
G.G. Akunna
Department of Anatomy, Faculty of Basic Medical Sciences, Federal University Ndufu Alike Ikwo, Ebonyi State, Nigeria
E.N. Obikili
Department of Anatomy, Faculty of Basic Medical Sciences, University of Nigeria, Enugu Campus, Nigeria
G.E. Anyanwu
Department of Anatomy, Faculty of Basic Medical Sciences, University of Nigeria, Enugu Campus, Nigeria
E.A. Esom
Department of Anatomy, Faculty of Basic Medical Sciences, University of Nigeria, Enugu Campus, Nigeria
International Journal of Cancer Research
Year: 2016 | Volume: 12 | Issue: 3-4 | Page No.: 176-187







ABSTRACT
Background and Objective: One of the several side effects of cancer treatment remains cytotoxicity. The curative effect of aqueous zest extract of Citrus paradisi on cisplatin-induced testicular degeneration was studied. Materials and Methods: Sixteen male Wistar rats (10-12 weeks old) weighing 237-276 g were divided into four groups. The first group of rats served as control and were treated orally with 2.5 mL kg–1 b.wt day–1, group B rats treated with single dose of 10 mg kg–1 b.wt., group C and D rats were treated with a single dose of 10 mg kg–1 b.wt. of cisplatin and then treated orally with 10 and 40 mg kg–1 b.wt. of aqueous zest extract of Citrus paradisi for 8 weeks. Results: Results showed a significant (p<0.05) decrease in testis weight, testis volume, sperm count, sperm motility and normal sperm morphology (p<0.01). Also, observed was marked degeneration and atrophied tubules with absence of late stage germ cells evidenced by significant reduction in tubular diameter (p>0.01), perimeter (p>0.01), length (p>0.001) and width (p>0.05) in germinal epithelia height (p>0.05), cross-sectional area, number of profiles per unit area and numerical density, PAS-positive materials and Ki67 cells of seminiferous tubules of the rats treated with cisplatin alone. Interestingly, all these parameters were attenuated in the groups that were post-treated with the extract. Conclusion: Based on these evidenced it was concluded that aqueous zest extract of Citrus paradisi have curative roles in the abatement of cisplatin-induced testicular toxicity in Wistar rats.
PDF Abstract XML References Citation
Received: July 23, 2016;
Accepted: August 29, 2016;
Published: September 15, 2016
How to cite this article
G.G. Akunna, E.N. Obikili, G.E. Anyanwu and E.A. Esom, 2016. Spermiographic, 2 and 3-dimensional Quantitative Analysis of
Testicular Tissues of Rat Submitted to Citrus paradisi Waste
Extract and Cisplatin-induced Cytotoxicity. International Journal of Cancer Research, 12: 176-187.
DOI: 10.3923/ijcr.2016.176.187
URL: https://scialert.net/abstract/?doi=ijcr.2016.176.187
DOI: 10.3923/ijcr.2016.176.187
URL: https://scialert.net/abstract/?doi=ijcr.2016.176.187
INTRODUCTION
The surface area of an interface per unit reference volume is known as surface density or specific surface1,2. Three dimensional evaluation of an organ describes numerical sampling methods for quantifying geometric parameters.
Using scientifically proven relationships, this study has aimed to evaluate in animal models, the curative role of aqueous zest extract of Citrus paradisi (AZECP) on cisplatin-induced cytotoxicity.
Citrus peel (especially orange) is often considered a waste from the main fruit which boasts of essential flavanones, polyphenols, anthocyanins and hydroxycinnamic acids. However, citrus peel has been reported to contain higher amounts of total phenolics such as limonene, hesperidin, narirutin, naringin and eriocitrin compared to the edible portions3-5. Their pharmacological activities as radical scavengers have been reported.
Cisplatin (cis-diamminedichloroplatinum [II]) is one of the standard anti-neoplastic drugs used in cancer therapy6,7. However, it results in spermiotoxicity thereby leading to temporary or permanent azoospermia8,9. Experimental models have severe testicular damage within days of cisplatin injection10-12. Free radical generation via an oxidative stress pathway has been implicated in cisplatin-induced testicular toxicity13. The final cellular outcome is generally apoptotic cell death.
Although, several herb has been tried to protect against drug-induced toxicity. This study has aimed to see one to cure the reproductive side effects accompanied by cisplatin chemotherapy.
MATERIALS AND METHODS
Plant source and identification: Fresh fruits of Citrus paradisi were collected from a cultivated farm land in Ikwo, Ebonyi State, Nigeria and was identified and authenticated (UNN/14/344) at the Department of Plant Science and Biotechnology, Faculty of Science, University of Nigeria, Nsukka by a botanist.
Preparation of aqueous zest extracts of Citrus paradisi (AZECP): Five hundred fruits were peeled with a zester or grater. The white portion of the peel under the zest (Mesocarp) was generally avoided by limiting the peeling depth14. The zest was thoroughly rinsed in distilled water and dried at room temperature about 2 weeks. It was then reduced to a powdered form by grinding.
Aqueous extraction of AZECP: Aqueous infusion was done by mixing a calculated volume of distilled water and powdered sample. The mixture was allowed to stand for 30 min before filtration. It was then centrifuged at about 3000×g for 5 min and the supernatant collected. The supernatant was cleared of particles by suction filtration using Whatmann No. 1 filter paper and cellulose filter paper. The extract was subsequently concentrated to dryness in vacuo at 40oC using a rotary evaporator and stored in a dessicator. Fresh solution of the extract was prepared in normal saline as vehicle when required15.
Chemicals: Anti-neoplastic agent cisplatin (cis-diamminedichloroplatinum [II]) was obtained from Alpha pharmacy and stores at 44, Ogui Road, Enugu, Enugu Nigeria.
Animal grouping and investigational procedure: Twenty male Wistar rats (10-12 weeks old) weighing 237-276 g were secured from the animal house of the Department of physiology, University of Nigeria, Enugu Campus. The animals were housed in well ventilated wire cages in the animal facility and an ethical approval was sorted and obtained from the Ethical Committee on animal use with reference NNHREC/05/01/2008b-FWA00002458-1RB00002323.
The animals were divided into four groups of five rats each. The rats in group A served as the control group and were treated orally with 2.5 mL kg–1 b.wt day–1, group B rats served as the positive control group and was given single dose of 10 mg kg–1 b.wt. of CIS intraperitoneally11, group C and D rats were given a single dose of 10 mg kg–1 b.wt. of CIS intraperitoneally, 2 weeks later they were given 10 and 40 mg kg–1 b.wt. of AZECP daily, respectively for 2 weeks 24 h later. The procedure lasted for 8 weeks.
Experimental procedures involving the animals and their care were conducted in conformity with international, national and institutional guidelines for the care of laboratory animals in Biomedical Research and Use of Laboratory Animals in Biomedical Research as promulgated by the Canadian Council of Animal Care16.
Further the experimental models used conformed to the guiding principles for study involving animals as recommended by the declaration of Helsinki and the guiding principles in the care and use of animals17.
Animal sacrifice and sample collection: The rats were first weighed and then sacrificed by cervical dislocation. The abdominal cavity was opened up through a midline abdominal incision to expose the reproductive organs. The testes were excised and trimmed of all fat. The testicular weights of each animal were evaluated with an electronic analytical and precision balance (BA 210S, d = 0.0001-Sartorius GA, Goettingen, Germany). The testes volumes were measured by water displacement method. The two testes of each rat were measured and the average value obtained for each of the two parameters was regarded as one observation. One of the testes of each animal was fixed in bouin’s fluid for histological and morphometric analysis.
Determination of epididymal sperm parameters
Spermatozoa progressive motility: The fluid obtained from the left cauda epididymis with a pipette was diluted to 0.5 mL with tris buffer solution18 (pH 7.5). A slide was placed on light microscope with heater table and an aliqout of this solution was on the slide. The microscopic field was scanned systematically and each spermatozoon encountered was assessed and for the purpose of the first part of the assessment, motility was classified as either motile or non-motile and percentage motility was evaluated visually at a magnification of x400. Motility estimates were performed from three different fields in each sample. The mean of the three estimations was used as the final motility score. Samples for motility evaluation were be incubated at 35°C. Further, motile spermatozoa was classified as either rapid linear progressive or sluggish non-linear motile.
Epididymal spermatozoa concentrations: Spermatozoa in the right epididymis were counted by a modified method of Yokoi and Mayi19. The epididymis was minced with anatomic scissors in 5 mL physiologic saline, placed in a rocker for 10 min and allowed to incubate at room temperature for 2 min. After incubation, the supernatant fluid was diluted 1:100 with solution containing 5 g sodium bicarbonate and 1 mL formalin (35%). Total sperm number was determined by using the new improved Neubeur’s counting chamber (haemocytometer). Approximately, 10 μL of the diluted sperm suspension was transferred to each counting chamber of the haemocytometer and was allowed to stand for 5 min. This chamber was then placed under a binocular light microscope using an adjustable light source. The ruled part of the chamber was focused and the number of spermatozoa counted in five of 16-celled squares. The sperm concentration was the calculated number multiplied by 5 and expressed as:
[x]×106 mL–1
where, [x] is the number of spermatozoa in a 16-celled square.
Normal and abnormal spermatozoa morphology: The sperm cells were evaluated with the aid of light microscope at x400 magnification. Caudal sperm will be taken from the original dilution for motility and diluted 1:20 with 10% neutral buffered formalin (Sigma-Aldrich, Oakville, ON, Canada). Five hundred sperm from the sample were scored for morphological abnormalities. Briefly, in wet preparations using phase-contrast optics, spermatozoa were categorized. In this study, spermatozoon was considered abnormal morphologically if it had one or more of the following features: Rudimentary tail, round head and detached head and will be expressed as a percentage of morphologically normal sperm20.
Tissue preparation for histology, histochemistry and immunohistochemistry: The organs were cut in slabs of about 0.5 cm thick and fixed in Bouin’s fluid for a day after which it was transferred to 70% alcohol for dehydration. The tissues were passed through 90% alcohol and chloroform for different durations before they were transferred into two changes of molten paraffin wax for 20 min each in an oven at 57°C. Serial sections of 5 μm thick were obtained from a solid block of tissue and were stained with haematoxylin and eosin stains after which they were passed through a mixture of equal concentration of xylene and alcohol.
Following clearance in xylene and the tissues were oven-dried. Light microscopy was used for the evaluations. Prior to embedding, it was ensured that the mounted sections to be cut by the rotary microtome were orientated perpendicular to the long axes of the testes. These sections were designated "Vertical sections". Serial sections of 4 μm thick were obtained from a solid block of tissue, fixed on clean slides to which Mayer’s egg albumin had been coated to cement the sections to the slides properly and were stained. For histological and histochemical study, sections were stained with H and E and Periodic Acid-Schiff (PAS) reaction with hematoxylin counterstaining after which they were passed through a mixture of equal concentration of xylene and alcohol. Following clearance in xylene the sections will be oven-dried21 between 35-40°C.
For immunohistochemical study, sections of testes were deparaffinized with xylene and followed by antigen retrieval by heating in citrate buffer (10 mM and 20 min). This was followed by endogenous peroxidase blocking in 3% H2O2 for 10 min and incubation with rabbit antimouse Ki67 monoclonal antibody (1:300; Millipore, AB9260). After washing the slides with phosphate buffered saline, the sections were incubated with their related secondary antibodies at room temperature for 1 h followed by detection with 3-amino-9-ethylcarbazole and a chromogen. The slides were counterstained with hematoxylin and mounted in Faramount aqueous mounting medium. Cellular proliferation was assessed by immunohistochemical detection of the Ki67 nuclear antigen. Cells that stained positive for Ki67 were counted in each sample. One hundred tubules were counted in each group and the percentage of positive cells in the tubules was determined. The slides was viewed under a research microscope connected to a computer monitor for qualitative and quantitative evaluation.
Determination of morphometric parameters: For each testis, seven "Vertical sections" from the polar and the equatorial regions were sampled22. Seven "Vertical sections" per testis was selected by a systematic sampling method that ensured fair distribution between the polar and equatorial regions of each testis. Briefly, a section was taken at the equator of each testis, one on each side of the equator, three-quarters of the distance between the pole and the equator, another half-way between each pole and the equator and one on each side of the equator and a quarter of the distance from each of the pole.
Diameter, perimeter, length, width, roundness, lumen diameter and germinal epithelia height of seminiferous tubules of the testes were estimated with a digimizer software programme. Unbiased numerical estimation of the following morphometric parameters was determined using a systematic random scheme23: Cross-sectional area of the seminiferous tubules (AC); number of profiles of seminiferous tubules per unit area of testis (NA) and numerical density of the seminiferous tubules (NV).
For each stereological parameter (D, AC, NA and NV) five randomly selected fields from all the seven sections of a single testis was viewed and estimation on each carried out. The average from a total of 70 readings from five fields in seven sections of the two testes of one rat was obtained and this was recorded as one observation. The evaluation of the diameter was done with calibrated eyepiece and stage grids mounted on a light research microscope at x100 magnification. Estimation of volume density of testicular components and number of seminiferous tubules was done on a computer monitor onto which a graph sheet was superimposed and on which slides were projected from a study light microscope (olympus).
Cross-sectional area (AC) of the seminiferous tubules of the testes: The cross-sectional areas of the seminiferous tubules was determined from the equation:
![]()
where, π is equivalent to 3.142 and D the mean diameter of the seminiferous tubules.
Number of profiles of seminiferous tubules in a unit area of testis (NA): The number of profiles of seminiferous tubules per unit area was determined by using the unbiased counting frame proposed by Gundersen and Jenson23. Using this frame, in addition to counting profiles completely inside the frame counted all profiles with any part inside the frame provided they did not touch or intersect the forbidden line (full-drawn line) or exclusion edges or their extension.
Numerical density (NV) of seminiferous tubules: This is the number of profiles per unit volume and was determined by using the modified Floderus equation:
NV = NA/(D+T)
where, NA is the number of profiles per unit area, D is the diameter and T the average thickness of the section.
Statistical analysis: The data were statistically analyzed and expressed as Mean±SD. Analysis was carried out using analysis of variance (ANOVA) with Scheffe’s post hoc test. The level of significance was considered at p<0.05 and p<0.01.
RESULTS
Testicular weight and volume: Results showed a significant (p<0.05) decrease in testis weight and volume of rat treated with CIS-alone. However, testis volume was not significantly (p>0.05) affected when compared to the negative control group (Fig. 1).
Group C animals had significant increase in testes weight but not testes volume while group D animals had a significant increase in test volume but not testes weight when compared to the positive control group.
Testes histological profiles: Rats that were treated with CIS-alone without pre-treatment or post-treatment with AZECP showed marked degeneration and atrophied seminiferous tubules, interstitial oedema, degenerated and vacuolated germinal epithelium, absence of late stage germ cells, degenerated spermatogenic cells and absence of sperm bundles in most tubules when compared to those of the control that had normal histological profiles (Fig. 2b).
The rats that had AZECP post-treatment showed a remarkable preservation of their seminiferous epithelium (Fig. 3c, d).
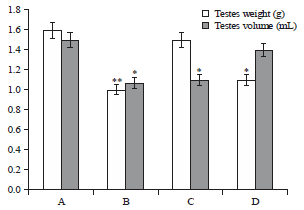 | |
| Fig. 1: | Effects of CIS and aqueous zest extract of Citrus paradisi on testicular weight and volume of male Rats, *Significant increases or decreases at p<0.05 when compared to negative control (group A), AZECP: Aqueos zest extract of Citrus paradisi, CIS: Cisplatin, NS: Normal saline, values are Mean±SD and n = 5 in each group |
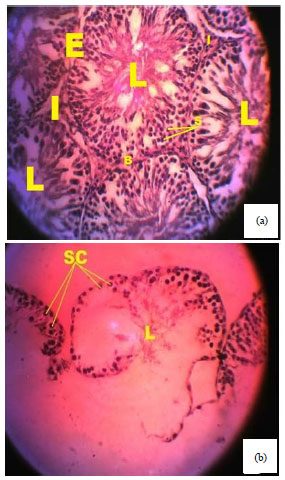 | |
| Fig. 2(a-b): | Histological micrograph of a cross-section of testis of group A rat (5 mL kg–1 NS) and group B (10 mg kg–1 CIS) for 8 weeks, stain: H and E, slide showing the seminiferous tubules, L: Lumen, E: Epithelium, I: Interstitium, S: Sertoli cells, L: Leydig cell, stain: Haematoxylin eosin and magnification: x400 |
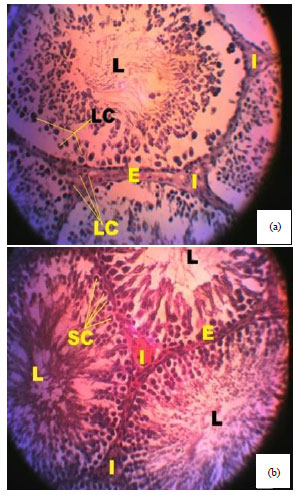 | |
| Fig. 3(a-b): | Histological micrograph of a cross-section of testis of group C (10 mg kg–1 CIS+10 mg kg–1 AZECP) and group D (10 mg kg–1 CIS+40 mg kg–1 AZECP) for 8 weeks, stain: H and E, slide showing the seminiferous tubules, L: Lumen, E: Epithelium, I: Interstitium, S: Sertoli cells, L: Leydig cell, stain: Haematoxylin, eosin and magnification: x400 |
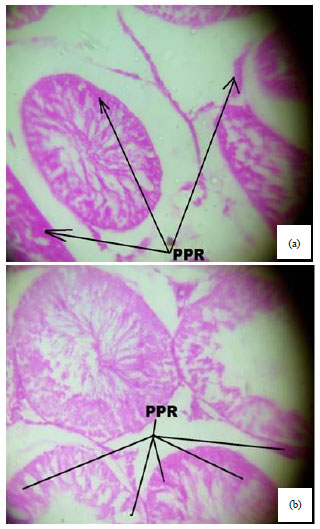
Testes histochemical and immunohistochemical profiles: The testes of rat treated with CIS without post-treatment with AZECP presented a decrease of PAS-positive materials. In these specimens, tunica albuginea, the boundaries of the seminiferous tubules as well as the intertubular connective tissue had weak PAS-positive materials (Fig. 4b).
Normal polysaccharides content were illustrated after post-treatment with AZECP (Fig. 4c, d). Numerous PAS-positive materials appeared in tunica albuginea as well as in the intertubular connective tissue of testes of control groups of rat (Fig. 4a). It showed normal cytoplasmic carbohydrate supplement in nearly all of the cells which were participated in spermatogenesis process.
Although, there was a significant decrease in Ki67 positive cells in the positive control group an d this was improved after post-treatment with AZECP (Fig. 5-8).
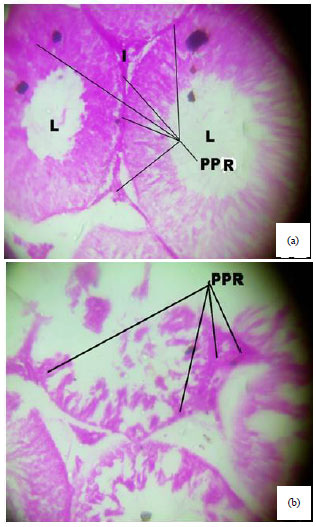 | |
| Fig. 4(a-b): | Histochemical micrograph of a cross-section of testis of group A rat (5 mL kg–1 NS), group B (10 mg kg–1 CIS) for 8 weeks, stain: H and E, slide showing the seminiferous tubules, L: Lumen, PPR: PAS-positive reactions, stain: PAS and magnification: x400 |
The activity of Ki67 appeared to increase in the control group (Fig. 5a, b) of rat. Ki67-stained cells were mainly localized in the spermatogonial germ cells in seminiferous tubules. Some Ki67-positive cells were observed in the interstitium.
Testes histo-morphometry: Diameter, perimeter, width (WST), roundness (RST) and lumen diameter (LD), germinal epithelia height (GEH), cross-sectional area (Ac) number of profiles per unit area (NA), numerical density (NV) and length of seminiferous tubules of Wistar rats.
The negative control group of rat had normal geometric parameters (mean diameter (278.5±91.6 μm), perimeter (900.4±121.0 μm) and length (342.7±72.3 μm) of the seminiferous tubules (Table 1).
 | |
| Fig. 5(a-b): | Histochemical micrograph of a cross-section of testis of group C (10 mg kg–1 CIS+10 mg kg–1 AZECP), group D (10 mg kg–1 CIS+40 mg kg–1 AZECP) for 8 weeks, stain: H and E, slide showing the seminiferous tubules, L: Lumen, PPR: PAS positive reactions, stain: PAS and magnification: x400 |
The group that had CIS-alone had a significant decrease in tubular diameter (p>0.01), perimeter (p>0.01) and length of the seminiferous tubules (p>0.001) when compared to the negative control.
Although, group D rats had a significant (p>0.05) reduction in tubular diameter when compared to the control, it was significant (p>0.05) different from the positive control group.
The animals that were treated with AZECP after exposure to CIS showed significant changes in their tubular diameter when compared to the positive control (Table 1).
The width, roundness and lumen diameter of the seminiferous tubules in the control rats were 232.7±24.6, 0.84±0.1 and 87.9±10.9 μm, respectively (Table 2).
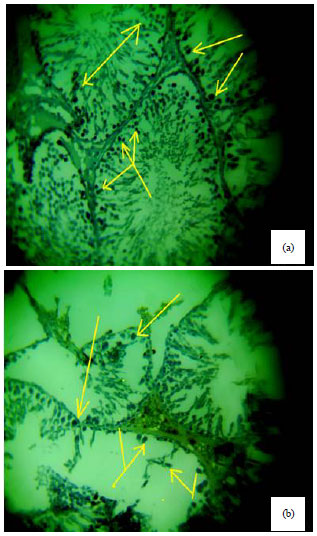 | |
| Fig. 6(a-b): | Immunohistochemical micrograph of a cross-section of testis of group A rat (5 mL kg–1 NS), group B (10 mg kg–1 CIS) for 8 weeks, stain: H and E, slide showing the seminiferous tubules, L: Lumen, PPR: PAS-positive reactions, Ki67 immunostaining and magnification: x400 |
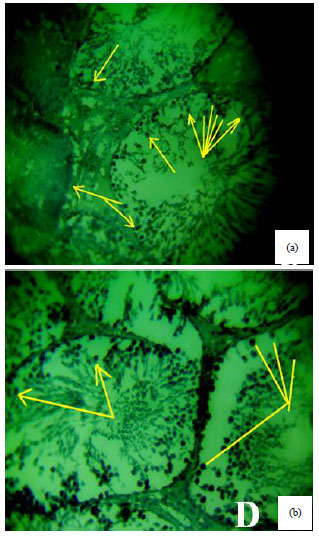 | |
| Fig. 7(a-b): | Immunohistochemical micrograph of a cross-section of testis of group C (10 mg kg–1 CIS+10 mg kg–1 AZECP) and group D (10 mg kg–1 CIS+40 mg kg–1 AZECP) for 8 weeks, stain: H and E, slide showing the seminiferous tubules, L: Lumen, PPR: PAS-positive reactions, Ki67 immunostaining and magnification: x400 |
| Table 1: | Effects of cisplatin and aqueous zest extract of Citrus paradisi on diameter (D), perimeter of seminiferous tubules (PST) cross-sectional area (Ac), number of profiles per unit area (NA) and length of seminiferous tubules (LST) of Wistar rats |
 | |
a,bSignificant increases or decreases at p<0.05 and p<0.01, respectively when compared to negative control (group A), values are Mean±SD, n = 4 in each group and *Significant increases or decreases at p<0.05 when compared to positive control | |
| Table 2: | Effects of cisplatin and aqueous zest extract of Citrus paradisi on width (WST), roundness (RST) germinal height (GEH), numerical density (NV) and lumen diameter (LD) of seminiferous tubules of Wistar rats |
 | |
| a,bSignificant increases or decreases at p<0.05 and p<0.01, respectively when compared to negative control (group A), values are means±SD and n = 5 in each group | |
| Table 3: | Effects of cisplatin and orange peel extract on sperm count, motility, progressivity and morphology in Wistar rats |
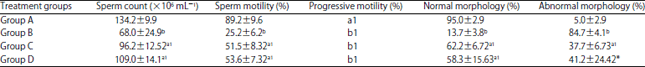 | |
a,bSignificant increases or decreases at p<0.05 and p<0.01, respectively when compared to negative control (group A), *Significant increases or decreases at p<0.05 when compared to positive control a1: Rapid linear progressive motility and b1: Sluggish linear or non-linear motility | |
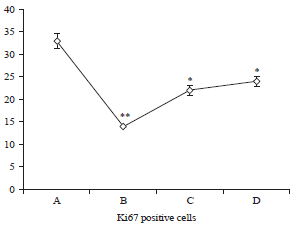 | |
| Fig. 8: | Effects of CIS and aqueous zest extract of Citrus paradisi on Ki67 Positive cells of male Rats, *’**Significant increases or decreases at p<0.05 and p<0.05, respectively when compared to negative control (group A), AZECP: Aqueos zest extract of Citrus paradisi, CIS: Cisplatin, NS: Normal saline, values are Mean±SD and n = 5 in each group |
There was a significant reduction (p>0.05) in width and increase (p>0.05) in lumen diameter of the tubules in rats treated with CIS (200.9±31.6 and 190.4±59.6 μm, respectively) (Table 2).
The animals that had AZECP after CIS treatment had significant increase in width and lumen diameter of the tubules when compared to the positive control group.
The mean number of germinal epithelia height, cross-sectional area, number of profiles per unit area and numerical density of seminiferous tubules in negative control groups were 25.3±2.8 μm, 38.8±6.33 Ac (×103 μm3), 36.26±6.9 NA (×10‾8 μm–2) and 17.16±4.9 (×10–10 μm–2), respectively (Table 1, 2).
However, there was a significant (p<0.05) increase in germinal height, cross-sectional area, number of profiles per unit area and numerical density in rats treated with AZECP after CIS when compared to the positive control (Table 1, 2).
Sperm parameters
Sperm count and motility in Wistar rats: Although, the mean sperm count and motility for the animals treated with only normal saline (the negative control groups) were normal at 134.2±9.9×106 mL–1 and 89.2±9.6%, respectively, results showed a significant (p>0.01) reduction in sperm count and motility in the group treated with CIS without post-treatment with AZECP (Table 3).
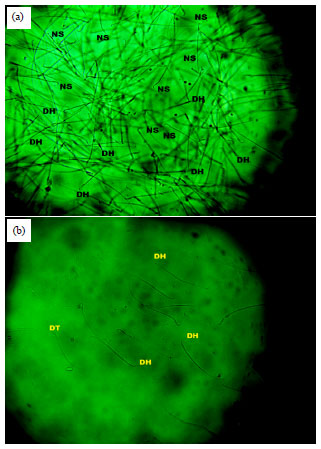 | |
| Fig. 9(a-b): | Sperm morphology of group A rat (5 mL kg–1 NS), group B (10 mg kg–1 CIS) for 8 weeks, RH: Round head, DH: Detached (double tail), RT: Round tail and TB: Twisted body |
Although, there was a significant decrease (p<0.05) in sperm count and motility in animals treated with AZECP after CIS when compared to the negative control, these values were significant (p<0.05) different from the positive control group (Table 3).
The result of normal sperm morphology and abnormal sperm morphology were in line with that of the count and motility. However, the sperm progressivity in all treated rat were different from those of the negative control (Table 3, Fig. 9, 10).
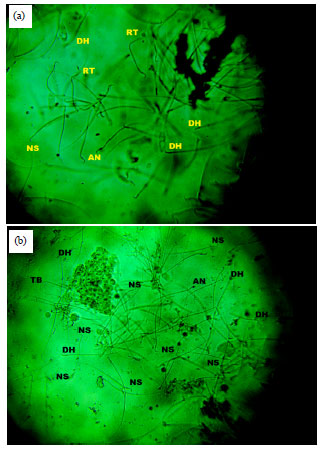 | |
| Fig. 10(a-b): | Sperm morphology of group C (10 mg kg–1 CIS+10 mg kg–1 AZECP) and group D (10 mg kg–1 CIS+40 mg kg–1 AZECP) for 8 weeks, RH: Round head, DH: Detached (double tail), RT: Round tail and TB: Twisted body |
DISCUSSION
The use of CIS, a potent anti-neoplastic drug with efficacy in a broad range of malignancies is limited by severe cytotoxic side effects24,25. Sperm cell is one of the main targets to a certain extent because of their elevated mitotic activities20. Animal-based studies have improved understanding the underlying processes and have also been the basis of study concerning protection and restoration of reproductive potential.
The animals that were treated with CIS-alone had testicular volumes of rat treated with CIS-alone had a significant decrease in testicular weight and testicular volume. It has been suggested that derangement in seminiferous tubules and leydig cells could lead to loss of about 70-80% of testicular mass26,27.
Although, testicular weight might appear a trivial result, this assessment has been indicated as sensitive end points that can be employed in evaluation of negative effect on male reproduction15,27. These results on these gross anatomical parameters in CIS-induced animals shown in this study have already been described by Malarvizhi and Mathur28 and can be attributed to leydig cell death which could have subsequently reduced testosterone production.
Ganaie29 showed that exposure to CIS, reduced the serum testosterone levels in experimental models. Also, it is the fact that the molecular mechanism of CIS-induced testicular toxicity is mediated by dysfunction of biosynthesis of testosterone30.
Marked degeneration and atrophied seminiferous tubules and decrease in geometric parameters, PAS-positive materials and expression of Ki-67 observed in this study are in accordance with several other finding31-35. Atessahin et al.20 provide well documented evidences of testicular morphologic and morphometric impairment following CIS challenge in animal models. Somatic cells (leydig and sertoli cells) and germ cells are the main target cells for platinum compounds such as CIS11, 36, 37.
The geometric parameters in this study go in line with the report that cisplatin-induced testicular degeneration results in vacuoles in the seminiferous epithelium38,39. In the same line, the reduction in germinal epithelium thickness and seminiferous tubular diameter might be due to the inhibition of spermatogenesis which might have resulted in decreased sperm count, motility and normal sperm morphology observed in this study.
Since, this result is comparable to that of Maines et al.40, the reduction in testosterone level may have depleted the germ cells at stages VII and VIII of seminiferous tubules. This is pertinent when one consider the report that conversion of immature spermatids to mature spermatids is considered to be thoroughly related to testosterone level41.
Cisplatin-based chemotherapy has been reported to cause infertility accompanied with significant reduction in sperm production in most testicular cancer patients40.
Oxidative stress has also been implicated in cisplatin-induced toxicity. The CIS has been reported to generate free radicals either by the enzymatic pathway of redox cycling between a semiquinone form and a quinone form or by the non-enzymatic pathway of forming a DOX-Fe3+ complex25,42.
Kim et al.43 observed that CIS could promote the increase in lipid peroxidation in vitro. Cao et al.44 also indicated that excessive oxidative stress reduced levels of key enzymatic and non-enzymatic antioxidants in leydig cells resulting in decline in testosterone synthesis and secretion44.
Sperm cell is one of the common body cells that are susceptible to CIS-induced oxidative stress. This increase oxidative stress damages the sperm membranes, proteins and DNA45,46. It has also been reported that adenosine triphosphate (ATP) is an energy source for sperm motility and its availability may be a limiting factor responsible for loss of sperm motility in CIS-treated rats47. This could explain the significant reduction in sperm concentration, sperm motility and normal sperm morphology along with a significantly increased abnormal sperm morphology rates and significantly reduced sperm progressivity as seen in CIS-alone group rats when compared to the control groups. This result is also in line with previous reports that demonstrated sperm impairment11,20,48-54. Laskey et al.55 also reported decrease in progressive motility and sperm normality.
Worthy of note is the remarkable normalization of the gross anatomical parameters, geometric parameters, sperm parameters and histomorphology in the groups of rats post-treated with AZECP. Although, testicular volume and weight are highly correlated56, the result was not in line with this as the groups treated with AZECP either had increase in weight or volume but not both. However, the results were in line with reports correlating testicular volume and testicular function30,28.
Due to the presence of antioxidant in herbal plants, they have been utilized in proteching in protecting against CIS toxicity57,58. Improved sperm motility and the function after antioxidant treatment have been reported severally20,59,60.
Ginkgo biloba, Zingiber officinale, Hibiscus sabdariffa and Curcuma longa have all been reported to reduce CIS-induced testicular toxicity61-65. Post-treatment with AZECP could have attenuated the CIS testicular derangement through a reduction of free radicals dependent lipid peroxidation.
Biochemical analysis of AZECP revealed the presence in high concentration of vitamin C, vitamin E, lycopene and polyphenolic flavonone glycosides, hesperiden, neohesperiden, nariratin, limonoid and naringin. These compounds are powerful antioxidants and free radical scavengers individually and collectively46-47.
CONCLUSION
The present study has shown that post-treatment with aqueous zest extract of Citrus paradisi containing powerful antioxidant vitamins and citrus bioflavonoids, exerted a potent testiculocurative activity against cisplatin-induced morphological, biochemical and functional impairment of the testis of Wistar rats. However, there was no notable difference in the effect of different doses.
The fact that aqueous zest extract of Citrus paradisi exerts profound curative effects on cisplatin-induced testicular toxicity in animal models warrants a further approach for its testiculocurative potential during cisplatin chemotherapy and testicular cancer in clinical practice.
SIGNIFICANCE STATEMENT
The present study has provided an addition to the body of evidence that cisplatin chemotherapy induces morphological, biochemical and functional impairment of the testis of Wistar rats.
It has also shown that post-treatment with aqueous zest extract of Citrus paradisi exerted a potent testiculocurative activity against cisplatin-induced testicular injury.
This study may have provided the first documented evidence of the efficacy of aqueous zest extract of Citrus paradisi to protect and attenuate the testicular toxicity associated with cisplatin chemotherapy and testicular neoplasm.
These results warrant a further approach for the testiculocurative potential of Citrus paradisi during cisplatin chemotherapy and testicular cancer in clinical practice.
ACKNOWLEDGMENT
The authors thank Mr. J.A. Oguntola and Dr. Egwu O.A for their help as regards to this experiment.
REFERENCES
- Yama, O.E., A.C. Ifeanyi, S.A. Oyebadejo, C.C. Noronha, S.L. Chai and A.O. Okanlawon, 2011. Three-dimensional quantitative analysis of testicular tissue sections of sprague-dawley rat: The after treatment responses to varying doses of Momordica charantia. Agric. Biol. J. North Am., 2: 1390-1394.
Direct Link - Xu, G.H., J.C. Chen, D.H. Liu, Y.H. Zhang, P. Jiang and X.Q. Ye, 2008. Minerals, phenolic compounds and antioxidant capacity of citrus peel extract by hot water. J. Food Sci., 73: C11-C18.
CrossRefDirect Link - Mohamed, N.A.E., A.A. Tohamy, B. Elgamal and A.E.A. Moneim, 2014. Ameliorative effect of citrus peel extract on castration-induced oxidative stress in liver and kidney of rats. J. Applied Pharmaceut. Sci., 4: 64-68.
Direct Link - Green, C.O., A.O. Wheatley, D.A. Mcgrowder, L.L. Dilworth and H.N. Asemota, 2013. Citrus peel polymethoxylated flavones extract modulates liver and heart function parameters in diet induced hypercholesterolemic rats. Food Chem. Toxicol., 51: 306-309.
CrossRefPubMedDirect Link - Ravindra, P., D.A. Bhiwgade, S. Kulkarni, P.V. Rataboli and C.Y. Dhume, 2010. Cisplatin induced histological changes in renal tissue of rat. J. Cell Anim. Biol., 4: 108-111.
Direct Link - Mashhadi, M.A., Z. Heidari and Z. Zakeri, 2013. Mild hypomagnesemia as the most common Cisplatin nephropathy in Iran. Iran. J. Kidney Dis., 7: 23-27.
PubMedDirect Link - Townsend, D.M., M. Deng, L. Zhang, M.G. Lapus and M.H. Hanigan, 2003. Metabolism of cisplatin to a nephrotoxin in proximal tubule cells. J. Am. Soc. Nephrol., 14: 1-10.
CrossRefDirect Link - Yao, X., K. Ichpisal, N, Kurtzman and K. Nugent, 2007. Cisplatin nephrotoxicity: A review. Am. J. Med. Sci., 334: 115-124.
CrossRefPubMedDirect Link - Malarvizhi, D. and P.P. Mathur, 1996. Effects of cisplatin on testicular functions in rats. Indian J. Exp. Biol., 34: 995-998.
PubMedDirect Link - Cherry, S.M., P.A. Hunt and T.J. Hassold, 2004. Cisplatin disrupts mammalian spermatogenesis, but does not affect recombination or chromosome segregation. Mutat. Res./Gen. Toxicol. Environ. Mutagen., 564: 115-128.
CrossRefPubMedDirect Link - Che, C.M. and F.M. Siu, 2010. Metal complexes in medicine with a focus on enzyme inhibition. Curr. Opin. Chem. Biol., 14: 255-261.
CrossRefDirect Link - Cvitkovic, E., 1998. A historical perspective on oxaliplatin: Rethinking the role of platinum compounds and learning from near misses. Semin. Oncol., 25: 1-3.
PubMed - Saalu, L.C., V.A. Togun, A.O. Oyewopo and Y. Raji, 2006. Artificial cryptorchidism and the moderating effect of melatonin (N-acetyl. 5 methoxy tryptamin) in sprague-dawley rats. J. Applied Sci., 6: 2889-2894.
CrossRefDirect Link - WMA and APS, 2002. Guiding principles for research involving animals and human beings. Am. J. Physiol.: Regul. Integr. Comp. Physiol., 283: R281-R283.
CrossRefPubMedDirect Link - Sonmez, M., G. Turk and A. Yuce, 2005. The effect of ascorbic acid supplementation on sperm quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology, 63: 2063-2072.
CrossRefDirect Link - Atessahin, A., I. Karahan, G. Turk, S. Gur, S. Yilmaz and A.O. Ceribasi, 2006. Protective role of lycopene on cisplatin-induced changes in sperm characteristics, testicular damage and oxidative stress in rats. Reprod. Toxicol., 21: 42-47.
CrossRefDirect Link - Qin, D. and M.A. Lung, 2002. Morphometric study on leydig cells in capsulotomized testis of rats. Asian J. Androl., 4: 49-53.
PubMed - Gundersen, H.J.G. and E.B. Jensen, 1987. The efficiency of systematic sampling in stereology and its prediction. J. Microsc., 147: 229-263.
CrossRefPubMedDirect Link - Cohen, S.M. and S.J. Lippard, 2001. Cisplatin: From DNA damage to cancer chemotherapy. Progr. Nucleic Acid Res. Mol. Biol., 67: 93-130.
CrossRefDirect Link - Cepeda, V., M.A. Fuertes, J. Castilla, C. Alonso, C. Quevedo and J.M. Perez, 2007. Biochemical mechanisms of cisplatin cytotoxicity. Anticancer Agents Med. Chem., 7: 3-18.
PubMedDirect Link - Setchell, B.P. and D.E. Brooks, 1988. Anatomy, Vasculature, Innervations and Fluids of the Male Reproductive Tract. In: The Physiology of Reproduction, Volume 2, Knobil, E. and J.D. Neil (Eds.). Chapter 19, Raven Press, New York, USA., ISBN-13: 9780881672817, pp: 753-836.
Direct Link - Creasy, D.M., 2003. Evaluation of testicular toxicology: A synopsis and discussion of the recommendations proposed by the Society of Toxicologic Pathology. Birth Defects Res. Part B: Dev. Reprod. Toxicol., 68: 408-415.
CrossRefDirect Link - Malarvizhi, D. and P.P. Mathur, 1995. Effect of cisplatin on physiological status of normal rat testis. Indian J. Exp. Biol., 33: 281-283.
PubMed - Ganaie, M.A., 2015. The protective effects of naringenin on testes gonadotoxicity induced by cisplatin in rats. Bull. Environ. Pharmacol. Life Sci., 5: 15-21.
Direct Link - Vawda, A.I., 1994. Effect of testosterone on cisplatin-induced testicular damage. Arch. Androl., 32: 53-57.
PubMed - Xu, B., S.E. Chia, M. Tsakok and C.N. Ong, 1993. Trace elements in blood and seminal plasma and their relationship to sperm quality. Reprod. Toxicol., 7: 613-618.
PubMed - Zhang, X., N. Yamamoto, S. Soramoto and I. Takenaka, 2001. Cisplatin-induced germ cell apoptosis in mouse testes. Arch. Androl., 46: 43-49.
PubMed - Sasaki, K., T. Murakami, M. Kawasaki and M. Takahashi, 1987. The cell cycle associated change of the Ki-67 reactive nuclear antigen expression. J. Cell. Physiol., 133: 579-584.
CrossRefDirect Link - Scholzen, T. and J. Gerdes, 2000. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol., 182: 311-322.
CrossRefDirect Link - Ferrara, D., N. Hallmark, H. Scott, R. Brown, C. McKinnell, I.K. Mahood and R.M. Sharpe, 2006. Acute and long-term effects of in utero exposure of rats to di(n-butyl) phthalate on testicular germ cell development and proliferation. Endocrinology, 147: 5352-5362.
CrossRefDirect Link - Gomez-Lazaro, M., F.J. Fernandez-Gomez and J. Jordan, 2004. p53: Twenty five years understanding the mechanism of genome protection. J. Physiol. Biochem., 60: 287-307.
CrossRefPubMedDirect Link - Boekelheide, K., H.A. Schoenfeld, S.J. Hall, C.C. Weng and G. Shetty et al., 2005. Gonadotropin-releasing hormone antagonist (Cetrorelix) therapy fails to protect nonhuman primates (Macaca arctoides) from radiation-induced spermatogenic failure. J. Androl., 26: 222-234.
CrossRefDirect Link - Nakai, M., R.A. Hess, J. Netsu and T. Nasu, 1995. Deformation of the rat Sertoli cell by oral administration of carbendazim (methyl 2-benzimidazole carbamate). J. Androl., 16: 410-416.
PubMedDirect Link - Narayana, K., U.J.A. D'Souza, P. Narayan and G. Kumar, 2005. The antiviral drug ribavirin reversibly affects the reproductive parameters in the male Wistar rat. Folia Morphologica, 64: 65-71.
PubMedDirect Link - Maines, M.D., P.M. Sluss and M. Iscan, 1989. cis-Platinum-mediated decrease in serum testosterone is associated with depression of luteinizing hormone receptors and cytochrome P-450scc in rat testis. Endocrinology, 126: 2398-2406.
CrossRefDirect Link - Cameron, D.F., K.E. Muffly and S.J. Nazian, 1993. Reduced testosterone during puberty results in a midspermiogenic lesion. Exp. Biol. Med., 202: 457-464.
CrossRefDirect Link - Davies, K.J.A., 1995. Oxidative stress: The paradox of aerobic life. Biochem. Soc. Symp., 61: 1-31.
CrossRefPubMedDirect Link - Kim, Y.K., J.S. Jung, S.H. Lee and Y.W. Kim, 1997. Effects of antioxidants and Ca2+ in cisplatin-induced cell injury in rabbit renal cortical slices. Toxicol. Applied Pharmacol., 146: 261-269.
CrossRefDirect Link - Cao, L., S. Leers-Sucheta and S. Azhar, 2004. Aging alters the functional expression of enzymatic and non-enzymatic anti-oxidant defense systems in testicular rat Leydig cells. J. Steroid Biochem. Mol. Biol., 88: 61-67.
CrossRefDirect Link - Kalender, Y., M. Yel and S. Kalender, 2005. Doxorubicin hepatotoxicity and hepatic free radical metabolism in rats: The effects of vitamin E and catechin. Toxicology, 209: 39-45.
CrossRefDirect Link - Vernet, P., R.J. Aitken and J.R. Drevet, 2004. Antioxidant strategies in the epididymis. Mol. Cell. Endocrinol., 216: 31-39.
CrossRefDirect Link - Lafuente, A. and A.I. Esquifino, 1999. Cadmium effects on hypothalamic activity and pituitary hormone secretion in the male. Toxicol. Lett., 110: 209-218.
CrossRefDirect Link - Silici, S., O. Ekmekcioglu, G. Eraslan and A. Demirtas, 2009. Antioxidative effect of royal jelly in cisplatin-induced testes damage. Urology, 74: 545-551.
CrossRefDirect Link - Cai, L., B.F. Hales and B. Robaire, 1997. Induction of apoptosis in the germ cells of adult male rats after exposure to cyclophosphamide. Biol. Reprod., 56: 1490-1497.
CrossRefPubMedDirect Link - Huddart, R.A., J. Titley, D. Robertson, G.T. Williams, A. Horwich and C.S. Cooper, 1995. Programmed cell death in response to chemotherapeutic agents in human germ cell tumour lines. Eur. J. Cancer, 31: 739-746.
CrossRefDirect Link - Chresta, C.M., J.R. Masters and J.A. Hickman, 1996. Hypersensitivity of human testicular tumors to etoposide-induced apoptosis is associated with functional p53 and a high Bax:Bcl-2 ratio. Cancer Res., 56: 1834-1841.
PubMedDirect Link - Meistrich, M.L., M. Finch, M.F. da Cunha, U. Hacker and W.W. Au, 1982. Damaging effects of fourteen chemotherapeutic drugs on mouse testis cells. Cancer Res., 42: 122-131.
PubMedDirect Link - Laskey, J.W., G.H. Rehnberg, S.C. Laws and J.F. Hein, 1984. Reproductive effects of low acute doses of cadmium chloride in adult male rats. Toxicol. Applied Pharmcol., 73: 250-255.
CrossRefDirect Link - Bailey, T.L., R.S. Hudson, T.A. Powe, M.G. Riddell, D.F. Wolfe and R.L. Carson, 1998. Caliper and ultrasonographic measurements of bovine testicles and a mathematical formula for determining testicular volume and weight in vivo. Theriogenology, 49: 581-594.
CrossRefDirect Link - Yang, H.S., D.K. Han, J.R. Kim and J.C. Sim, 2006. Effects of α-Tocopherol on cadmium-induced toxicity in rat testis and spermatogenesis. J. Korean Med. Sci., 21: 445-451.
CrossRefPubMedDirect Link - Azu, O.O., I.O.D. Francis, A.O. Abraham, C.N. Crescie, O.E. Stephen and O.O. Abayomi, 2010. Protective agent, Kigelia Africana fruit extract, against cisplatin-induced kidney oxidant injury in Sprague-Dawley rats. Asian J. Pharm. Clin. Res., 3: 84-88.
Direct Link - Lirdi, L.C., T. Stumpp, E. Sasso‐Cerri and S.M. Miraglia, 2008. Amifostine protective effect on cisplatin-treated rat testis. Anatomical Record, 291: 797-808.
CrossRefDirect Link - Peng, S.J., R.K. Lu and L.H. Yu, 1997. [Effects of semen cuscutae, Rhizoma curculiginis, Radix morindae officinalis on human spermatozoan's motility and membrane function in vitro]. Chin. J. Integr. Tradit. Western Med., 17: 145-147, (In Chinese).
PubMedDirect Link - Amin, A. and A.A. Hamza, 2006. Effects of Roselle and Ginger on cisplatin-induced reproductive toxicity in rats. Asian J. Androl., 8: 607-612.
CrossRefPubMedDirect Link - Amin, A., D. Mahmoud-Ghoneim, M.I. Syam and S. Daoud, 2012. Neural network assessment of herbal protection against chemotherapeutic-induced reproductive toxicity. Theoret. Biol. Med. Modell., Vol. 9.
CrossRefDirect Link - Ilbey, Y.O., E. Ozbek, M. Cekmen, A. Simsek, A. Otunctemur and A. Somay, 2009. Protective effect of curcumin in cisplatin-induced oxidative injury in rat testis: Mitogen-activated protein kinase and nuclear factor-kappa B signaling pathways. Hum. Reprod., 24: 1717-1725.
CrossRefDirect Link - Yüce, A., A. Ateşşahin, A.O. Çeribaşı and M. Aksakal, 2007. Ellagic acid prevents cisplatin-induced oxidative stress in liver and heart tissue of rats. Basic Clin. Pharmacol. Toxicol., 101: 345-349.
CrossRefPubMedDirect Link - Atasayar, S., H. Gurer-Orhan, H. Orhan, B. Gurel, G. Girgin and H. Ozgunes, 2009. Preventive effect of aminoguanidine compared to vitamin E and C on cisplatin-induced nephrotoxicity in rats. Exp. Toxicol. Pathol., 61: 23-32.
CrossRefDirect Link
Elijah B. Kelvin Reply
Nice one