
K. Busambwa
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
R.L. Miller-Cebert
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
L. Aboagye
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
L. Dalrymple
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
J. Boateng
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
L. Shackelford
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
L.T. Walker
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
M. Verghese
Department of Food and Animal Sciences, Nutrition Biochemistry and Carcinogenesis Laboratory, Alabama A and M University, Normal, Al, 35762, United State of America
International Journal of Cancer Research
Year: 2014 | Volume: 10 | Issue: 1 | Page No.: 27-36







ABSTRACT
Peas (yellow and green) and lentils contain bioactive compounds with protective properties. Sprouting of grains has shown to improve nutritional value due to activation of hydrolytic enzymes. The aim of this study was to determine anticarcinogenic potential of sprouted and non-sprouted green-split and yellow peas and lentils at 5 and 10% on Azoxymethane (AOM)-induced Aberrant Crypt Foci (ACF) in Fisher 344 male rats. Seventy-eight rats were randomly assigned to 13 groups: Control (C) received AIN-93G diet; treatment diets included Lentils sprouted and non-sprouted (LS, LNS), Green peas sprouted and non-sprouted (GS, GNS), Yellow peas sprouted and non-sprouted (YS, YNS) at 5 and 10% levels in an AIN 93 based diet. At 7 and 8 week, rats received 2 subcutaneous injections of AOM at 16 mg kg-1 b.wt. Rats were euthanized at 17 week by CO2 Asphyxiation. Total ACF ranged from 88 to 181.33 in rats fed C, LS and LNS, GS and GNS, YS and YNS. ACF reductions (%) compared to C were 68.56; 63.25 and 51.46 in rats fed LS at 10%, LNS at 10% and YNS at 10%, respectively. Glutathione content, Glutathione-S-transferase and Catalase activities were significantly higher (p<0.05) in rats fed treatment diets compared to C. Findings from this study indicate that sprouted and non sprouted green split and yellow peas and lentils reduced AOM-induced aberrant crypt foci in F344 male rats with the greatest reduction in rats fed Lentil sprouted (10%). Selected legumes appear to have beneficial potential in reducing colon cancer.
PDF Abstract XML References Citation
Received: January 19, 2013;
Accepted: September 07, 2013;
Published: February 28, 2014
How to cite this article
K. Busambwa, R.L. Miller-Cebert, L. Aboagye, L. Dalrymple, J. Boateng, L. Shackelford, L.T. Walker and M. Verghese, 2014. Inhibitory Effect of Lentils, Green Split and Yellow Peas (Sprouted and
Non-sprouted) on Azoxymethane-induced Aberrant Crypt Foci in Fisher 344 Male
Rats. International Journal of Cancer Research, 10: 27-36.
DOI: 10.3923/ijcr.2014.27.36
URL: https://scialert.net/abstract/?doi=ijcr.2014.27.36
DOI: 10.3923/ijcr.2014.27.36
URL: https://scialert.net/abstract/?doi=ijcr.2014.27.36
INTRODUCTION
Cancer results from mutations responsible for inhibiting oncogene and tumor suppressor gene functions which lead to uncontrolled cell growth. This disease remains extremely complex and continues to grow in the industrialized world due to lifestyle factors. In 2007, the disease claimed the lives of about 7.6 million people worldwide (ACS, 2009-2010). According to the American Cancer Society, an estimated about 102,480 new cases of colon cancer to be diagnosed in the United States of America in 2013 and from which about 50,830 deaths are expected (ACS, 2011). Colon cancer forms in the tissues of the colon. Colon cancer is diagnosed with high frequency in the U.S. due to lifestyle choices such as physical inactivity and/or unbalanced diet (NCI, 2008). Studies have shown that because colon cancer is linked to diet, it is the easiest cancer to prevent (Giovannucci, 2002; Giovannucci and Willett, 1994; Liu, 2003; Platz et al., 2000). Diet plays an important part in cancer prevention. Therefore, a balanced diet low in fat and rich in fresh fruits, vegetables and whole grains is highly recommended for increased protection against colon cancer (Rajamanickam and Agarwal, 2008).
Originally from India, lentils are legumes that grow in a pod like peas or beans. Lentils contain dietary fiber, folate, carbohydrate, complex minerals (calcium, magnesium) and vitamins (A, C, E, D and B) (Roy et al., 2010). The essential amino acids, isoleucine and lysine are also found in lentils (Roy et al., 2010). Moreover, lentils are one of the highest sources of a number of phenolic compounds including phenolic acid, flavan-3-ols, flavones and flavonol glycosides, anthocyanins and proanthocyanidins (Xu and Chang, 2010). These phenolic compounds with antioxidant activity may play an important role in protection against human diseases. Peas are the seed-pod of the legume, Pisum Sativum and are among the longest cultivated plants. Common peas are usually used as vegetable, fresh or frozen in Indian cuisine. Peas are also grown to produce dry peas such as split peas and come in two varieties, green and yellow. About 700,000 metric tons of split peas were produced in the US in 2009 (FAOUN, 2010). Peas have a significant amount of fiber and soluble fiber, proteins, complex carbohydrates, folates and iron (Faoun, 2010). Sprouting of seeds has the beneficial effects such as reducing anti-nutrient factors namely tannins, phytates and increasing the bioavailability of micro and macronutrients (Hedges and Lister, 2006), also increasing the digestibility of proteins and carbohydrates (Chavan et al., 1989).
Azoxymethane is an effective and potent carcinogen inducing colon carcinoma (Chen and Huang, 2009). The metabolite products of AOM have the affinity to bind to cellular compounds such as DNA causing DNA mutations which may lead to the formation of preneoplastic lesions, aberrant crypt foci. ACF are histopathological markers for colon cancer. When left untreated, these preneoplastic lesions can evolve into polyps which are precancerous growth in the tissues of colon (Takayama et al., 1998).
The objective of this study was to determine the chemopreventive effects of lentils, green split peas and yellow peas (sprouted and non-sprouted) at 5 and 10% levels on the incidence of AOM-induced colon carcinogenesis in Fisher 344 male rats.
MATERIALS AND METHODS
Animal experiment: Animal housing and diet: Fisher 344 male rats were obtained from Harlan, IN in January 2011 and housed in Animal Laboratory of the Department of Food and Animal Sciences at Alabama A and M University for a period of 13 weeks. They were randomly assigned in pairs in stainless cages (2 rats/cage). The temperature and relative humidity were kept at 20±1°C and 50%, respectively. Light and dark cycles were kept at 12 h each. The rats were allowed 2 week to acclimate to the conditions described above and fed an American Institute of Nutrition diet (AIN-93G)-control (Reeves, 1997) following by the treatment diets composed of sprouted and non-sprouted lentils, yellow and green split peas at 5 and 10% levels for a 13-week period. All diets were prepared according to guidelines of the American Institute of Nutrition (AIN) and stored for no more than a 2 week period. Ingredients were provided by The National Bean Council (CA). Feed and water were provided as needed. Daily feed intakes and weekly body weight were recorded.
AOM injection: All rats received two azoxymethane (AOM) injections (Sigma Chemicals, St. Louis, MO) at 7 and 8 weeks of age. AOM was administered s/c at 16 mg kg-1 body in 0.09% saline solution.
Colon sample collection and counting of aberrant crypt foci: At 17 weeks of age, rats were killed by CO2 asphyxiation. Colons, livers were removed. Enumeration of ACF was performed following protocol described by Bird (1987).
Colons were longitudinally cut open then flushed with Phosphate-buffered Saline (PBS) (0.1 M, pH 7.2) (Fisher Scientific, Suwannee, GA). The split colons were placed on a filter paper then immersed in 10% buffered formalin (Fisher Scientific). The colons were separated into proximal and distal sections and each portion was cut into 2 cm long segment. These segments were stained with 0.5% methylene blue solution (Sigma Chemicals, St. Louis, Mo). The segments of colon were placed on a slide for microscopic examination and the total number of ACF and crypts were counted.
Preparation of liver samples for enzymatic activities: Approximately 5 g of liver sample was homogenized in 50 mL of potassium phosphate buffer (0.1 m, pH 7.2) and centrifuged at 15,0000xg for 20 min. Supernatant was collected for catalase, superoxide dismutase and Glutathione s-transferase assays. For Glutathione assay, the supernatant was deproteinized then stored at -20°C.
Catalase assay: Catalase activity in rat liver was assayed following the methods described by Johansson and Borg (1988). The method is based on the reaction of the enzyme with methanol in the presence of H2O2. One unit of CAT was defined as the amount of enzyme needed to cause the formation of 1 nmol of formaldehyde min-1. The absorbance was recorded using microplate reader (synergy HT) at 540 nm.
Superoxide dismutase (SOD) assay: SOD activity was carried out spectrophotometrically according to methods described by Folhe and Otting (1984). Clear supernatant was mixed with tetrazolium salt for detection of superoxide radicals generated by xanthine oxidase. One unit of SOD was expressed as the amount of enzyme required to exhibit 50% dismutation of the superoxide radical. The absorbance was measured at 440-460 nm using a microplate reader (Synergy HT).
Glutathione-s-transferase assay: GST activity in animal liver was assayed according to protocol as described by Habig et al. (1974). Liver supernatant was mixed with 1-chloro2, 4-dinitrobenzene (CDNB) (Fisher Scientific, Suwanne, GA). GST was determined at 340 nm and expressed as μmol mg-1.
Glutathione assay: GSH serves as a nucleophilic substrate in the detoxification of xenobiotics. GSH content was determined following methods as described by Griffith (1980). Using 5, 5’-dithio-bis-(2-nitrobenzoic acid) as Ellman’s reagent (Fisher Scientific, Suwannee, GA), GSH was determined at 405-414 nm and expressed as μM.
Statistical analysis: Data were analyzed using the Statistical Analysis System (SAS) version 9.2 and ANOVA. The values are given as Mean±SEM and Means are separated using Tukey’studentized test. Levels of significance are tested at p<0.05.
RESULTS
Weight gain and daily feed intake in 344 fisher rats: Significant (p<0.05) differences in average weight gain (g week-1) were observed among rats fed control, 5 and 10% LS, 5% LNS and rats fed 10% LNS, 5 and 10% GS, GNS, YS and 5% YNS (Table 1). There were, however no significant differences in weight gain among rats fed control, 5 and 10% LS, 5 and 10% LNS (32.21, 31.76, 32.60, 31.43, 32.03, respectively). Feed intake (g) was significantly (p<0.05) different among rats fed 10% GNS and 5% YNS (19.13 and 22.73). There were no significant differences in feed intake between rats fed 5 and 10% GS, 5% GNS, 5 and 10% YS and 10% YNS.
Number of ACF incidence in proximal and distal colon of rats: Higher numbers of ACF were primarily observed in the distal colon of rats compared to the proximal section of the colon of rats (Table 2). Aberrant crypt foci incidence in distal colon of control fed rats was significantly higher (p<0.05) than in the experimental groups (Table 2).
| Table 1: | Effect of diets on weight gain and feed intake in Fisher 344 male rats |
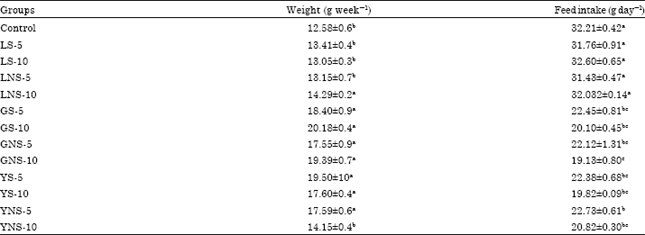 | |
| abcMeans in column with the same superscript do not significantly differ (p<0.05). Values are Means±SE. LS: Lentil sprouted, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted | |
| Table 2: | Effect of diets on total No. of AOM-induced ACF incidence in colon of Fisher 344 male rats |
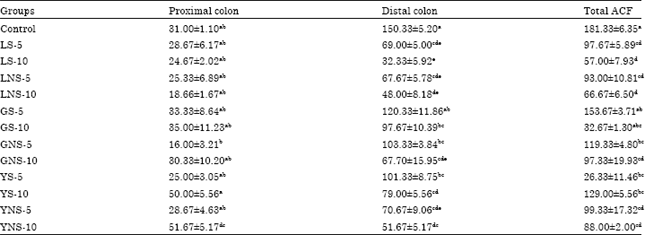 | |
| abcMeans in column with the same superscript do not significantly differ (p<0.05). Values are Means±SE. LS: Lentil sprouted, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted | |
Among the experimental groups, there were significant (p<0.05) differences in the number of ACF in both the proximal and distal colon. A higher total number of ACF were observed in the rats fed GS at 5% (153.67) compared to other treated groups. Percent reductions of ACF ranged from a low of 15.47% in rats fed GS-5 (153.67) to a high of 68.51% in rats fed LS-10 (57.00) in rats fed treatment diets compared to the control group (Table 2).
Crypt multiplicity: There were significant differences (Fig. 1) in large ACF with 4 aberrant crypts per focus among treatment and control groups. A low number of aberrant crypt foci with 3 crypts per focus were seen in the rats fed LS at 10%. ACF with 3, 4 and 5 or more crypts per focus were significantly higher (p<0.05) in control compared to the treatment groups. The size of the ACF is defined as crypt multiplicity. ACF with more than 5 crypts has a higher probability of progression to tumors. Colons of treatment rats did not exhibit significant differences in their number of crypts (mostly ACF with 1-2 crypts). The rats fed GS at 5 and 10% and YS at 5 and 10% had high number of ACF with 1-2 crypts compared to other treatment groups.
Catalase assay: Catalase is an antioxidant enzyme involved in the detoxification of hydrogen peroxide, a reactive oxygen species which is a toxic product in aerobic metabolism. Catalase activity (nmol min mL-1) was significantly (p<0.05) lower (82.80) in control compared to treatment groups (Table 3). Among the treatment groups, catalase activity in rats fed LS at 5 and 10% (358.98, 360.53) was higher than in rats fed GS, GNS, YS and YNS (Table 3). The lowest catalase activity among the treatment groups was seen in rats fed GS at 10%.
Superoxide dismutase activity: Superoxide dismutase is an enzyme that catalyzes the dismutation of the superoxide anion to molecular oxygen and hydrogen peroxide. SOD is involved as a crucial cellular antioxidant. SOD activity (U mL-1) was significantly (p<0.05) higher in rats fed control (289.20) (Table 3) compared to rats fed LS, LNS, GS, YS and YNS at 5% compared to rats fed LS, LNS, GS, YS and YNS at 10% (265.92, 272, 242.73, 105.92, 259.52 and 198.32, 166.40, 25.11, 80.57, 236.93, respectively) (Table 3).
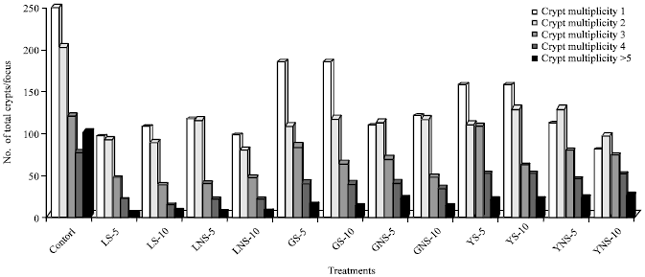 | |
| Fig. 1: | Effect of diet on No. of crypts per focus in isher 344 male rats (n = 78). LS: Sprouted lentil, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted |
| Table 3: | Glutathione-s-transferase, Superoxide dismutase, Catalase activities and GSH content in fisher 344 male rats |
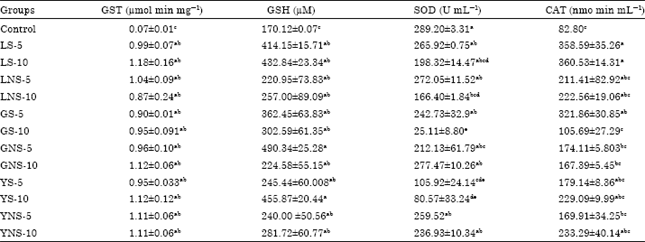 | |
| abcedMeans in column with the same superscript do not significantly differ (p<0.05). Values are means±SE. LS: Lentil sprouted, LNS: Lentil non-sprouted, GS: Green split peas sprouted, GNS: Green split peas non sprouted, YS: Yellow peas sprouted, YNS: Yellow peas non-sprouted | |
SOD activity was significantly (p<0.05) lower in rats fed GS at 10% (25.11) and fed YS at 10% (80.57) than in rats fed GS at 10% (242.76) and fed YS at 5% (105.92).
Glutathione levels (μM): Total GSH content (μM) in the liver of rats fed control (170.12±0.066) was significantly (p<0.05) lower compared to treatment groups (Table 3). Higher GSH content was observed in rats fed GNS at 5% (490.34) and in rats fed YS at 10% (455.87) compared to the other treatments. Higher GSH content was seen in rats fed LS at 10%, LNS-10%, GS at 5%, GNS at 5%, YS at 10%, YNS at 10% (432.84, 257, 362.45, 490.34, 455.87, 281.72, respectively) compared to rats fed LS at 5%, LNS at 5%, GS at 10%, GNS at 10%, YS at 5% and YNS at 5% (414.15, 220.95, 302.59, 224.58, 245.44, 240, respectively).
Glutathione-S-transferase (GST) activity: Total GST activity (μmol min mg-1) was significantly (p<0.05) different between rats fed control and treatment groups (0.7 and 0.99, 1.18, 1.04, 0.87, 0.90, 0.95, 0.96, 1.12, 0.95, 1.12, 1.11, 1.11, respectively) (Table 3). Higher GST activity, numerically, was observed in rats fed LS at 10% (1.18) compared to other treatments. Lowest GST activity was seen in rats fed LNS-10% (0.87).
DISCUSSION
The purpose of the study was to investigate the inhibitory effect of lentils, green split pea and yellow peas (sprouted and non-sprouted) at 5 and 10% (LS-5, LNS-10, LNS-5, LNS-10, GS-5, GS-10, GNS-5, GNS-10, YS-5, YS-10, YNS-5, YNS-10), respectively on the incidence of Azoxymethane-induced ACF in Fisher 344 male rats and their effect on selected antioxidant enzymes activities (Catalase, CAT; Superoxide dismutase, SOD) and phase II enzyme (Glutathione-s-transferase, GST).
Results from this study showed that AOM-induced ACF were significantly lower in rats fed lentils, green split and yellow peas (sprouted and non-sprouted) compared to rats fed the control diet. Also, glutathione levels, catalase and glutathione-s-transferase activities were significantly enhanced in treatment diet fed groups compared to control. Meanwhile, SOD activity was significantly higher in control compared to the treatment groups.
The various bioactive components of legumes, such as fiber, vitamin E and phytochemicals have been showed to possess effective antioxidant properties with anticarcinogenic potential (Bolton et al., 1981). Sprouting of seeds may have positive results such as production of short chain fatty acids, enhancement of bioavailability of both macro and micronutrients such as minerals, vitamins (folate, vitamins B and C) and phytochemicals (Hedges and Lister, 2006). However, the sprouting process and seed type have an impact on phytochemical levels (Hedges and Lister, 2006). These phenolic compounds have been shown to contribute to the overall antioxidant activity with the capability of exerting protective effect against certain chronic diseases (Fernandes et al., 2010). Rats fed the control diet had significantly (p<0.05) higher feed intake but lower weight gain compared to the treatment groups. This may be due to preneoplastic lesions present in colon preventing the absorption of nutrients. In this regard, ACF was significantly higher in rats fed the control diet compared to those fed the treatment diets. Previous research (Boateng et al., 2009) has shown that the reduction of ACF incidence in rats fed a diet rich in fiber mainly fermentable fiber may be associated with decreased interaction of tumor promoters in mucosa by reducing transit time of colon content by enhancing beneficial intestinal bacteria flora due to fermentation of fiber resulting in the lowering of colonic pH thus inhibit the process of cancer formation (Kanda et al., 2012). Among the treatment groups, those that were fed lentils sprouted and non-spouted had significantly lower ACF incidence in both proximal and distal portion of the colon. This has been observed in several studies, Faris et al. (2009) showed that in addition to soluble and insoluble fiber, lentils are good source of folate, important element in DNA repair via methylation. Also, lentils contain several phenolic compounds such as catechin and non phenolic compounds that may exert a synergistic anticarcinogenic effect (Faris et al., 2009). The sight difference in ACF inhibitory effect between lentils, green split and yellow peas, sprouted and non-sprouted may be attributed to the difference in polyphenolics compounds present mainly in the seed coats of the whole seed.
According to Shirtliff and Bird (1996), ACF number and crypt multiplicity are important in the prediction of the pathogenesis of colon cancer. ACF with 1-2 crypts per foci are more sensitive to elimination but this is not the case for ACF with 3-4 crypts (Shirtliff and Bird, 1996). However, ACF with increasing crypts multiplicity are differently regulated by carcinogenesis modulating agents (Bird, 1995) These ACF acquire complete growth autonomy thus increasing tumor outcome. This study demonstrates that feeding a diet consisting of lentils, green split and yellow peas, sprouted and non-sprouted significantly reduced ACF that contained <5 crypts per foci but had lesser effects on those with crypt multiplicity of 1-2 and 3-4. Foci with 3 or more crypts were higher in rats fed control compared to rats fed treatment diets. The reduction in formation of ACF with higher crypt multiplicity in rats fed treatment diets (LS, LNS, GS, GNS, YS, YNS) when compared with the control diet, indicates that a diet consisting of legumes may have a protective effect against carcinogenesis. Sprouted and non-sprouted legumes have been reported to contain a range of minerals such as zinc, magnesium, calcium; antioxidant compounds such as phenolics, flavonoids and catechins; B vitamins (Folates) (Hedges and Lister, 2006). These suppressing agents have been found to exert their anti-carcinogenic effect by several mechanisms, including induction of apoptosis, activation of liver detoxification enzymes and suppression of altered gene expression by DNA methylation (Faris et al., 2009).
As chemopreventive agents, phytochemicals have the ability to induce phase I and phase II enzyme metabolism of carcinogens. According to Boateng et al. (2007), diet that contains a considerable amount of cysteine, may increase the activity of hepatic GST, SOD, CAT activities and GSH content in liver of rats. These detoxification enzymes play a role in the defense system of the body by preventing the accumulation of reactive oxygen species which may lead to DNA damage therefore resulting in carcinogenesis (Pham et al., 2009). In this study, we saw a significant increase in hepatic and detoxification enzyme activities (GST, CAT, GSH) in rats fed selected legumes compared to rats fed the control diet. Glutathione s-Transferase activity in rats fed selected legumes was significantly higher compared to the rats fed the control diet. The increase in the hepatic GST levels was concurrent with significant reduction of total ACF numbers only for LS-5, LNS-5, GNS-10, YNS-5 and YNS-10 diet groups. The increased GSH content of rats fed treatment diets (LS-5, LS-10, GS-5, YS-10, YNS-10) mirror the CAT activity in rats fed the same treatment diets. CAT is involved in the removal of hydrogen peroxide resulted from dismutation of superoxide anion by the action of SOD. We however observed an increased SOD activity in rats fed the control diet compared to rats fed treatment diets. This contradicted the findings reported by Kanda et al. (2012) and Casaril et al. (1994). They reported a decreased or unchanged SOD expression in cancer cells. Our findings correspond with findings reported by Pham et al. (2009) who reported an elevated level of SOD in cancer cell lines compared with normal tissues. According to Satomi et al. (1995), SOD activity in cancerous cells will increase with the progression of cancer and with the evolving invasion. These discrepancies may be explained by the different stages of tumors at the time of study (Skrzycki et al., 2009). The action of SOD balances the formation and elimination of ROS to maintain proper cellular status. SOD catalyzes the reduction of superoxide to hydrogen peroxide which is then decomposed by the action of catalase.
CONCLUSION
This research has shown that lentils, green split and yellow peas sprouted and non-sprouted have a chemopreventive effect against early carcinogenesis by reducing the incidence in AOM-induced ACF and by enhancing phase I and phase II detoxification enzyme activity in fisher 344 male rats. Diet rich in legumes may contribute to the improvement of a healthy life style and lentils, green split and yellow peas (sprouted and non-sprouted) contain essential nutrients and phytochemicals with chemopreventive potential necessary for the amelioration of health. These preliminary results must be followed by further long-term studies (animal end-point tumor model) and clinical trials for more conclusive evidence.
ACKNOWLEDGEMENT
The lentils, green split and yellow peas were courtesy of Inland Empire Foods, Inc., 5425 Wilson Street, Riverside CA 92509; and the Alabama Agricultural Experimental Research Station, Normal Al 35762.
REFERENCES
- Bird, R.P., 1987. Observation and quantification of aberrant crypts in the murine colon treated with a colon carcinogen: Preliminary findings. Cancer Lett., 37: 147-151.
PubMed - Bird, R.P., 1995. Role of aberrant crypt foci in understanding the pathogenesis of colon cancer. Cancer Lett., 93: 55-71.
CrossRefPubMedDirect Link - Boateng, J.A., M. Verghese, L.T. Walker, L.A. Shackelford and C.B. Chawan, 2007. Inhibitory effects of selected dry beans (Phaseolus spp L) on azoxymethane-induced formation of aberrant crypt foci in Fisher 344 male rats. Nutr. Res., 27: 640-646.
CrossRefDirect Link - Boateng, J., M. Verghese, V. Panala, L.T. Walker and L. Shackelford, 2009. Protective effects of rice bran on chemically induced colon tumorigenesis may be due to synergistic/additive properties of bioactive components. Int. J. Cancer Res., 5: 153-166.
CrossRefDirect Link - Bolton, R.P., K.W. Heaton and L.F. Burroughs, 1981. The role of dietary fiber in satiety, glucose and insulin: Studies with fruit and fruit juice. Am. J. Clin. Nutr., 34: 211-217.
Direct Link - Casaril, M., F. Corso, A. Bassi, F. Capra and G.B. Gabrielli et al., 1994. Decreased activity of scavenger enzymes in human hepatocellular carcinoma, but not in liver metastases. Int. J. Clin. Lab. Res., 24: 94-97.
CrossRef - Chavan, J.K., S.S. Kadam and L.R. Beuchat, 1989. Nutritional improvement of cereals by sprouting. Crit. Rev. Food Sci. Nutr., 28: 401-437.
CrossRefPubMedDirect Link - Faris, M.A.I.E., H.R. Takruri, M.S. Shomaf and Y.K. Bustanji, 2009. Chemopreventive effect of raw and cooked lentils (Lens culinaris L.) and soybeans (Glycine max) against azoxymethane-induced aberrant crypt foci. Nutr. Res., 29: 355-362.
CrossRefDirect Link - Fernandes, A.C., W. Nishida and R.P. Da Costa Proenca, 2010. Influence of soaking on the nutritional quality of common beans (Phaseolus vulgaris L.) cooked with or without the soaking water: A review. Int. J. Food Sci. Technol., 45: 2209-2218.
CrossRef - Folhe, L. and F. Otting, 1984. Superoxide dismutase assays. Methods Enzymol., 105: 93-104.
PubMedDirect Link - Giovannucci, E., 2002. Modifiable risk factors for colon cancer. Gastroenterol. Clin. North Am., 31: 925-943.
CrossRef - Giovannucci, E. and W.C. Willett, 1994. Dietary factors and risk of colon cancer. Ann. Med., 26: 443-452.
PubMed - Griffith, O.W., 1980. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem., 106: 207-212.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Johansson, L.H. and L.A. Borg, 1988. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem., 174: 331-336.
CrossRefPubMedDirect Link - Kanda, B., J. Boateng, L. Shackelford, S. Appiah, K. Campbell L.T. Walker and M. Verghese, 2012. Effects of processed peaches (Prunus persica) in reducing azoxymethane (AOM) induced Aberrant Crypt Foci (ACF) in fisher 344 male rats. Int. J. Cancer Res., 8: 1-14.
CrossRef - Liu, R.H., 2003. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr., 78: 517S-520S.
CrossRefPubMedDirect Link - Pham, T.M., Y. Fujino, M. Ando, K. Suzuki and K. Nakachi et al., 2009. Relationship between serum levels of superoxide dismutase activity and subsequent risk of lung cancer mortality: Findings from a nested case-control study within the Japan Collaborative Cohort Study. Asian Pac. J. Cancer Prev., 10: 75-79.
PubMed - Platz, E.A., W.C. Willett, G.A. Colditz, E.B. Rimm, D. Spiegelman and E. Giovannucci, 2000. Proportion of colon cancer risk that might be preventable in a cohort of middle-aged US men. Cancer Causes Control, 11: 579-588.
PubMed - Reeves, P.G., 1997. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr., 127: 838S-841S.
PubMedDirect Link - Roy, F., J.I. Boye and B.K. Simpson, 2010. Bioactive proteins and peptides in pulse crops: Pea, chickpea and lentil. Food Res. Int., 43: 432-442.
CrossRefDirect Link - Satomi, A., S. Murakami, T. Hashimoto, K. Ishida, M. Matsuki and M. Sonoda, 1995. Significance of superoxide dismutase (SOD) in human colorectal cancer tissue: Correlation with malignant intensity. J. Gastroenterol., 30: 177-182.
CrossRef - Shirtliff, N. and R.P. Bird, 1996. Growth features of aberrant crypt foci that resist modulation by cholic acid. Carcinogenesis, 17: 2093-2096.
PubMed - Skrzycki, M., M. Majewska, M. Podsiad and H. Czeczot, 2009. Expression and activity of superoxide dismutase isoenzymes in colorectal cancer. Acta Biochim. Polt., 56: 663-670.
PubMed - Takayama, T., S. Katsuki, Y. Takahashi, M. Ohi and S. Nojiri et al., 1998. Aberrant crypt foci of the colon as precursors of adenoma and cancer. N. Engl. J. Med., 339: 1277-1284.
Direct Link - Xu, B. and S.K.C. Chang, 2010. Phenolic substance characterization and chemical and cell-based antioxidant activities of 11 lentils grown in the Northern United States. J. Agric. Food Chem., 58: 1509-1517.
CrossRef