteroenteritis, septicaemia and other food borne infection. A. hydrophila have been frequently isolated from milk and dairy products (Keilwein, 1971) in variable frequencies. In recent years, A. hydrophila has been increasingly recorded from cases of diarrhoea and other infections (Ananthan and Alavandi, 1999; Vila et al., 2003). It can produce a number of potential virulence factors including cytotoxin, haemolysin and cytotoxic enterotoxin (Krovacek et al., 1991) and has been received increased attention as an emerging human pathogen. Moreover, many of these strains are able to grow and produce virulence in refrigerated temperature (Kirov, 1997). This raises the possibility of high number of organisms in foods. Under these, Aeromonas strains harboring one more virulence genes have been shown to produce toxins. Still, little is known about the impact of pathogenic A. hydrophila on food related illness.
Since Aeromonas has gained importance in recent years as an etiological agent and the presence of Aeromonas in milk was not investigated in this region, the present study was carried out to find the load of incidence of A. hydrophila in milk. Selected isolates were also typed by RAPD and ERIC PCRs to determine the genetic homogeneity among the strains.
MATERIALS AND METHODS
Raw milk samples were collected from various milk vendors of Coimbatore, south India during June-November 2003 (Table 1). The collected samples were aseptically processed with in two hours. The quality of the milk was detected by MBRT test.
Isolation and Identification of A. hydrophila
The milk samples were enriched in Alkaline Peptone Water (APW) as recommended by Shread et al. (1981). A loopful of enriched culture was streaked on SAA and Rimler Shotts agar medium (Palumbo, 1985). Yellow to honey coloured, oxidase, catalase, glucose, fermentation positive colonies were taken and other biochemical identifications were performed (Kaper, 1981; Buchanan and Gibbons, 1984; Palumbo, 1985) and followed by the final confirmation of A. hydrophila using Aero II (48) key. Two control strains of A. hydrophila (MTCC 646 and ATCC 14715) were used in parallel.
Determination of Multiple Antibiotic Resistance
Antimicrobial resistance was performed by using agar disk diffusion method (Bauer et al., 1966). A. hydrophila strains were tested against commercially available antibiotic discs (HiMedia, India) (Table 2).
| Table 1: | Percentage of incidence of A. hydrophila |
 | |
| Table 2: | Percentage of antibiotic resistant A. hydrophila (n = 18) isolated from milk |
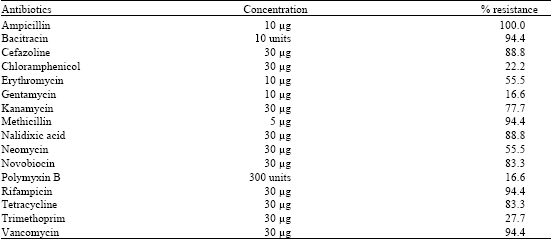 | |
The Multiple Antibiotic Resistance (MAR) index, when applied to a single isolate, is defined as a/b, where a represents the number of antibiotics to which the isolate was resistant and b represents the number of antibiotics to which the isolates was exposed. MAR index higher than 0.2 identifies organisms originated from high-risk sources of contamination, where antibiotics are often used. MAR indices less than, or equal to 0.2 identify strains from environments where antibiotics are seldom or never used (Krumperman, 1985).
Haemolysin Assay
The haemolytic activity of A. hydrophila was determined by both blood agar plate and microtitre plate assay.
Blood Agar Plate Assay
Haemolytic activity was determined as a zone of haemolysis around the colonies on blood agar plates containing 5% (v/v) human blood, after 24 h incubation at 37°C (Brenden and Janda, 1987).
Microtitre Plate Assay
A. hydrophila strains were cultured in 5 mL of BHIB (HiMedia, India) and incubated at 37°C for 16-18 h. Supernatant fluids were carefully collected after centrifugation at 10,000 X g for 30 min at 4°C. One hundred micro liters of supernatant fluids were mixed with equal volume of 2% (v/v) suspension of human erythrocytes in a 96 well “V”-bottom micro titre tray. The mixture was incubated for 1 h at 37°C and then for 1 h at 4°C. Erythrocyte suspension in PBS was included in each as a negative control. Haemolysin production was recorded by visual inspection (Burke et al., 1983). Each culture filtrate was tested in duplicate.
Proteolytic Activity
The proteolytic activity was determined by modified method of Charney and Tomerelli (1947). The test culture was streaked on to nutrient agar plates and after incubation the cells were harvested using sterile physiological saline (0.85% NaCl in distilled water). Then it was centrifuged at 12,000 rpm for 30 min and pellet was suspended in physiological saline. The OD of the suspension was adjusted to 1 OD. One milliliter of the cell suspension was added to the nutrient broth (100 mL) and incubated in a shaker at 30°C. The cell suspension was centrifuged at 15,000 rpm for 30 min and the cell free supernatant fluid was taken as crude enzyme. One mL of crude enzyme was mixed with substrate (Casein according to hammerstein, 7.0 g (HiMedia, India) in 1.2% Tris (hydroxy methyl) aminomethane (HiMedia, India), pH 7.6 and 8 mL of 1N HCl) and was kept in a water bath at 37°C for 30 min. The reaction was stopped by adding 10 mL of trichloroacetic acid and kept for 37°C for 30 min. It was filtered through Whatman No. 24 filter paper. The absorbance was determined of each sample filtrate at 275 nm (Hitachi-U 3210, Japan). The test was run in duplicate and the average was defined as the enzyme activity, which liberated 1 μg of Tyrosine mL-1 of the reaction mixture/minute under standard conditions. Proteolytic units of activity (PU mL-1) were recorded as changes in absorbance at 275 nm mL-1 cell free supernatant min-1 at 37°C (Mc Mahon, 2000).
Effect of NaCl and pH on Growth
A. hydrophila cells were grown in BHIB (HiMedia, India) medium for 18 h at 37°C. The cells were concentrated by centrifugation at 3000 rpm for 15 min and washed three times with 0.85% of sterile saline. After the final wash, 1 mL of 1 OD concentration cells were inoculated into different concentrations (0.3, 0.5, 0.7, 1.0, 1.5 and 2.0%) of NaCl solutions and incubated at 37°C for 24 h. Another 1 mL of 1 OD concentration of A. hydrophila strains were inoculated onto the BHIB, containing different range of pH such as 4, 5, 6, 7, 8 and 9 and incubated at 37°C for 24 h. After incubation the growth was measured in a spectrophotometer (Hitachi-U 3210, Japan) at 670 nm.
Molecular Typing
DNA Extraction
Four strains were randomly selected and grown overnight at 37°C on a LB broth. Reference strain such as MTCC 646 and ATCC 14715 culture were used in parallel. Total DNA was purified from overnight culture by using GENTRA DNA extraction kit (Gentra System, Inc., USA), according to the manufacturer’s instructions. The extracted DNA was diluted into 20 ng μL-1 and used as working concentration for Randomly Amplified Polymorphic DNA (RAPD) PCR and Entrobacterial Repetitive Intergenic Consensus Sequence (ERIC) PCR tests.
RAPD-PCR
One hundred ng of each DNA was individually used and a final volume of 50 μL was used for PCR amplification. RAPD mixtures were prepared as previously described by Oakey et al. (1996). A typical RAPD PCR reaction mix contained 5 μL of 10X PCR buffer II (100 mM Tris-HCl. pH 8.3; 500 mM KCL), 1 μL of dNTPs (10 mM), 7 μL of primer, 0.4 μL of Taq DNA polymerase, 1 μL (100 ng μL-1) of template DNA and 35.6 μL of DNA quality water to make a final volume up to 50 μL DNA. Amplification was performed using thermal cycler (MJ Research, Model PTC 100 Watertown, Mass., USA). The polymerase chain reaction DNA amplification protocol was a modification of that of proposed by Delamare et al. (2002). The amplification conditions were as follows: 35 cycles of denaturation at 94°C for 10 sec, annealing at 37°C for 30 sec and extension at 72°C for 1 min. A negative control including all components except genomic DNA was included in all thermal cycle runs. The primer used in this study was 5’-TCG CGA GCT G-3’.
ERIC-PCR
The ERIC - PCR method utilizes primers complementary to ERIC sequences of bacterial genomic DNA (Versalovic, 1991). Following primers were used for ERIC 1 (5’-ATG TAA GCT CCT GGG GAT TCA C-3’) and ERIC 2 (5’-AAG TAA GTG ACT GGG GTG AGC G-3’). One hundred ng of DNA sample was used and final volume of 50 μL was used for PCR amplification. A typical ERIC-PCR reaction mix contained 2.5 μL of 10X PCR buffer II (100 mM Tris-HCl, pH 8.3; 500 mM KCl), 0.2 μL of dNTPs (10 mM), 1.25 μL of primer (ERIC I), 1.25 μL of primer (ERIC II), 0.2 μL of Taq DNA polymerase, 1 μL (100 ng) of template DNA and 43.6 μL of DNA quality water to make a final volume up to 50 μL. DNA amplification was performed using thermal cycler (MJ Research, Model PTC 100 Watertown, Mass., USA). Slight modification was made in the ERIC-PCR cycle, which was described by Szczuka and Kaznowski (2004). The reaction mixtures were denaturated at 94°C for 3 min and then subjected to 35 cycles of denaturation at 94°C for 30 sec, annealing at 45°C for 1 min and extension at 72°C for 5 min and final extension at 72°C for 5 min.
Amplified products were resolved by electrophoresis in 1.5% (w v-1) agarose gels containing ethidium bromide (1.6 mg mL-1) at 11 V cm-1 for 90 min in TBE buffer (0.089 M Tris, 0.089 M boric acid, 0.002 M EDTA (pH 8)). Sizes of the amplified products were determined by comparison with 2 Log DNA ladder (New England Biolabs, UK). All PCR and bacterial enumeration were conducted in duplicate.
RESULTS
Out of 105 samples, 17.14% of them were found to be contaminated with A. hydrophila. None of the milk samples were positive for A. hydrophila during June and July. About 21.05% of A. hydrophila strains were contaminated during August 2003. During November 2003, 40% were positive for A. hydrophila (Table 1). Among 105 milk samples, 30.48% of samples were of poor quality (data not shown). Their methylene blue reduction time was recorded as 15 min.
| Table 3: | MAR index and antibiotic resistance patterns of A. hydrophila isolated from milk |
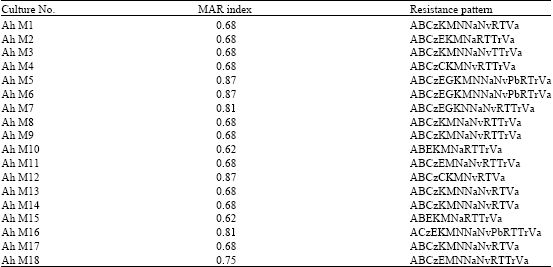 | |
| Table 4: | Haemolytic and proteolytic activity of A. hydrophila isolated from milk |
 | |
All the 18 isolates were subjected to antibiotic resistance tests (Table 2). Predominantly, the strains were resistant towards ampicillin (100%), bacitracin (94.4%), methicillin (94.4%), rifampicin (94.4%), vancomycin (94.4%), cefazolin (88.8%) and nalidixic acid (88.8%). Five groups in MAR index were recorded. About 66.6% of the strains showed a MAR index value of 0.6 and 27.7% of the strains of 0.8. The results exhibited that the indices were higher than 0.2 (Table 3).
About 94.4% of the isolates were haemolysin producers at 37°C. The isolates varied in their ability to lyse the RBC’s of human origin. Overall 72, 22 and 6% of isolates were beta, alpha and gamma haemolysis, respectively (Table 4). The protease activity of all the A. hydrophila were tested in neutral pH condition at 37°C (Table 4). The protease activity was more in Ah M 7 (146.02 PU mL-1). All other isolates showed moderate protease activity. The influence of various concentrations of NaCl on the growth of all the eighteen strains of A. hydrophila showed that the maximum growth was in 1.5% NaCl. The growth of the organisms were imperfect at 0.3 to 1.0% NaCl. Ah M15 strain has shown growth at 2% NaCl and Ah M7 at 3% NaCl concentration (Fig. 1).
A. hydrophila strains grew well at pH 4-9. At pH 6 and 8, most of the strains showed moderate growth. Optimum growth was found at pH 7 for the 61.1% of isolates. Ah M9 and Ah M14 have shown maximum growth at pH 9 (Fig. 2).
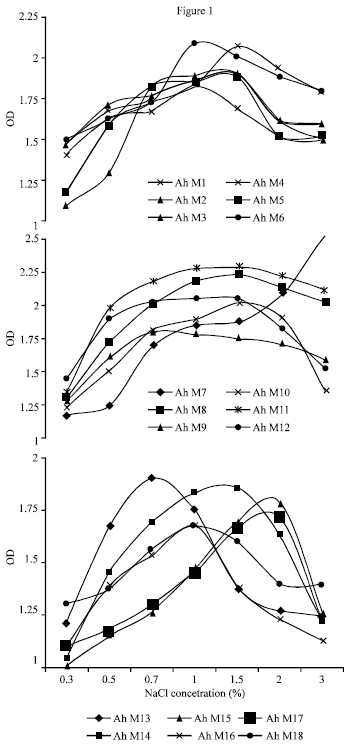 | |
| Fig. 1: | Influence of NaCl concentration on the growth of A. hydrophila isolated from milk |
The RAPD and ERIC profiles of A. hydrophila strains are given in Fig. 3 and 4, respectively. Lane 1 and 2 represents reference strains MTCC 646 (Ah T1) and ATCC 14715 (Ah T2). In RAPD profiles, the type strains produced 7 and 5 bands, the size ranging from 0.85-3.0 kb.
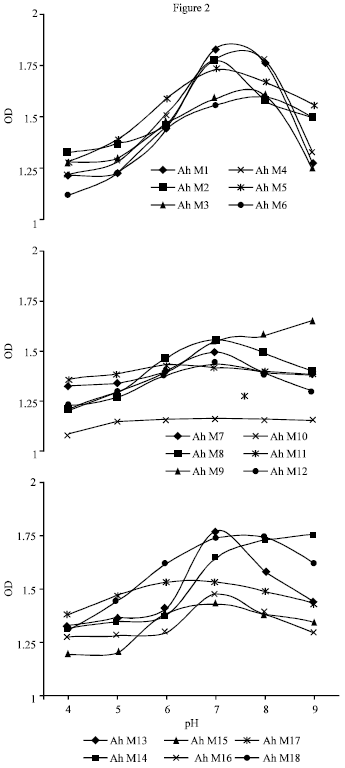 | |
| Fig. 2: | Influence of pH on the growth of A. hydrophila isolated from milk |
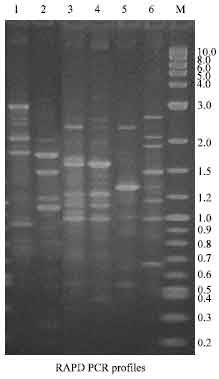 | |
| Fig. 3: | RAPD profiles of A. hydrophila. Lane 1: Ah T1 (MTCC 646); Lane 2: Ah T2 (ATCC 14715); Lane 3: Ah M1 (Milk); Lane 4: Ah M2 (Milk); Lane 5: Ah M3 (Milk); Lane 6: Ah M4 (Milk); Lane M: 2 log DNA ladder (0.1-10 kb); |
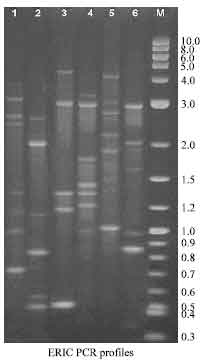 | |
| Fig. 4: | ERIC PCR profiles of A. hydrophila. Lane 1: Ah T1 (MTCC 646); Lane 2: Ah T2 (ATCC 14715); Lane 3: Ah M1 (Milk); Lane 4: Ah M2 (Milk); Lane 5: Ah M3 (Milk); Lane 6: Ah M4 (Milk); Lane M: 2 log DNA ladder (0.1-10 kb) |
The fingerprint of the milk isolates (Ah M1 to Ah M4) consisted of 8 to 4 amplification bands. The Ah M1 showed eight bands with molecular size of 2.4, 1.9, 1.7, 1.6, 1.2, 1.1 and 1.0 kb. In Ah M2, DNA profile consisting the fragments of 2.4, 1.6, 1.2, 1.1 and 1.0 kb. Ah M3 appeared to be identical and shared the following common bands: 2.4, 1.3 and 1.0 kb. The molecular size of 2.7, 2.1, 1.9, 1.5, 1.1, 1.0 and 0.6 kb were obtained in the sixth lane (Ah M4). Ah M1 and Ah M2 represent four distinct common bands, from that three bands were similar to the ATCC strain (Ah T2). In this study, the pattern of the amplified PCR product showed only one common band with molecular weight in 1.0 kb in all the strains tested except MTCC type strains. None of the ERIC profile was found identical. Also no common bands were found among the profiles (Fig. 4). However, Ah M1 and Ah M2 showed two common amplified DNA products such as 3.0 and 1.3 kb. The Ah M3 and Ah T2 type strain have two similar amplified DNA in the position of 2.0 and 0.85 kb. The strains used in this study showed major diverse bands.
DISCUSSION
Milk is an excellent culture medium for many kinds of microorganisms, being high moisture, nearly natural pH and rich in microbial nutrients. Several types of bacteria that grow and become significant will depend on the initial microflora of the milk (Barmley and McKinnon, 1990). The incidence and properties of Aeromonas species in milk is the vehicle for the transmission of Aeromonas gastroenteritis, because of the subsequent handling of the milk. Some of these strains were indistinguishable from diarrhea-associated strains and were able to produce exotoxins at 37°C and adhere to epithelial cells. A total of 105 raw milk samples were subjected to MBRT. About 30.48% of milk reduction time was recorded at 15 min, it was expressed to confirm the poor quality.
The presence of A. hydrophila in the food chain should not be ignored. More information is needed to evaluate its public health significance as a potential cause of food poisoning (Buchanan and Palumbo, 1985). In this study, 17.14% of the milk samples were contaminated with A. hydrophila. Melas et al. (1999) reported that a total of 138 raw cow’s milk samples were contaminated with A. hydrophila.
The antibiotic resistance pattern of the A. hydrophila exhibited multiple antibiotic resistances; however resistance was shown against cefazoline, rifampicin, methicillin and vancomycin. The previous study of Ramteke et al. (1994) showed the occurrence of chloromphenicol resistant strains in India and this was similar to present study. Chaudhury et al. (1996) reported a growing incidence of multiple antibiotic resistant Aeromonas species from clinical and environmental sources. World wide, there is growing concern about increased prevalence of antibiotic resistance and it is now generally accepted that the main risk factor is increased use of antibiotics (Levy, 1992; Radu et al., 2003). Such high level of multiple resistances may arise from selective pressure due to indiscriminate use of antibiotics. The frequencies of bacterial strains resistant to antimicrobial agents increased dramatically in the environment as a consequence of these drugs (Levy, 1992; Young, 1993). A transfer of plasmid-encoded resistance to antimicrobial agent is a significant public health concern; the possibility of antibiotic resistance between A. hydrophila in milk has attracted attention.
The A. hydrophila strains showed multiple antibiotic resistance, which was similar to earlier reports (De Vincente et al., 1990; Davies, 1992; Vivekanandhan et al., 2002; Radu et al., 2003). MAR index value of 0.2 or above is said to be the strain from high-risk source contamination (Krumperman, 1985; Vivekanandhan et al., 2002). The strains isolated from milk exhibited their MAR index value of more than 0.2 recorded. The release of MAR organisms may have originated from the human sources and may ultimately pave way for the cross contamination of food, water and other matter (Krumperman, 1985).
About 94.4% were positive for haemolysin activities. The isolates varied in their ability to lyse the erythrocytes of human origin. Attention has been given on the haemolysin of A. hydrophila, because the production of haemolytic toxin has been regarded as indication of pathogenic potential, though non-haemolytic aeromonads have also been implicated as human pathogen (Namdari and Bottone, 1990). As defined by Wong et al. (1996), all the Aeromonas isolates with haemolysin positive genotype were virulent in the suckling mouse model assay. The involvement of Aeromonas species in food poisoning is still controversial and the number of potential enterotoxins that have suggested over the years may show that the pathogenesis of Aeromonas infection is complex and multifactorial (Granum et al., 1998; Chopra and Houston, 1999). Granum et al. (1998) reported haemolytic positive strains from tap water, dairy products and raw fish and it is very interesting to note that all A. hydrophila strains were positive for haemolysin production. Handfield et al. (1996) reported that haemolysin producing strains were more frequently encountered in dairy products. Apparently, Aeromonas strains, which were able to infect the human gasterointestinal tract, were rarely found in drinking water and food (Handfield et al., 1996). Alternatively, the haemolysin may be released in an inactive form into culture supernatants and then be activated later by protease produced by the cells; this could explain the inhibition of this biological activity (Nomura et al., 2000).
Proteolytic activity was found in the culture supernatant but not in the cells and the increase in activity did not directly parallel cell growth (O’Rielly and Day, 1983). In our study, the culture supernatant (crude enzyme) digested the protein substrate at 30°C by 30 min. Castro-Escarpulli et al. (2003) reported that 61% of the Aeromonas isolates produced protease. The role of protease in disease induction and lesion formation may provide the basis for Aeromonas disease control. It strongly suggested the involvement of protease in pathogenecity and the regulation of protease production by A. hydrophila. However, it has not been thoroughly investigated (O’Rielly and Day, 1983). The protease cleaves the prohaemolysin (activated aerolysin-protoxin) resulting in the removal of a 42 aminoacid peptides, which converts the prohaemolysin into haemolysin (Nomura et al., 1999). The haemolysin and protease production were found more frequently in the clinical samples, which may be important in colonization through the disruption of the intestinal barrier (Sechi et al., 2002).
The environmental factors influence the growth of A. hydrophila. The NaCl concentration of 0.7 and 1% favoured the growth of 22.2% of the isolates. About 44.4% of the isolates grew well at 1.5% NaCl. The similar findings were reported by Vivekanandhan et al. (2003) and stated that increase in the salt concentration resulted in the decrease in growth. Palumbo et al. (1985) reported that most of the clinical isolates could grow at 4% NaCl at 28°C, while our Ah M7 survive at 3% NaCl but did not multiply. However, Popoff (1984) indicated that motile Aeromonas do not grow at 5% NaCl, but in a previous study, Popoff and Veron (1976) reported that one strain grew at 10% NaCl. The experiment on the influence of pH on A. hydrophila revealed that moderate growth occurred in the pH 7 and 8. The similar findings were reported by Vivekanandhan et al. (2003). The population level increased while increasing the alkalinity. However viable cells were present in acidic pH also. Similar trend of growth was reported by Popoff and Lallier (1984).
Determining the RAPD and ERIC PCR profiles of A. hydrophila has been inconclusive, probably because of the differences in diarrhoeagenic potential among strains (Borchardt et al., 2003). In recent reports, Davin-Regli et al. (1998) and Aguilera-Arreola et al. (2005) proved that both RAPD and ERIC-PCRs are powerful tools for the differentiating the strains of A. hydrophila, while Szczuka and Kaznowski (2004) reported that both the RAPD and ERIC-PCRs have the same discriminatory power for the species of Aeromonas. They have also stated that Repetitive Extragenic Polindromic (REP) PCR is less effective for differentiating Aeromonas isolates.
A great heterogeneity within the strains of A. hydrophila used in these studies was demonstrated by RAPD and ERIC-PCRs. Davin-Regli et al. (1998) found 6 different patterns among 10 clinical isolates of A. hydrophila tested. They also found that all the 13 water isolates of A. hydrophila used as controls gave different profiles. They concluded that no fingerprints were common to both environmental and clinical isolates of A. hydrophila. Moyer et al. (1992) also found no genetic relationship between clinical and environmental strains of A. hydrophila. The RAPD profiles showed few similarities between the Ah M1 and Ah M2. The amplified DNA of 1.0 kb occur in all the milk strains (Ah M1to Ah M4) and ATCC strains. But in ERIC-PCR profile, no such similarity was observed. Kuhn et al. (1992) have reported that a single strain could be found in several sampling sites and at different times. The ERIC profile did not show any such similarity between the strains used in the study.
This clearly revealed the wide genetic diversity in the A. hydrophila isolates from milk and type strains tested i.e., the co-existence of multiple A. hydrophila strains among various sources. In the present investigation, RAPD or ERIC-PCRs demonstrated no relationship between milk and typed strains of A. hydrophila. Moderate similarity occurred in RAPD fingerprintings, but not in the ERIC fingerprinting. Variation in the growth of A. hydrophila stains due to the environmental factors could be possible because of its habitat under natural and artificial stress. The results suggest that pathogenic Aeromonas strain could be of public health significance in milk and milk products that have an extended shelf-life at all conditions. Currently, adequate awareness is essential for preventing the spread of food borne pathogens.
Moreover, it has been shown that the level of contamination at the moment of consumption depends up on the initial contamination and the opportunity for the growth and/or survival during processing, preservation and storage of milk and milk products. Polymorphism found among milk isolates of A. hydrophila is very high and highly heterogeneous. Further research is also needed to establish contamination routes. So far, the significance of A. hydrophila in milk remains undefined, although there is a growing concern about the consumption of Aeromonas contaminated milk by children, the elderly and immunocompromised.
ACKNOWLEDGMENTS
The authors acknowledge the Head, Department of Environmental Sciences, Bharathiar University, Coimbatore-641 046, India for providing necessary facilities. We are also greatful to Dr. Duncan Colquhoun, Senior Research Scientist Section for Fish Health, National Veterinary Institute, Oslo, Norway.
REFERENCES
- Aguilera-Arreola, M.G., C. Hernandez-Rodriguez, G. Zuniga, M.J. Figueras and G. Castro-Escarpulli, 2005. Aeromonas hydrophila clinical and environmental ecotypes as revealed by genetic diversity and virulence genes. FEMS Microbiol. Lett., 242: 231-240.
CrossRefDirect Link - Bauer, A.W., W.M.M. Kirby, J.C. Sherris and M. Turck, 1966. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol., 45: 493-496.
CrossRefPubMedDirect Link - Burke, V., M. Gracey, J. Robinson, D. Peck, D. Beaman and C. Bundell, 1983. The microbiology of childhood gasteroenteritis. Aeromonas species and other infective agents. J. Infect. Dis., 148: 68-74.
Direct Link - Castro-Escarpulli, G., M.J. Figueras, G. Aguilera-Arreola, L. Soler and E. Fernandez-Rendon et al., 2003. Characterization of Aeromonas sp. isolated from frozen fish intended for human consumption in Mexico. Int. J. Food Microbiol., 84: 41-49.
Direct Link - Charney, J. and R.M. Tomarelli, 1947. A colorimetric method for the determination of the proteolytic activity of duodenal juice. J. Biol. Chem., 171: 501-505.
Direct Link - Chaudhary, A., G. Nath, B.N. Shukla and S.C. Sanyal, 1996. Biochemical characterisation, enteropathogenicity and antimicrobial resistance plasmids of clinical and environmental Aeromonas isolates. J. Med. Microbiol., 44: 434-437.
Direct Link - Chopra, A.K. and C.W. Houston, 1999. Enterotoxins in Aeromonas associated gasteroenteritis. Microb. Infect., 1: 1129-1137.
Direct Link - De Vicente, A., M. Aviles, J.C. Codina, J.J. Borrego and P. Romero, 1990. Resistance to antibiotics and heavy metals of Pseudomonas aeruginosa isolated from natural waters. J. Applied Bacteriol., 68: 625-632.
Direct Link - Handfield, M., P. Simard, M. Couillard and R. Letarte, 1996. Aeromonas hydrophila isolated from food and drinking water: Hemagglutination, hemolysis and cytotoxicity for a human intestinal cell line (HT-29). Applied Environ. Microbiol., 62: 3459-3461.
Direct Link - Krovacek, K., A. Faris and I. Mansson, 1991. Growth and toxin production by Aeromonas hydrophila and Aeromonas sobria at low temperatures. Intl. J. Food Microbiol., 13: 165-176.
Direct Link - Melas, D.E., D.K. Papageorgiou and A.I. Mantis, 1999. Enumeration and confirmation of Aeromonas hydrophila, Aeromonas caviae and Aeromonas sobria isolated from raw milk and other milk products in Northern Greece. J. Food Protec., 62: 463-466.
Direct Link - Nomura, T., H. Hamashima and K. Okamoto, 2000. Carboxy terminal region of haemolysin of Aeromonas sobria triggers dimerization. Microbial Pathogenesis, 28: 25-36.
Direct Link - Nomura, T., Y. Fujii and K. Okamoto, 1999. Secretion of haemolysin of Aeromonas sobria as protein and contribution of the propeptide region removed from the protoxin to the proteolytic stability of the toxin. Microbiol. Immunol., 43: 29-38.
Direct Link - O'Reilly, T. and D.F. Day, 1983. Effects of cultural conditions on protease production by Aeromonas hydrophila. Applied Environ. Microbiol., 45: 1132-1135.
Direct Link - Oakey, H., J.J.T. Ellis and L.F. Gibson, 1996. Differentiation of Aeromonas genomospecies using random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR). J. Applied Bacteriol., 80: 402-410.
Direct Link - Popoff, M. and R. Lallier, 1984. Biochemical and serological characteristics of Aeromonas. Meth. Microbiol., 16: 127-127.
Direct Link - Radu, S., H. Ahmad, F.H. Ling and Abdul Reezal, 2003. Prevalence and resistance to antibiotics for Aeromonas species from retail fish in Malaysia. Int. J. Food Microbiol., 81: 261-266.
CrossRefDirect Link - Szczuka, E. and A.Kaznowski, 2004. Typing of clinical and environmental Aeromonas sp. strains by random amplified polymorphic DNA PCR, repetitive extragenic palindromic PCR and enterobacterial repetitive intergenic consensus sequence PCR. J. Clin. Microbiol., 42: 220-228.
CrossRefDirect Link - Versalovic, J., T. Koeuth and R. Lupski, 1991. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucl. Acids Res., 19: 6823-6831.
CrossRefPubMedDirect Link - Vivekanandhan, G., K. Savithamani, A.A.M. Hatha and P. Lakshmanaperumalsamy, 2002. Antibiotic resistance of Aeromonas hydrophila isolated from fish and prawn of South India. Int. J. Food Microbiol., 76: 165-168.
PubMedDirect Link - Wong, C.Y.F., G. Mayrhofer, M.W. Heuzenroeder, H.M. Atkinson, D.M. Quinn and R.P.L. Flower, 1996. Measurement of virulence of aeromonads using a suckling mouse model of infection. FEMS Immunol. Med. Microbiol., 15: 233-241.
Direct Link - Young, H.K., 1993. Antimicrobial resistance spread in aquatic environments. J. Antimicrob. Chemother., 31: 627-635.
PubMedDirect Link