
Abeer Alqatati
Department of Clinical Laboratory Sciences, Faculty of Science, University of Jordan, Amman, Jordan
Abdel-Sattar S. Hamad Elgazwy
Department of Chemistry, Faculty of Science, Ain Shams University, Abbassia, 11566 Cairo, Egypt
Saeb Aliwaini
Department of Biological Sciences and Biotechnology, Faculty of Sciences, Islamic University of Gaza, Gaza 108, Palestine
LiveDNA: 970.15194
International Journal of Cancer Research
Year: 2020 | Volume: 16 | Issue: 2 | Page No.: 40-47







ABSTRACT
Background and Objective: Palladacycles have been reported to exert significant anticancer activities against different cancer cells. The current study conducted to evaluate the anti-tumor activity of a new palladacycle complex (ASH10) in estrogen receptor-positive (MCF7) and triple-negative (HCC1937) breast cancer cells. Materials and Methods: The effect of ASH10 on cell proliferation was tested by MTT assay. Scratch assay was used to test the anti-migration ability of ASH10. Apoptosis induced by ASH10 was measured by different methods including nuclear staining and detection of apoptosis markers by western blotting. Autophagy induced by ASH10 also measured by LC3II puncta staining and western blotting using the LC3II antibody. Results: The MTT results showed that the ASH10 compound has a strong anti-growth effect. Data showed that ASH10 induces its cytotoxic effect by inducing DNA damage followed by cell cycle arrest, intrinsic apoptosis and autophagy. Compared to untreated cells, ASH10 treated cells showed high levels of DNA damage markers p-H2AX, p-ATM and p53. Importantly, the cell cycle arrest marker p21 was similarly induced by ASH10 treatment. Furthermore, ASH10 induced significant levels of apoptosis as evidenced by nuclear fragmentation and an increase in the levels of PARP cleavage. Interestingly, ASH10 also activated the formation of autophagosomes and increased the autophagy marker LC3II. Inhibition of autophagy led to a decrease in ASH10 cytotoxicity suggesting that ASH10 induced autophagy was a cytotoxic mechanism. Conclusion: These results demonstrated that ASH10 is a potential effective compound in the treatment of both ER-negative and ER-positive breast cancer cells.
PDF Abstract XML References Citation
How to cite this article
Abeer Alqatati, Abdel-Sattar S. Hamad Elgazwy and Saeb Aliwaini, 2020. Newly Synthesized Palladium (II) Complex ASH10 Induces Apoptosis and Autophagy in Breast Cancer Cells. International Journal of Cancer Research, 16: 40-47.
DOI: 10.3923/ijcr.2020.40.47
URL: https://scialert.net/abstract/?doi=ijcr.2020.40.47
DOI: 10.3923/ijcr.2020.40.47
URL: https://scialert.net/abstract/?doi=ijcr.2020.40.47
INTRODUCTION
Breast cancer is the most common cancer among females worldwide1. Despite the huge efforts to cope with this problem, there is still a limited success with most of the current treatments. Transition metals such as platinum have potential therapeutic and diagnostic values. To date, cisplatin and Pt(II) analogs, carboplatin and oxaliplatin remain important chemotherapeutic agents that have been used widely to treat several cancers including breast, ovarian, cervical and non-small-cell lung cancer2. However, due to the severe side effects and multi-drug resistance associated with these treatments, there has been an ongoing search for safer and more effective metallic compounds. Recently, palladacycles have been reported as potential anti-cancer compounds against a wide range of cancers including melanoma and breast cancer3-5. Importantly, while both platinum and palladium compounds induce DNA damage and cell death, the palladium compounds have a superior degree of cytotoxicity in different cancer cells6. Several palladium-based compounds have been shown to induce their cytotoxic effects via a mechanism involving the induction of DNA damage. In agreement with this suggestion, studies showed that AJ-5, a binuclear palladacycle complex with 1,2-bis(diphenylphosphino)ethane as co-ligand, treatment increased the level of γH2AX, a marker of double-strand DNA breaks, in different types of cancer cells3,4. Furthermore, numerous reports suggested that the anti-tumor activity of palladium complexes results from the induction of different mechanisms such as apoptosis and autophagy4. Similar findings were also reported for other palladium compounds. For example, Biphosphinic Paladacycle Complex (BPC11) activated both intrinsic apoptosis and autophagy in melanoma cells7. However, inhibition of BPC11 induced autophagy potentiated the cytotoxic effect of BPC11 which indicates that autophagy may be acting as a mechanism of melanoma cell resistance in this case. However, there is a controversy as to whether the activation of autophagy in response to palladium treatments promotes or inhibits cell death. The current study described the antitumor activity of the newly synthesized palladium compound ASH10 in ER-negative HCC1937 and ER-positive MCF7 breast cancer cells.
MATERIALS AND METHODS
Study area: The study was carried out from Jun 2018 to Sep 2019. The chemical synthesis and characterization of ASH10 took place at the Chemistry Department, Faculty of Science, Ain Shams University, Egypt. The biological experiments were performed at the Department of Biology and Biotechnology, Islamic University of Gaza.
Chemistry: Reactions were carried out without precautions to exclude light, atmospheric oxygen and moisture unless otherwise stated. Melting points were determined on a Reichert apparatus. Elemental analyses were carried out with a Carlo Erba 1106 microanalyzer results agreed favorably with IR calculated values. IR spectra were recorded on a Perkin-Elmer 16F PC FT-IR spectrometer (ESCA System, Waltham, MA, USA) with Nujol mulls between polyethylene sheets or KBr pellets. NMR spectra were recorded on a Bruker AC 200, Avance 300 or a Varian Unity 300 spectrometer at room temperature unless otherwise stated. Chemical shifts were referenced to TMS (1 H and 13 C (1 H) and H3PO4 (31P). The NMR probe temperature was calibrated using ethylene glycol 1 H NMR standard methods. Chromatographic separations were carried out by TLC on silica gel (70-230 mesh).
Preparation and characterization of ASH100: The compound ASH10 (Fig. 1) was prepared as described earlier8.
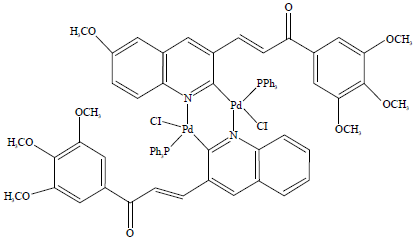 | |
| Fig. 1: | Structural formula of ASH10 complex (C79H68Cl2N2O9P2Pd2) |
A mixture of [Pd(dba)2] (432 mg, 0.75 mmol), triphenylarsine (459.3 mg, 1.5 mmol) and of 3-(2-chloro-6-methoxyquinolin-3-yl)-1-(3,4,5-trimethoxy phenyl)prop-2-en-1-one (143 day) (310.38 mg, 0.75 mmol) was mixed under N2 in dry acetone (30 mL). The reaction mixture was stirred for 3-5 hrs at room temperature, then was concentrated and CH2Cl2 was added. The solution was then passed through a pad of silica gel-MgSO4 (3:1) in a fritted funnel and then evaporated under reduced pressure to dryness and washed with Et2O, filtered to give pale brown solid product (348 mg, 41% yield). TLC (ethanol: pet. ether = 1:1), Rf = 0.739; m.p : 164-166 ºC; IR(ʋ, cm-1): ʋ(C = O) 1660, ʋ(C = N, C = C) 1620, 1585, ʋ(C-O) 1255, 1125, ʋ(C-H aliphatic) 2925, ʋ( = C-H) 2990 and ʋ(C-H aromatic) 3055 cm–1. 1H-NMR (300 MHz, CDCl3): δ 8.25-8.30 (day, 1H, = CH-CO), δ 7.16-7.108 (day, 1H,Ar-CH=), δ 4.02 (sec, 6H, m-OCH3), δ 4.25 (sec, 3H, Quinoline-OCH3), δ 3.71 (sec, 3H, p-OCH3), δ 8.65 (sec, 1H, Quinoline-H4), δ 7.85-7.83 (day, 1H,Quinoline-H8), δ 7.65 (sec, 1H, Quinoline-H5) and δ 7.55-7.35 (m, 33H, Ar-H)8.
Cell culture and treatments: Human triple-negative breast cancer cell line HCC1937 obtained from Prof Adrienne L Edkins, Rhodes University, Grahamstown, South Africa and human breast adenocarcinoma (ER+) MCF7 from Dr. Mazen ALzaharna, Islamic University of Gaza, Palestine. Cells were maintained in Dulbecco's modified Eagle's medium (RPMI 1640) (Biological Industries, Cat #01-106-1A). All media were supplemented with 10% fetal bovine serum (FBS) and 1% Penicillin-Streptomycin Solution) (Biological Industries, Cat #03-031-1B). Cells were maintained at 37°C in a 5% CO2-95% air-humidified incubator. Autophagy inhibitor 6-Amino-3-methylpurine (3MA) (10 mM) (Sigma-Aldrich, Cat#M9281-100MG) was added 1 hr before the treatment with ASH10. The ASH10 complex was dissolved in DMSO and used at different concentrations ranging from (0.0-50 μM).
Cytotoxicity assays: To determine the cytotoxic effect of ASH10, breast cancer cells were seeded at a density of (7000 cells/well) and treated after 24 hr with a range of ASH10 concentrations (0-50 μM) or vehicle for 48 hrs. Cell viability was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenylte-trazolium bromide (MTT) assay according to the manufacturer’s instructions (Santa Cruz, Cat No. sc-359848A). Briefly, 10 μL of MTT solution was added to each well and incubated for 4 hrs at 37°C. This was followed by the addition of 100 μL solubilization buffer and incubated overnight at 37°C. Absorbance at 585 nm was determined for each well and the mean cell viability was calculated as a percentage of the mean vehicle control by using automated ELIZA Analyzer (CF-fiocchetti, Italy). Three independent experiments were performed to determine the IC50 of ASH10. IC50 values were calculated by a nonlinear regression based on sigmoidal dose-response (variable slope), using GraphPad Prism 5.0 (GraphPad software, Inc., USA). The IC50 values were calculated from the linear equation from Microsoft office excel as described earlier9.
Cell migration assay: Cells were grown to 90-100% confluence in 35 mm tissue culture dishes. A linear wound was made by scratching through the monolayer using a sterile 200 μL pipette tip. To remove cell debris, the growth medium was replaced after the cells were washed 2-3 times with phosphate-buffered saline (PBS) and treated with ASH10. Then the plates were incubated at 37°C in 5% CO2. Several markings were made along the edges of the scratch line, which were used as reference points and the wound widths measured at the time of the scratching (0 hr) and thereafter at 24 and 48 hrs intervals after scratching. Pictures were taken using a phase-contrast microscope and Image J software was used to measure the wound width.
Western blotting: Cells were harvested and the protein was prepared as described previously10. The protein concentration for each cell extract determined using the BCA Protein Assay kit (Pierce, USA), with bovine serum albumin as the standard. Equal amounts of protein loaded in each lane and resolved on 6-15 % SDS-PAGE gels, then transferred electrophoretically to a Hybond ECL nitrocellulose membrane (Amersham Biosciences, USA). Membranes blocked for 1 hr at room temperature with PBS containing 5% non-fat dry milk and probed with appropriate primary antibodies O/N at 4°C with shaking. Membranes washed in PBS containing 0.1% Tween 20 (PBS/T) and incubated either with donkey anti-goat (Santa Cruz Biotechnology, CA, USA), goat anti-mouse or goat anti-rabbit IgG peroxidase-conjugated secondary antibodies (1:5000) (BioRad, Hercules, CA, USA) in blocking solution at room temperature with shaking for 1 hr. Membranes again washed in PBS/T and visualized by enhanced chemiluminescence (Pierce, USA) reaction detection system as previously described11. The following antibodies were used: anti-PARP1/2 (sc-7150), anti-p53 (sc-126), anti-p21 (sc-756), anti-ATM (sc-23921), anti-caspase 9 (sc-56076), anti-α-Tubulin (sc-8035) (Santa Cruz, California, USA), anti-p38 (M0800) (Sigma, St. Louis, MO, USA) and following antibodies were obtained from Cell Signaling (Boston, MA, USA): anti-phospho-H2AX (#2577), anti-phospho-ATM (#4526) and anti-LC3II (# 2775).
Detection of apoptosis: Cells were treated with ASH10 for 48 hrs and then stained with DAPI for 10 min. The cells were viewed by fluorescence microscopy (Zeiss, Germany). Data presented as the number of apoptotic cells per 100 cells in total.
Autophagy assays: Cells were transiently transfected with 2 mg mL–1 GFP-LC3 plasmid using Lipofectamine (Invitrogen, Carlsbad, CA, USA) according to manufacturers’ instructions3. Twenty four hours post-transfection, cells were visualized by fluorescence microscopy using an Axiovert fluorescent microscope (Zeiss, Germany) to test GFP expression. The cells were then treated with ASH10 for 48 hrs, after which they were fixed with 4% paraformaldehyde and stained with Hoechst 33342 for nuclei observation. Autophagy was quantified by counting the GFP-LC3II puncta at 200× magnification in twenty fields of view using an Axiovert fluorescent microscope (Zeiss, Germany) and divided by the total number of transfected cells within these fields and the number of GFP-LC3 puncta/cell are presented as Means±SEM of three independent experiments.
Statistical analysis: Data presented are Mean±SEM (Standard error of the means) of three independent experiments. Statistical significance was assessed between the groups using the Student’s t-test. A value of p<0.05 was accepted as statistically significant.
RESULTS
ASH10 inhibits cell proliferation and migration of human breast cancer cells: To investigate the antiproliferative effect of ASH10 on breast cancer, the MCF7 (ER-positive) and the HCC1937 (triple-negative) were treated with a range of ASH10 concentrations (0-50) μM for 48 hrs and MTT assay was used to determine cell proliferation. Results showed that ASH10 has a strong dose-dependent inhibition of cell proliferation in both breast cancer cell lines (Fig. 2a). To further explore the anti-migration properties of ASH10, a wound-healing assay was performed and a significant reduction in cell motility was observed for both cell lines exposed to 1.0 μM ASH10 for 48 hrs (Fig. 2b and c). This concentration was used to reduce the cell-killing effect of ASH10.
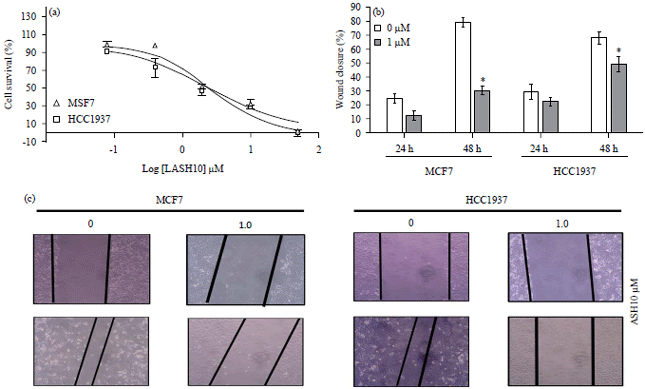 | |
| Fig. 2(a-c): | ASH10 induces cytotoxicity (a) and inhibits the migration of breast cancer cells (b and c) |
(a) Cell viability was assessed by MTT assay after 48 hr of treatment (b) Cell motility was assayed at the indicated times after the addition of either vehicle (control) or ASH10 (1.0 μM) for 48 hrs, (c) At specified time points (x-axis) cells were photographed (*p<0.001, student’s t-test) | |
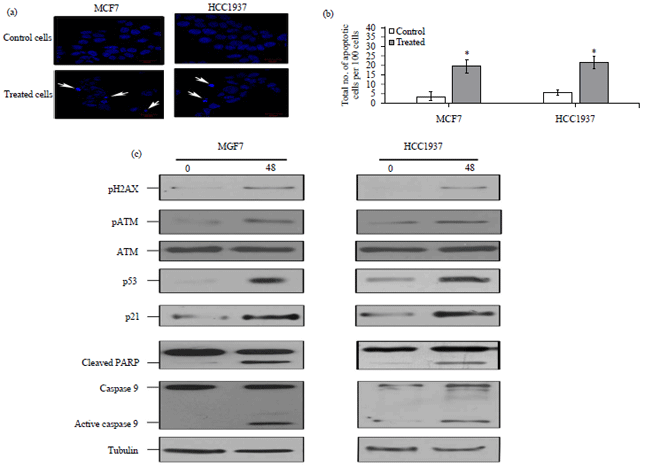 | |
| Fig. 3: | ASH10 induces (a, b) DNA fragmentation and (c) intrinsic apoptosis in breast cancer cells |
(a) Representative fluorescent photomicrographs of MCF7 and HCC1937 cells treated with 3.0 and 2.0 μM of ASH10, respectively, (b) Quantitative results for the number of apoptotic cells per 100 cells in total, Western blotting of protein from the breast cancer cells treated as in (a) and analyzed with the mentioned proteins antibodies, Tubulin was used as a loading control | |
Taken together these results demonstrate that ASH10 has potent cytotoxic and anti-migratory effects on both ER-positive and triple-negative breast cancer cells.
ASH10 induces DNA damage, cell cycle arrest and apoptosis in breast cancer cells: Figure 3a shows that ASH10 treatment induced several morphological changes associated with apoptosis such as chromatin condensation, nuclear fragmentation and nucleus margination (marked by arrows in Fig. 3a). The number of apoptotic cells per 100 cells was determined and expressed in Fig. 3b. The obtained results showed that ASH10 treatment induced significant levels (about 20%) of apoptosis in both breast cancer cells. Furthermore, immunoblotting was performed with antibodies to DNA damage markers as well as to a number of proteins involved in cycle arrest and apoptosis. Figure 3c shows that ASH10 induces DNA damage as evidenced by the induction of p-H2AX and p-ATM in both breast cancer cell lines. Under the conditions tested, there was a clear p53 response which correlated with an increase in levels of the cell cycle regulator p21. Furthermore, ASH10 treated cells showed increased levels of the intrinsic apoptosis markers, PARP cleavage and active caspase 9. Taken together these observations suggested that ASH10 induced DNA damage, cell cycle arrest and intrinsic apoptosis.
ASH10 induces autophagy in breast cancer cells: Indeed, ASH10 treatment induced a significant increase of GFP-LC3 puncta, a marker of autophagosomes, from basal levels of 8.0-29.5% in HCC1937 cells and from 1.3-15.4% in WM1158 cells (Fig. 4a). These findings were also confirmed by western blotting where ASH10 treatment induced high levels of LC3II in both cancer cell lines (Fig. 4b).
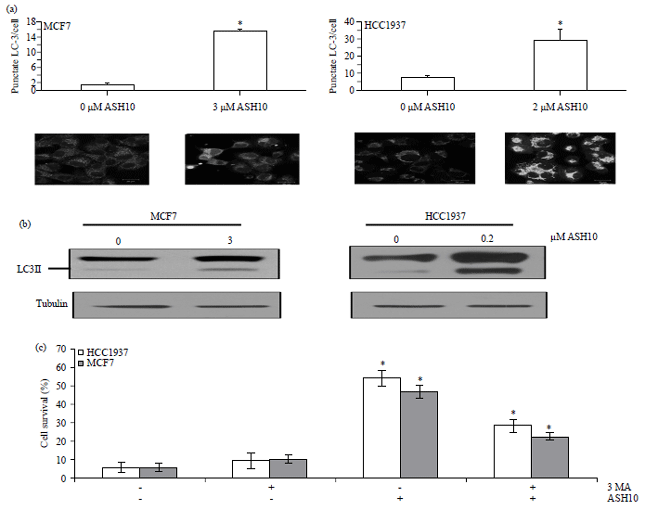 | |
| Fig. 4(a-c): | ASH10 induces (a) LC3II puncta, (b) LC3II level and (c) cytotoxicity in MCF7 and HCC1937 cells |
(a) The number of GFP-LC3 puncta/cell induced by ASH10 are presented in the graphs as Means±SEM of three independent experiments (**p<0.001, student’s t-test), (b) Western blotting of protein from cells treated as in (a) and tubulin as a loading control, (c) MTT assay showing the percentage of ASH10 cytotoxicity when cells are treated with ASH10 as in (a) in the presence or absence of 3 MA (10 μM) an autophagy inhibitor (*p<0.05, Student’s t-test) | |
Importantly, co-treatment of ASH10 with 3MA, an autophagy inhibitor, resulted in a significant reduction in ASH10 cytotoxicity as measured by MTT assay (Fig. 4c). Together, these observations indicate that ASH10 induces autophagy as a mechanism of cytotoxicity in breast cancer cells.
DISCUSSION
Metal-based compounds particularly platinum compounds have been used successfully to treat different cancers however, they are associated with severe toxicity and tumor drug resistance. Palladium based compounds have previously been shown to cause DNA damage and apoptosis in different cancer cells3,4. To understand the molecular mechanism underpinning ASH10 cytotoxicity, breast cancer cells were treated with vehicle or ASH10 (3.0 and 2.0 μM for MCF7 and HCC1937 respectively) for 48 hrs and the percentage of apoptosis was first quantified using DAPI staining. DAPI is a nuclear stain widely used to determine the level of apoptosis12. Numerous studies demonstrated that several palladium-based compounds are safer and more effective than platinum analogous particularly because they seem to be more active at lower concentrations13,14. The two platinum compounds commonly used for treating metastatic breast cancer are carboplatin and cisplatin15. Both compounds are used to treat triple-negative breast cancer (TNBC) which represents 15-20% of breast cancer cases. However, the inhibitory concentration 50% (IC50) for both compounds is higher than 10 μM in the TNBC cell line HCC193716,17. On the other hand, MCF-7 cells (estrogen receptor-positive) were more resistant to both compounds with IC50s higher than 30 μM15-17. The current study showed that after 48 hrs of ASH10 treatment, IC50 values of <3.2 μM were obtained in both HCC1937 and MCF7 cells. These observations suggest that the palladium compound ASH10 displays more potent anti-cancer activity against both types of breast cancer cells. In agreement with these results, a new palladium compound AJ-5 showed more potent effect against both types of breast cancer cells with IC50s less than 0.2 μM4. Furthermore, the current study demonstrated that ASH10 treatment significantly inhibits cell survival and migration capacity of breast cancer cells. Targeting apoptosis is one of the most common therapeutic strategies to treat cancer. However, cancer cells use different mechanisms to inhibit apoptotic cell death and to develop drug resistance. Therefore, drugs that can induce multiple mechanisms of cell death are preferred. Figure 3 shows that ASH10 induces double-strand DNA damage and intrinsic apoptosis in both cancer cell lines. These observations are similar to previous results with AJ-5, which induces intrinsic apoptosis in melanoma and breast cancer cells3,4. The current study showed that ASH10 also induces autophagy in breast cancer cells Fig. 4. There is, however, a controversy as to whether the induction of autophagy in response to chemotherapies promotes or inhibits cell death. Several studies have shown that autophagy serves as a resistance mechanism in cancer cells and that it reduces apoptosis18. For instance, inhibition of autophagy enhanced the cytotoxic effect of 5-FU, camptothecin, ursolic acid and cisplatin in different cancer cells19-23. However, there is also a growing body of evidence suggesting that autophagy may enhance cell death induced by several agents. For example, inhibition of autophagy induced by 5-FU, arsenic trioxide (As2O3) and paclitaxel induce cytotoxicity in many cancer types reversed the cytotoxic effect of these drugs24-32. The present study showed that inhibition of ASH10 induced autophagy results in a decrease of cytotoxicity which indicates that ASHI1 induced autophagy might be a mechanism of cell death.
This study provides new evidence that palladium-based compounds are promising anticancer compounds. Furthermore, it uncovers the mechanism of action by which ASH10 exerts anti-tumor activities. However, more studies are required to confirm its activities in the body and whether it has any side effects as the current study didn’t test the effect of ASH10 on any normal cells.
CONCLUSION
Taken together, the current study showed that the palladacycle ASH10 has a potent cytotoxic effect against breast cancer cells by activating both apoptosis and autophagy. This study, therefore, provides compelling evidence that ASH10 may be an effective chemotherapeutic drug in the treatment of breast cancer. Further studies are required to confirm the anticancer effect of ASH10 in vivo.
SIGNIFICANCE STATEMENT
This study discovers the anticancer properties of a new palladium based compound that can be beneficial for the development of new breast cancer treatments. This study will help the researchers to uncover the mechanism of action of similar compounds against cancer cells.
REFERENCES
- Siegel, R.L., K.D. Miller and A. Jemal, 2020. Cancer statistics, 2020. C.A. Cancer J. Clin., 70: 7-30.
CrossRefDirect Link - Aliwaini, S., J. Bleloch, S. Kimani and S. Prince, 2016. Induction of Autophagy and Apoptosis in Melanoma Treated with Palladacycle Complexes. In: Autophagy: Cancer, Other Pathologies, Inflammation, Immunity, Infection and Aging, Volume 10, Hayat, M.A. (Ed.). Chapter 12, Academic Press, New York, USA., ISBN-13: 9780128054215, pp: 231-247.
Direct Link - Aliwaini, S., A.J. Swarts, A. Blanckenberg, S. Mapolie and S. Prince, 2013. A novel binuclear palladacycle complex inhibits melanoma growth in vitro and in vivo through apoptosis and autophagy. Biochem. Pharmacol., 86: 1650-1663.
CrossRefDirect Link - Aliwaini, S., J. Peres, W.L. Kroger, A. Blanckenberg and J. de la Mare et al., 2015. The palladacycle, AJ-5, exhibits anti-tumour and anti-cancer stem cell activity in breast cancer cells. Cancer Lett., 357: 206-218.
CrossRefDirect Link - Blanckenberg, A., S. Aliwaini, S.W. Kimani, A. van Niekerk, A. Neumann-Mufweba, S. Prince and S.F. Mapolie, 2017. Preparation, characterization and evaluation of novel 1,3,5-triaza-7-phosphaadamantane (PTA)-based palladacycles as anti-cancer agents. J. Organometallic Chem., 851: 68-78.
CrossRefDirect Link - Kruszewski, M., E. Bouzyk, T. Oldak, K. Samochocka and L. Fuks et al., 2003. Differential toxic effect ofcis-platinum(II) and palladium(II) chlorides complexed with methyl 3,4-diamine-2,3,4,6-tetradeoxy-?-L-lyxo-hexopyranoside in mouse lymphoma cell lines differing in DSB and NER repair ability. Teratog. Carcinog. Mutagen., 23: 1-11.
CrossRefDirect Link - Gigli, R., G.J.S. Pereira, F. Antunes, A. Bechara and D.M. Garcia et al., 2016. The biphosphinic paladacycle complex induces melanoma cell death through lysosomal–mitochondrial axis modulation and impaired autophagy. Eur. J. Med. Chem., 107: 245-254.
CrossRefDirect Link - Elgazwy, A.S.S.H., 2008. Synthesis and characterization of pyrido[1,2-a]quinoline palladacycles. Monatsh. Chem., 139: 1285-1297.
CrossRefDirect Link - Al-Qatati, A. and S. Aliwaini, 2017. Combined pitavastatin and dacarbazine treatment activates apoptosis and autophagy resulting in synergistic cytotoxicity in melanoma cells. Oncol. Lett., 14: 7993-7999.
CrossRefDirect Link - Aliwaini, S., A.M.Awadallah, R.Y. Morjan, M. Ghunaim and H. Alqaddi et al., 2019. Novel imidazo[1,2‑a]pyridine inhibits AKT/mTOR pathway and induces cell cycle arrest and apoptosis in melanoma and cervical cancer cells. Oncol. Lett., 18: 830-837.
CrossRefDirect Link - Fulda, S. and K.M. Debatin, 2006. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene, 25: 4798-4811.
CrossRefDirect Link - Pajaniradje, S., K. Mohankumar, R. Pamidimukkala, S. Subramanian and R. Rajagopalan, 2014. Antiproliferative and apoptotic effects of Sesbania grandiflora leaves in human cancer cells. BioMed. Res. Int.
CrossRefDirect Link - Ulukaya, E., F. Ari, K. Dimas, E.I. Ikitimur, E. Guney and V.T. Yilmaz, 2011. Anti-cancer activity of a novel palladium(II) complex on human breast cancer cells in vitro and in vivo. Eur. J. Med. Chem., 46: 4957-4963.
CrossRefPubMedDirect Link - Tanaka, M., H. Kataoka, S. Yano, H. Ohi and K. Kawamoto et al., 2013. Anti-cancer effects of newly developed chemotherapeutic agent, glycoconjugated palladium (II) complex, against cisplatin-resistant gastric cancer cells. BMC Cancer.
CrossRefDirect Link - Pandy, J.G.P., J.C. Balolong-Garcia, M.V.B. Cruz-Ordinario and F.V.F. Que, 2019. Triple negative breast cancer and platinum-based systemic treatment: a meta-analysis and systematic review. BMC Cancer.
CrossRefDirect Link - Tassone, P., P. Tagliaferri, A, Perricelli, S, Blotta and B. Quaresima, 2003. BRCA1 expression modulates chemosensitivity of BRCA1-defective HCC1937 human breast cancer cells. Br. J, Cancer, 88: 1285-1291.
CrossRefDirect Link - Corkery, B., J. Crown, M. Clynes and N.O. Donovan, 2009. Epidermal growth factor receptor as a potential therapeutic target in triple-negative breast cancer. Annals Oncol., 20: 862-867.
CrossRefDirect Link - Viola, G., R. Bortolozzi, E. Hamel, S. Moro and P. Brun et al., 2012. MG-2477, a new tubulin inhibitor, induces autophagy through inhibition of the Akt/mTOR pathway and delayed apoptosis in A549 cells. Biochem. Pharmacol., 83: 16-26.
CrossRefDirect Link - Pan, X., X. Zhang, H. Sun, J. Zhang, M. Yan and H. Zhang, 2013. Autophagy inhibition promotes 5-Fluorouraci-Induced apoptosis by stimulating ROS formation in human non-small cell lung cancer A549 cells. PLoS ONE.
CrossRefDirect Link - Abedin, M.J., D. Wang, M.A. Mcdonnell, U. Lehmann and A. Kelekar, 2007. Autophagy delays apoptotic death in breast cancer cells following DNA damage. Cell Death Differ., 14: 500-510.
CrossRef - Liu, L., M. Yang, R. Kang, Z. Wang and Y. Zhao et al., 2011. DAMP-mediated autophagy contributes to drug resistance. Autophagy, 7: 112-114.
CrossRefDirect Link - Xu, Y., H. Yu, H. Qin, J. Kang and C. Yu et al., 2012. Inhibition of autophagy enhances cisplatin cytotoxicity through endoplasmic reticulum stress in human cervical cancer cells Cancer Lett., 314: 232-243.
CrossRefDirect Link - Shin, S.W., S.Y. Kim and J.W. Park, 2012. Autophagy inhibition enhances ursolic acid-induced apoptosis in PC3 cells. Biochimica et Biophysica Acta (BBA) - Mol. Cell Res., 1823: 451-457.
CrossRefDirect Link - Xiong, H.Y., X.L. Guo, X.X. Bu, S.S. Zhang and N.N. Ma et al., 2010. Autophagic cell death induced by 5-FU in Bax or PUMA deficient human colon cancer cell. Cancer Lett., 288: 68-74.
CrossRefDirect Link - Shen, Z.X., G.Q. Chen, J.H. Ni, X.S. Li and S.M. Xiong et al., 1997. Use of arsenic trioxide (As2O3) in the treatment of acute promyelocytic leukemia (APL): II. Clinical efficacy and pharmacokinetics in relapsed patients. Blood, 89: 3354-3360.
CrossRefPubMedDirect Link - Soignet, S.L., P. Maslak, Z.G. Wang, S. Jhanwar and E. Calleja et al., 1998. Complete remission after treatment of acute promyelocytic leukemia with Arsenic Trioxide. N. Engl. J. Med., 339: 1341-1348.
CrossRefDirect Link - Kanzawa, T., Y. Kondo, H. Ito, S. Kondo and I. Germano, 2003. Induction of autophagic cell death in malignant glioma cells by Arsenic Trioxide. Cancer Res., 63: 2103-2108.
Direct Link - Rousselot, P., S. Labaume, J.P. Marolleau, J. Larghero, M.H. Noguera, J.C. Brouet and J.P. Fermand, 1999. Arsenic trioxide and melarsoprol induce apoptosis in plasma cell lines and in plasma cells from myeloma patients. Cancer Res., 59: 1041-1048.
PubMedDirect Link - Jung, S.Y., S. Lee, H.B. Kang, H.A. Park and S.K. Chang et al., 2010. Antitumor activity of 3,4-dihydroquinazoline dihydrochloride in A549 xenograft nude mice. Bioorg. Med. Chem. Lett., 20: 6633-6636.
CrossRefDirect Link - Andreopoulou, E. and J.A. Sparano, 2013. Chemotherapy in patients with anthracycline and taxane-pretreated metastatic breast cancer: an overview. Curr. Breast Cancer Rep., 5: 42-50.
CrossRefDirect Link - Xi, G., X. Hu, B. Wu, H. Jiang, C.Y.F. Young, Y. Pang and H. Yuan, 2011. Autophagy inhibition promotes paclitaxel-induced apoptosis in cancer cells. Cancer Lett., 307: 141-148.
CrossRefDirect Link - Veldhoen, R.A., S.L. Banman, D.R. Hemmerling, R. Odsen and T. Simmen et al., 2013. The chemotherapeutic agent paclitaxel inhibits autophagy through two distinct mechanisms that regulate apoptosis. Oncogene, 32: 736-746.
CrossRefDirect Link