
B. Kanda
Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
J. Boateng
Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
L. Shackelford
Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
S. Appiah
Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
K. Campbell L. T. Walker
Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
M. Verghese
Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
International Journal of Cancer Research
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 1-14







ABSTRACT
Peach is an important fruit consumed worldwide. This study was conducted to investigate the effects of processed peaches (dried, frozen and juice) on Azoxymethane-induced (AOM) Aberrant Crypt Foci (ACF) in Fisher 344 male rats, their effect on selected hepatic enzyme activities and to determine the effect of processing (Blanching+Freezing, Blanching+Freeze drying) on total phenolic content and antioxidant potential (DPPH and FRAP). Rats were fed AIN93G as control (CON), treatment diets containing 2 and 4% peaches, and pasteurized juice (PJ) (2 and 4%). Rats received AOM injections s/c at 16 mg kg-1 b wt. during the 7th and 8th wk of age and killed by CO2 asphyxiation at 17 weeks of age. ACF incidence was higher in rats fed CON compared to treatment diets. Rats fed PJ had the lowest ACF (100 and 59 for 2 and 4%) compared to the treatment groups. Blanching (STB+BWB) influenced the incidence of ACF with STB having lower ACF numbers compared to BWB. GST, SOD and CAT (mmol/mg) activities, (9.56-1.43, 1.98-0.49 and 0.53-0.33), respectively were higher in rats fed treatment diets, compared to CON. Total phenolics, ranged from 98.09-120.38, 111.05-120.19, 104.76-122.09 (mg/GAE/g fwb) in FU, FRB and FDB peaches, respectively. FRAP (μg g-1) in FU, FRB and FDB peaches ranged from 0.028-0.042 while (%) DPPH (T30) ranged from 21.588-73.449. Results indicate processing of fruits may impact its role in chemoprevention, total phenolics and antioxidant activity, since fruits are highly perishable; optimum technique is required to minimize losses during and after processing.
PDF Abstract XML References Citation
Received: October 22, 2011;
Accepted: October 28, 2011;
Published: February 13, 2012
How to cite this article
B. Kanda, J. Boateng, L. Shackelford, S. Appiah, K. Campbell L. T. Walker and M. Verghese, 2012. Effects of Processed Peaches (Prunus persica) in Reducing Azoxymethane (AOM) Induced Aberrant Crypt Foci (ACF) in Fisher 344 Male Rats. International Journal of Cancer Research, 8: 1-14.
DOI: 10.3923/ijcr.2012.1.14
URL: https://scialert.net/abstract/?doi=ijcr.2012.1.14
DOI: 10.3923/ijcr.2012.1.14
URL: https://scialert.net/abstract/?doi=ijcr.2012.1.14
INTRODUCTION
Excluding skin cancer, colorectal cancer has been found to be the 3rd most common cancer diagnosed in both men and women in the United States. According to the American Cancer Society (ACS, 2009), colorectal cancer cases in the U.S include 101, 100 new cases, with 52,010 found in men and 54,010 found in women. The overall lifetime risk for developing colorectal cancer is about 1 in 19 hence and this risk being slightly higher in males than in females. Some of the risk factors associated with the development of colon cancer include age, gender, lifestyle, genetics and diet (Boateng et al., 2006).
Diets rich in fruits and vegetables may offer protection against chronic diseases such as cancer (colon) and heart diseases (Rimm et al., 1996; Steinmetz and Potter, 1996; Van't Veer et al., 2000). Vegetables that are common in human diets have been reported to have antitumorigenic effects (Bingham, 1990; Bresnick et al., 1990; Wattenberg and Coccia, 1991). A highly inverse association between intake of total fruits and vegetables and certain cancers was reported by Temple and Gladwin (2003). These protective effects are attributed to the presence of various functional components such as phenolic compounds, vitamin C and E, provitamins, minerals and fiber. At present, there is overwhelming evidence suggesting that these compounds have bioactive components which effectively scavenge Reactive Oxygen Species (ROS) that may cause oxidative damage to lipids, proteins, nucleic acids and reduce cell proliferation in cancer cell lines (Kampa et al., 2000; Meyer et al., 2005).
Azoxymethane (AOM) (CH3-[O] N = N-CH3), is the proximate metabolite of 1, 2-Dimethylhydrazine (DMH) (CH3-NH-NH-CH3), used for induction of ACFs in rodents (Jackson et al., 1999; Jackson et al., 2003). It is reported that AOM is a very potent colon specific carcinogen. In addition, it is regarded as one of the most popular chemical carcinogens used to investigate the effects of diet modification on colorectal carcinogenesis in rats in chemopreventive studies (Sohn et al., 2001; Takahashi et al., 1991; Boateng et al., 2006; Nosho et al., 2006).
ACFs have been described as microscopic precancerous lesions found in the colonic mucosa of rodents treated with a chemical carcinogen such as Azoxymethane. They have been characterized as multiple or single crypts, having altered luminal openings, exhibiting thickened epithelia (Bird, 1995).
Peach (Prunus persica) belongs to the Rosaceae (Rose) family and is related to stone fruits such as plums, cherries and almonds. The peach is a native of China where it signifies long life. It can be consumed either fresh or canned and its flavor can be incorporated into many beverages (Rieger, 2006). Even though peaches have a lower total antioxidant capacity compared to other fruits such as strawberry, apple or orange (Huber and Rupasinghe, 2009), they are nutritionally important because they are one of the most important commodities consumed worldwide. Polyphenols are the main sources of antioxidants in peaches, although Vitamin C and carotenoids also contribute to it (Gil et al., 2002).
Fruits and vegetables are considered to be the major contributors of ROS-scavenging antioxidants. However, various factors such as post harvest storage may have an impact on the nutrient quality and degrade phenolic constituents and other antioxidants. Recently, even a mild thermal processing technique such as blanching has raised concern regarding its impact on functional components, yet thermal processing is an important method of preserving food and maintaining functional attributes such as increasing shelf life, texture, flavor and color (Fellows, 2000).
Based on the potential effects of processing on fruits (peach) and their potential benefit against chronic diseases, the objective of our study was to investigate the effects of processing on antioxidant capacity in peaches (dried, frozen and juice) on Azoxymethane (AOM) induced aberrant crypt foci (ACF) in Fisher 344 male rats and their effect on selected enzyme activities.
MATERIALS AND METHODS
Experimental design and animal housing/ diets: Forty Fisher 344 male weanling rats of 3-4 weeks old obtained from Harlan, (IN) were randomly assigned to 10 groups (4 rats each) following a 1-week acclimatization period.
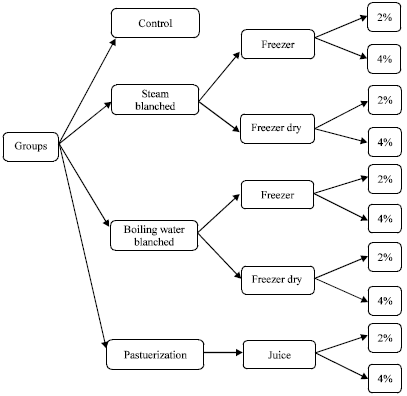 | |
| Fig. 1: | Experimental design. Duration of study =13 week, Number of Groups = 9, Control (AIN 93 G) = 1, Treatment Groups = 10, Total No. of rats N = 44, Rats per group n = 4 |
The groups were assigned to the following diets; Control (CON) were assigned AIN-93G (Reeves et al., 1993), groups receiving peaches were assigned: AIN-93G+2 and 4% steam blanched freeze dry (STBFD), AIN-93G 2 and 4% peach water blanched freeze dry (WTBFD), AIN-93G 2 and 4% peach steam blanched freezer (STBFR), AIN-93G 2 and 4% peach water blanched freezer (WTBFR), respectively. The groups receiving fruit juice were given 2 and 4% Peach Juice (PJ) instead of water. Diet modifications were made based on nutrient composition of peach and juice to maintain isocaloric nature. Rats were fed diets for 13 weeks. Light and dark cycles were maintained at 12 h cycle and the temperature and relative humidity were maintained at 21±1°C and 50%, respectively. Rats were housed in stainless steel cages (2 rats/cage). Feed and water were provided ad-libitum. Rats fed pasteurized peach juices (2 and 4%) were given fresh juice (100 mL) daily. Biweekly body weights and weekly feed intakes were recorded. The diets were prepared fresh weekly and stored at 4°C. Ingredients for making AIN-93 diets were obtained from MP Biomedical (CA). The Institutional Animal Care and Use Committee of Alabama A and M University approved all protocols involving rats (June, 2009) (Fig. 1).
Sample preparation: Peaches were obtained from a local farmers’ market (Huntsville, Al). The fruits were sorted, cleaned and sliced into pieces. Peaches were blanched using a steamer steam (80°C for 1 min) and water blanched (90°C for 30 sec), then sealed in plastic bags, covered with aluminum foil and stored in a conventional home freezer (-20°C) for 3 days. A vertical freezer (Virtis Consol 24, Gardiner, NY) was used for freeze-drying the blanched samples. The dried samples were powdered using a food processor (Robot coupe, Blixer RSI, BS3). Samples were placed into airtight amber containers and stored at -20°C. To prepare peach juice, fruits were sorted; cleaned, deseeded, sliced and blended in a food processor. The homogenate was filtered using cheesecloth and the juice was pasteurized (HTST 72°C for 10 sec) prior to bottling. Concentrated fruit juices were appropriately diluted daily (20 mL/100 mL). Undiluted juice was stored at 4°C.
AOM injection, asphyxiation and tissue sample collection: At 7 and 8 weeks of age, all rats received injections of Azoxymethane (AOM) subcutaneously (NCI Chemical Repository, Kansas City, MO) in saline at 16 mg kg-1 body weight to facilitate ACF induction. At the end of 17 weeks, the rats were fasted overnight and euthanized by CO2 asphyxiation. Following asphyxiation, the liver, cecum and colons were collected from each rat and washed in Phosphate Buffered Saline (PBS) (0.1 M, pH 7.2). Cecal pH, cecal and liver weights were recorded. Livers were subsequently immersed in liquid nitrogen and stored at -80°C until further analysis. Colons from each group were rinsed with PBS (0.1 M, pH 7.2), split longitudinally and fixed flat on a coded strip of filter paper in 10% buffered formalin (Fisher Scientific, Suwannee, GA).
Enumeration of Aberrant Crypt Foci (ACF): Quantification of aberrant crypt foci (ACF) was determined as described by Bird (1987). The colons were sectioned into proximal and distal and each half cut into 2 cm long segments. Segments were stained with 0.2% methylene blue staining solution (Sigma chemicals, St. Louis, MO) and ACF was scored.
Glutathione S-transferase (GST) activity: Glutathione S-transferase (a critical phase II detoxification enzyme) activity in the liver of rats was assayed according to the protocol outlined by Habig et al. (1974). Using 1-chloro 2, 4-dinitrobenzene (Fisher Scientific, Suwannee, GA), as substrate. GST was determined at 340 nm and expressed as μmol mg protein.
Hepatic Superoxide-Dismutase (SOD) activity: Hepatic SOD was determined according to a technique described by Fridivich (1989) with modifications. Xanthine oxidase was used as a substrate. The change in absorbance at 440 nm was monitored every 30 sec for 150 sec. A single unit of enzyme was defined as the quantity of SOD required to produce 50% inhibition of autoxidation.
Catalase (Antioxidative enzyme) activity: Catalase is an intracellular antioxidant enzyme, which protects against oxidative damage occurring as a result of harmful by-products of hydrogen peroxide produced during normal metabolism. Hepatic Catalase activity was determined according to the procedure described by Johansson and Borg (1988) with some modifications. The method is based on the reaction of enzyme with methanol in the presence of an optimal concentration of H2O2. The formaldehyde produced is measured colorimetrically with purpald as the chromogen. Catalase activity was determined against a formaldehyde standard, at an absorbance of 540 nm using a Microplate Elisa reader (Synergy HT) and expressed as micromoles of H2O2 decomposed per mg protein (μmol/mg).
Determination of protein in liver samples: Protein content of samples was determined using Pierce BCA protein assay kit (Sigma Chemical, St Louis, MO).
Chemical analysis of peach extracts
Fruit preparation and extraction: Peaches were sorted, cleaned, cored and lyophilized. The lyophilized peaches were ground into a powder using a commercial food processor (Robot coupe, Blixer RSI, BS3) and the extraction of phenolic compounds was performed as described by Kim et al. (2003). A mixture consisting of 10 g of the lyophilized samples and 100 mL of 80% methanol was homogenized for 1 h and filtered using Whatman No. 2 filter paper. Solvent was removed using a rotary evaporator (Buchi Rotavapor 2025, Gardner Denver Thomas Inc and Niles, IL) at 40°C. Remaining phenolic concentrate was dissolved in 50 mL of 100% methanol and diluted to a final volume of 100 mL using deionized distilled water (ddH20). The phenolic extracts were stored at -20°C until further analysis (Dewanto et al., 2002).
Determination of total phenolics in processed peaches: Total phenolic concentration was determined using a modification of the Folin-Ciocalteu’s method. Briefly, 12.5 μL of sample appropriately diluted was added to 50 μL of ddH2O. Twelve and a half microliters of Folin-Ciocalteu’s phenol reagent was added to the mixture. After 5 min, 7% NaCO3 solution (125 μL) was added to the mixture. Prior to spectrometric analysis, the samples were incubated for 90 min at 25°C. The absorbance of the sample was measured at 750 nm versus a reagent blank using the Microplate reader. A standard curve for total phenolics was developed using Gallic acid standard solution (2:10-10:0). Total phenolic concentration was expressed in mg Gallic Acid Equivalents (GAE) per 100 g of fruit. A replicate of three samples was prepared for analysis.
Free-radical-scavenging activity in processed peaches: Phytochemicals present in fruits and vegetables have been reported to counteract the oxidative effects of free radicals due to their ability to act as free-radical scavengers. The free-radical scavenging activity of peaches was measured as described by Brand-Williams et al. (1995) with slight modifications. The extracts were appropriately diluted to various concentration ratios and 80 μL of each sample volume was added to 220 μL of 0.1 M DPPH (1, 1-Diphenyl-2-picryl-hydrazyl) radical solution. The DPPH radical was dissolved in 80% methanol. The mixture was agitated and incubated at room temperature for 30 min under restricted light before analysis using a Microplate reader at wavelength of 517 nm. Free radical scavenging activity was measured as the amount of extract required to decrease the initial absorbance of DPPH radical concentration by 50% (IC50) as compared to the control according to the equation below.
%DPPH = Acontrol - Asample/Acontrol x100 |
Determination of ferric reducing antioxidant power (FRAP) in processed peaches: The FRAP assay was conducted according to Benzie and Strain (1999) with some modifications. Ten microliters of appropriately diluted sample was combined with 290 μL of FRAP reagent into a 96-well plate. The FRAP reagent was prepared by combining 300 mM acetate buffer (pH 3.6) with 10 mM 2,4,6-tri (2-pyridyl)-s-triazine(TPTZ) in 40 mM HCL and 20 mM ferric chloride (FeCL3.H20) at 10:1:1 ratios. The mixture was subsequently incubated for 10 min at 37°C. Following incubation, the samples were analyzed at an absorbance of 593 nm using a (Synergy HT) Microplate reader (Biotek Instruments INC, Vermont, USA). The change in absorbance (ΔA = Δ4 min-Δ0 min) was compared to ferrous sulfate (FeSO4 7H20) standards (0.1 mM-1.0 mM) at 1 min intervals for 4 min. The samples were analyzed in three replicates and the concentration of Fe3+ was expressed as μmol of Fe2+ g. The FRAP reagent was freshly prepared as needed.
Statistical analysis: Data were analyzed using the SAS statistical program (SAS, 2006). Results were performed using one-way ANOVA. Values are given as Means±SE and means were separated using Tukey’s studentized range test. Differences between treatment groups were tested by student’s t test and paired t test. Unless otherwise indicated levels of significance were tested at p<0.05.
RESULTS
Effects of diets on feed intake and weight gain: Although weight gain did not significantly (p<0.05) differ among the treatment groups (Table 1), rats fed treatment diet had a significantly (P<0.05) higher weight gain compared to the control (Table 1). Among rats fed treatment diets, 2% STB FD and 4% WTB FR had the highest weight gain (224±2.17 and 232±6.73, respectively). While there were no significant (p<0.05) differences in weight gain among the rats fed treatment diets, daily feed intake (g/day) was significantly (p<0.05) different (19.93±0.57 to 19.05±0.39) among treatment groups compared to control (12.73±0.46). Daily feed intake was significantly (p<0.05) higher in the group fed 2% STB FD and 2% STB FR (19.93±0.56 and 19.76±0.51, respectively), compared to control. Even though there was no significant (p<0.05) difference in weight gain between the groups fed 2 and 4% WTB FR (216± 8.51 and 232±6.73, respectively) and 2% juice (200±10.61), daily feed consumption significantly (p<0.05) differed between these groups.
Cecal weight and cecal pH in fisher 344 male rats fed processed peaches: Rats fed control had significantly (p<0.05) lower cecal weight compared to rats fed treatment diets (Table 2). Among treatment groups, rats fed 2% juice and 4% STB FR had the lowest cecal weight. However, there were no significant (p<0.05) differences in cecal pH of rats fed processed peaches as compared to control, except for rats fed 4% STB FR (7.42±0.08). Among the treatment groups, rats fed 4% STB FR had the lowest cecal pH (7.42±0.08).
Effect of treatment diet on number of ACF in colon of AOM induced Fisher 344 Male rats: ACF in the proximal colon were lower compared to distal colon. As previously stated, ACF in humans are lower in proximal section of the colon compared to the distal portion.
| Table 1: | Processed Peaches on weight gain and feed intake in Fisher 344 male rats |
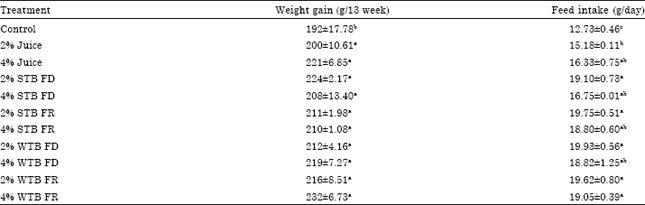 | |
| STB FD: Steam blanched freeze dry; STB FR: Steam blanched freezer; WTB FD; Water blanched freeze dry; WTB FR; Water blanched freezer. Values are Means±SEM; n = 4. Values not sharing a common superscript are significantly different (p<0.05) using Tukey’s studentized range test | |
| Table 2: | Processed peaches on cecal weight and cecal pH in Fisher 344 male rats |
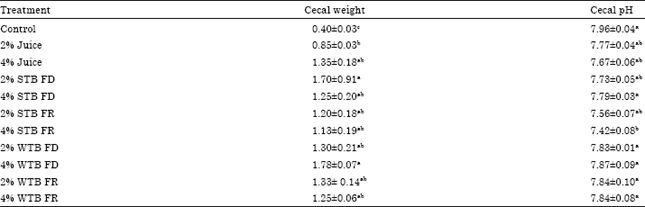 | |
| STB FD: Steam blanched freeze dry; STB FR: Steam blanched freezer; WTB FD; Water blanched freeze dry; WTB FR; Water blanched freezer. Values are Means±SEM; n = 4. Values not sharing a common superscript are significantly different (p<0.05) using Tukey’s studentized range test | |
 | |
| Fig. 2: | Aberrant crypts in proximal colon of Fisher 344 male rats fed processed peaches. Abbreviations: STB FD: Steam Blanched Freeze Dry; STB FR: Steam Blanched Freezer; WTB FD; Water Blanched Freeze Dry; WTB FR: Water Blanched Freezer. Values are Means. Bars not sharing a common superscript are significantly different (p<0.05) using Tukey’s Studentized Range Test |
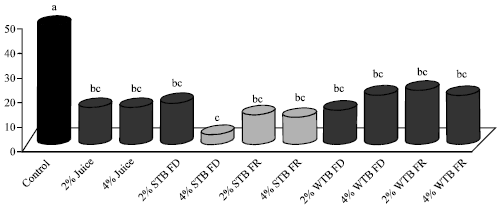
Rats fed treatment diets had significantly (p<0.05) lower ACF incidence (both in the distal and proximal colon) compared to control (Fig. 2, 3). Among rats fed treatment diets, there were no significant (p<0.05) differences in the number of ACF in rats fed 2% JUICE, 4% STB FR and 2% STB FD (Fig. 3). Groups fed 4% STB FD, 4% WTB FD and 4% WTB FR had no significant (p<0.05) differences in the number of ACF, however, rats fed 4% JUICE, 4%STB FR and 2% WTB FR had the lowest number of ACF as compared to the other treatment groups including the control (Fig. 3).
Compared to the control, there were 20-60% reductions in ACF numbers in the distal section of colon in rats fed treatment diet and 20-80% reductions in ACF numbers in the proximal section of colon.
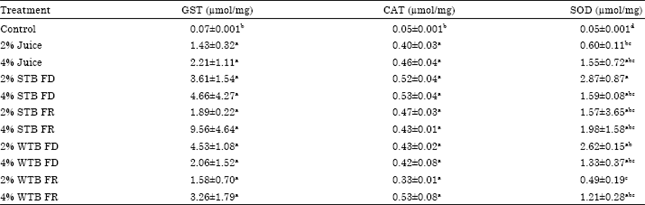
Total hepatic GST activity in Fisher 344 Male rats fed processed peaches: Compared to control, total glutathione-S-transferase (GST) activity (Mmol/mg) in liver of rats fed treatment diets were significantly (p<0.05) higher (Table 3).
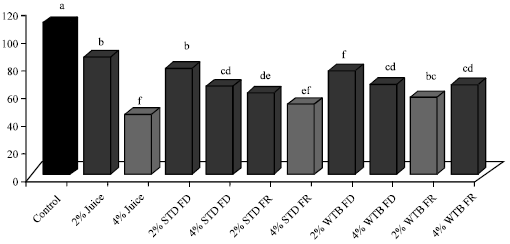 | |
| Fig. 3: | Aberrant crypts in distal colon of Fisher 344 male rats fed processed peaches. Abbreviations: STD FD: Steam Blanched Freeze Dry; STD FR: Steam Blanched Freezer; WTB FD; Water Blanched Freeze Dry; WTB FR; Water Blanched Freezer. Values are Means; abcBars not sharing a common superscript are significantly different (p<0.05) using Tukey’s Studentized Range Test |
| Table 3: | Hepatic enzyme activities in rats fed Processed Peaches |
 | |
| STB FD: Steam blanched freeze dry; STB FR: Steam blanched freezer; WTB FD; Water blanched freeze dry; WTB FR; Water blanched freezer. Values are Means±SEM; n = 4. Values not sharing a common superscript are significantly different (p<0.05) using Tukey’s studentized range test | |
However, there were no significant (p<0.05) differences in GST activity among rats fed treatment diets. Among the treatment groups, total GST activity was higher in rats fed steam blanched diets as compared to rats fed water blanched peach diets.
Total hepatic CAT activity in Fisher 344 Male rats fed processed peaches: Total Catalase (CAT) activity (μmol/mg) in liver of rats fed processed peaches was significantly (p<0.05) higher compared to control (Table 3) but there were no significant (p<0.05) differences among rats fed treatment diets. Among the treatment groups, rats fed 2 and 4% STB FD and 4% WTB FR had the highest CAT activity.
Total hepatic SOD activity in Fisher 344 Male rats fed processed peaches: Superoxide dismutase (SOD) is a metalloenzyme that catalyzes the dismutation of superoxide anion to molecular oxygen and hydrogen peroxide and thus forms a crucial part of the cellular antioxidant defense mechanism.
| Table 4: | Phenolic content, DPPH and FRAP activity in fresh and processed peaches |
 | |
| FU: Fresh unblanched; FU WTB: Fresh water blanched; FSTB: Fresh steam blanched; FR WTB: Freezer water blanched; FR STB: Freezer steam blanched; FD WTB: Freeze dry water blanched; FD STB; Freeze dry steam blanched; DPPH: Diphenyl-1-picrylhydrazyl; FRAP: Ferric reducing antioxidant power. Values are Means±SEM; n = 4. Values not sharing a common superscript are significantly different (p<0.05) using Tukey’s studentized range test | |
SOD activity (mmol/mg) was significantly (p<0.05) higher in rats fed treatment diet compared to rats fed the control diet (Table 3). However, rats fed 2% STB FD and 2% WTB FD had higher SOD activity compared to other treatment groups.
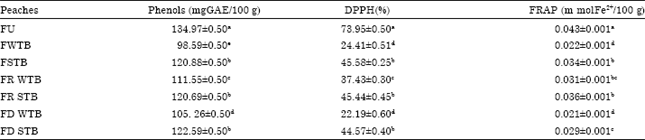
Total phenolic content and free radical scavenging activity of processed peaches: Many studies have suggested that the protective role of phenolic and flavonoid compounds in fruits and vegetables may be responsible for their anti-carcinogenic, antioxidative and anti-proliferative properties. Several researchers have suggested that it may be beneficial to utilize more than one antioxidant test due their specificity and the intricate nature of several food matrices. In this experiment, we utilized the (1, 1-Diphenyl-2-picryl-hydrazyl) DPPH radical solution and ferric reducing antioxidant power (FRAP) tests to obtain an overall profile of the total antioxidant capacity in fresh and processed peaches. The DPPH antioxidant test demonstrates the free radical scavenging activity of the antioxidants present in the peach extracts, whereas the FRAP assay demonstrates the antioxidants’ capacity to reduce the oxidizing species that could potentially damage biological molecules.
In fresh unblanched peaches, total phenolic content, FRAP and DPPH activities were significantly (p<0.05) higher compared to processed peaches (Table 4). Steam blanched freeze dried peaches and steam blanched frozen peaches had a significantly (p<0.05) higher phenolic content, FRAP and DPPH activity compared to the respective water blanched peaches. Steam blanched peaches retained most of its phenolic content and thus had a higher antioxidative activity as compared to water blanched peaches and this might be attributed to less leaching of bioactive components during processing.
DISCUSSION
The aim of this study was to investigate the inhibitory effects of processed peaches (meal) (STB FD, STB FR, WTB FD, WTB FR) at 2 and 4% concentrations and peach juice at 2 and 4% levels, respectively on the formation of AOM-induced ACF in Fisher 344 male rats, their effect on selected hepatic enzyme activities (GST, SOD and CAT) and to determine the effect of processing (Blanching+Freezing, Blanching+Freeze drying) on total phenolic content and antioxidant potential by (2, 2-diphenyl-1-picrylhydrazyl (DPPH) activity and Ferric Reducing Antioxidant Potential (FRAP) in Peaches.
Results from our study showed that AOM-induced ACF were significantly lower in rats fed processed peaches compared to rats fed the control diet. Also, selected hepatic and antioxidative enzyme activities were significantly enhanced in peach fed groups compared to the control.
Fruits provide an optimal mix of bioactive components such as phenolics, carotenoids and flavonoids. However, the health promoting capacity of fruits depends on their processing history since processing is expected to affect the content, activity and bioavailability of bioactive compounds. From this study, we noted that, blanching played a significant role in retention of phenolic compounds as shown by their antioxidative capacities (Table 4). According to Southon (1998) fruits and vegetables are blanched prior to freezing or freeze-drying. Blanching represents a useful tool in preventing enzymatic oxidation which is the main cause of loss of naturally occurring antioxidants in raw material of plant origin. However, blanching may have negative effects on nutrients such as vitamins and phenolic compounds which are relatively unstable when subjected to heat treatments (Prochaska et al., 2000). In this study, total phenolic content and antioxidant capacity in blanched peaches were significantly (p<0.05) lower than the fresh sample. These results agree with the findings of Xu and Chang (2008), who reported a loss of 40-50% in phenolic content and a decrease in DPPH activity of blanched green pea, yellow pea and chick pea due to leaching of soluble antioxidant components during processing compared to raw peas. Amin et al. (2006) reported a loss of 71% in total phenolic content in blanched Amaranthus compared to raw. Studies performed on strawberry (Klopotek et al., 2005), demonstrated a relationship between the decrease of antioxidant capacity and phenolic content and the processing necessary to obtain pasteurized juice.
Previous studies have shown contradictory findings on the effects of blanching on total phenolic content and antioxidant capacity of fruits and vegetables. Oboh and Akindahunsi (2004) reported an increase in phenolic content up to 200% after blanching green leafy vegetables. This increment was suggested due to breakdown of tannins during the heating process which increased their extractability. Regardless of the findings, antioxidant capacity has been closely related to phenolics, flavonoids and other compounds present in fruits and vegetables. In this regard, ACF was significantly lower in rats fed blanched peaches either freeze-dried or conventionally frozen compared to those fed the control diet. Research has shown that application of thermal processing over a long period of time leads to rapid release of bound bioactive components which are found in the natural matrix of the fruit, thus, making it more bioavailable (Dewanto et al., 2002). Phenolic and flavonoid content in processed peaches may partly explain the protective effects attributed to the significant (p<0.05) decrease in crypt formation, since antioxidants (phenolics) have the ability to reverse, suppress or prevent progression of disease from pre-invasive cancer to frank malignancy (Sporn et al., 1976). Conventionally they have been classified into either blocking or suppressing agents. Blocking agents, where they prevent the occurrence of the damage by inhibiting the activation of a carcinogen, inhibiting its interaction with vital cellular macromolecules or by inducing rapid deactivation and clearance of the carcinogen. Suppressing agents, on the other hand, control or reverse the damage caused by a carcinogen.
Reduction in ACF incidence in rats fed processed peaches may also be linked to their soluble fermentable fiber content. According to Hill (1998), fruits such as peaches are fiber-rich foods. Dietary Fiber (DF) mainly fermentable fiber may influence colon risk through proposed mechanisms such as increasing fecal bulk (diluting carcinogens); increasing transit time through the colon (reducing interactions of carcinogens with mucosal cells); direct binding of carcinogens; modifying the mix and enzyme activities of intestinal bacterial flora (decreasing concentrations of secondary bile acids) and producing Short-Chain Fatty Acids (SCFAs) by fermentation which may inhibit carcinogenesis through effects on colonic pH and increased availability of butyrate. Butyrate promoted growth arrest, differentiation and apoptosis in colon and breast cancer cell lines (Hill, 1998). According to Kahlon and Smith (2007), bile acid binding which is one of many mechanisms by which certain foods prevent cancer was higher in peaches compared to grapes, pears and apricots. Binding of bile acids and prevention of their recirculation resulted in reduced fat absorption, excretion of cancer causing toxic metabolites and cholesterol and this is believed to be the mechanism by which food fractions lower cholesterol and prevent cancer. This according to Kahlon and Smith (2007) may be due to the soluble fermentable fiber content in the fruit. Peaches contain mainly phenolic acids, chlorogenic and neo-chlorogenic acids; the flavan-3-ols, catechin and its derivatives and procyanidins; the flavonols, quercetin and its derivatives and small amounts of anthocyanins, cyanidin 3β-glucoside and cyanidin 3β-rutinoside (Tomas-Barberan et al., 2001). These phenolics have been the subject of many health-related claims supported by in vitro and in vivo studies and related to their properties as antioxidants and anticarcinogenic dietary compounds (Dechsupa et al., 2007; Granado-Serrano et al., 2007; Hou et al., 2003). Noratto et al. (2009) confirmed that phenolic acids present in peaches may have potential as chemopreventive dietary compounds because of relatively high growth inhibition exerted on the estrogen-independent MDA-MB-435 breast cancer cell line and low toxicity exerted in the normal MCF-10 A cells. Noratto et al. (2009) suggested that peach phenolics, mainly chlorogenic and neo-chlorogenic acids may have the potential in therapy and chemoprevention of metastatic breast cancer.
One of the known chemopreventive activity of phytochemicals is the induction of phase II metabolizing enzymes such as Glutathione-S-Transferase (GST), Superoxide-Dismutase (SOD), and Catalase (CAT) according to Block and Gyllenhaal (2002). Polyphenols may be regarded as xenobiotics by animal cell due to their interaction with phase I and phase II enzymes according to Moskaug et al. (2004). Chemical carcinogens may be activated via phase I metabolism or detoxified by phase II enzymes. Therefore, modulating phase I and phase II enzyme activity by phytochemicals can be protective or detrimental depending on the nature of the chemical carcinogen and the type of modulation, i.e., induction or inhibition of carcinogen activation and detoxification. Induction is often accomplished via activation of the expression of the enzyme, whilst inhibition on the other hand, is often due to direct inhibition of metabolic activity of the enzyme (Block and Gyllenhaal, 2002), or occasionally, to inhibition of its gene expression.
In this study, we saw a significant increase in hepatic and antioxidative enzyme activities (GST, SOD and CAT) in rats fed processed peaches compared to rats fed the control diet. Catalase is one of the important antioxidative enzymes involved in removal of hydrogen peroxide produced by the action of Superoxide dismutase (SOD), we observed a 10-50 fold increase in hepatic CAT and SOD activity in rats fed processed peach diets compared to rats fed the control diet indicating that phytochemicals present in processed peaches may have the ability to induce antioxidative enzymes such as CAT and SOD.
CONCLUSION
We have shown that, processed peaches (meal+juice) at 2 and 4% level inhibited the incidence of AOM-induced ACF in Fisher 344 male rats and significantly (p<0.05) enhanced hepatic and antioxidative enzyme activities in Fisher 344 male rats. This research also suggests that the phenolic content and free radical scavenging activity in processed peaches used in this study may also be a potential mechanism resulting in the lower number of ACF. The results from this study are preliminary, further long-term studies including end-point tumor models and clinical trials may be necessary for a more conclusive evaluation of this study.
ACKNOWLEDGMENTS
This study was supported by funding from The Agricultural Experimental Research Station, Special thanks to Winfred Thomas Agricultural Research Station, Alabama Agricultural and Mechanical University.
REFERENCES
- Amin, I., Y. Norazaidah and K.I.E. Hainida, 2006. Antioxidant activity and phenolic content of raw and blanched Amaranthus species. Food Chem., 94: 47-52.
CrossRefDirect Link - Benzie, I.F.F. and J.J. Strain, 1999. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Method Enzymol., 299: 15-27.
CrossRefPubMedDirect Link - Bingham, S.A., 1990. Mechanisms and experimental and epidemiological evidence relating dietary fibre (non-starch polysaccharides) and starch to protection against large bowel cancer. Proc. Nutr. Soc., 49: 153-171.
PubMed - Bird, R.P., 1987. Observation and quantification of aberrant crypts in the murine colon treated with a colon carcinogen: Preliminary findings. Cancer Lett., 37: 147-151.
PubMedDirect Link - Bird, R.P., 1995. Role of aberrant crypt foci in understanding the pathogenesis of colon cancer. Cancer Lett., 93: 55-71.
CrossRefPubMedDirect Link - Block, K.I. and C. Gyllenhaal, 2002. Clinical corner: Herb-drug interactions in cancer chemotherapy: Theoretical concerns regarding drug metabolizing enzymes. Integr. Cancer Ther., 1: 83-89.
PubMed - Bresnick, E., D.F. Birt, K. Wolterman, M. Wheeler and R.S. Markin, 1990. Reduction in mammary tumorigenesis in the rat by cabbage and cabbage residue. Carcinogenesis, 11: 1159-1163.
PubMed - Dechsupa, S., S. Kothan, J. Vergote, G. Leger and A. Martineau et al., 2007. Quercetin, siamois 1 and siamois 2 induce apoptosis in human breast cancer MDA-mB-435 cells xenograft in vivo. Cancer Biol. Ther., 6: 56-61.
PubMed - Dewanto, V., X. Wu, K.K. Adom and R.H. Liu, 2002. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem., 50: 3010-3014.
CrossRefDirect Link - Fridivich, I., 1989. Superoxide dismutases: An adaptation to a paramagnetic gas. J. Biol. Chem., 264: 7761-7764.
PubMedDirect Link - Gil, M.I., F.A. Tomas-Barberan, B. Hess-Pierce and A.A. Kader, 2002. Antioxidant capacities, phenolic compounds, carotenoids, and vitamin C contents of nectarine, peach, and plum cultivars from California. J. Agric. Food Chem., 50: 4976-4982.
CrossRefPubMedDirect Link - Noratto, G., W. Porter, D. Byrne and L. Cisneros-Zevallos, 2009. Identifying peach and plum polyphenols with chemopreventive potential against estrogen-independent breast cancer cells. J. Agric. Food Chem., 57: 5219-5226.
CrossRef - Granado-Serrano, A.B., M.A. Martin, M. Izquierdo-Pulido, L. Goya, L. Bravo and S. Ramos, 2007. Molecular mechanisms of (-)-epicatechin and chlorogenic acid on the regulation of the apoptotic and survival/proliferation pathways in a human hepatoma cell line. J. Agric. Food Chem., 55: 2020-2027.
CrossRef - Hou, R.C.W., H.L. Chen, J.T.C. Tzen and K.C.G. Jeng, 2003. Effect of sesame antioxidants on LPS-induced NO production by BV2 microglial cells. Neuroreport, 14: 1815-1819.
Direct Link - Huber, G.M. and H.P. Rupasinghe, 2009. Phenolic profiles and antioxidant properties of apple skin extracts. J. Food Sci., 74: C693-C700.
CrossRefDirect Link - Jackson, P., D. Cooper, P. O'Connor and A. Povey, 1999. The relationship between 1, 2 dimethylhydrazine dose and the induction of colon tumors: Tumor development in female SWR mice does not require a K-ras mutational event. Carcinogenesis, 20: 509-513.
PubMed - Jackson, L., W. Wahli, L. Michalik, S.A. Watson and T. Morris et al., 2003. Potential role for peroxisome proliferator activated receptor (PPAR) in preventing colon cancer. Gut, 52: 1317-1322.
Direct Link - Johansson, L.H. and L.A. Borg, 1988. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem., 174: 331-336.
CrossRefPubMedDirect Link - Kahlon, T.S. and G.E. Smith, 2007. In vitro binding of bile acids by bananas, peaches, pineapple, grapes, pears, apricots and nectarines. Food Chem., 101: 1046-1051.
CrossRef - Kampa, M., A. Hatzoglou, G. Notas, A. Damianaki and E. Bakogeorgou et al., 2000. Wine antioxidant polyphenols inhibit the proliferation of human prostate cancer cell lines. Nutr. Cancer, 37: 223-233.
PubMed - Kim, D.O., S.W. Jeong and C.Y. Lee, 2003. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem., 81: 321-326.
CrossRefDirect Link - Meyer, F., P. Galan, P. Douville, I. Bairati and P. Kegle et al., 2005. Antioxidant vitamin and mineral supplementation and prostate cancer prevention in the SU.VI.MAX trial. Int. J. Cancer, 116: 182-186.
CrossRefDirect Link - Moskaug, J.O., H. Carlsen, M. Myhrstad and R. Blomhoff, 2004. Molecular imaging of the biological effects of quercetin and quercetin-rich foods. Mech. Ageing Dev., 125: 315-324.
CrossRef - Nosho, K., H. Yamamoto, M. Mikami, H. Taniguchi and T. Takahashi et al., 2006. Overexpression of poly(ADP-ribose) polymerase-1 (PARP-1) in the early stage of colorectal carcinogenesis. Eur. J. Cancer, 42: 2374-2381.
CrossRef - Oboh, G. and A.A. Akindahunsi, 2004. Change in the ascorbic acid, total phenol and antioxidant activity of sun-dried commonly consumed green leafy vegetables in Nigeria. Nutr. Health, 18: 29-36.
CrossRefDirect Link - Prochaska, L.J., X.T. Nguyen, N. Donat and W.V. Piekutowski, 2000. Effects of food processing on the thermodynamic and nutritive value of foods: Literature and database survey. Med. Hypotheses, 54: 254-262.
CrossRef - Reeves, P.G., F.H. Nielsen and G.C. Fahey Jr., 1993. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr., 123: 1939-1951.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Sohn, K.J., S.A. Shah, S. Reid, M. Choi and J. Carrier et al., 2001. Molecular genetics of ulcerative colitis-associated colon cancer in the interleukin 2-and beta(2)-microglobulin-deficient mouse. Cancer Res., 61: 6912-6917.
PubMed - Sporn, M.B., N.M. Dunlop, D.L. Newton and J.M. Smith, 1976. Prevention of chemical carcinogenesis by vitamin A and its synthetic analogs (retinoids). Fed. Proc., 35: 1332-1338.
PubMedDirect Link - Steinmetz, K.A. and J.D. Potter, 1996. Vegetables, fruit and cancer prevention: A review. J. Am. Dietetic Assoc., 96: 1027-1039.
CrossRefDirect Link - Takahashi, T., T. Takahashi, H. Suzuki, T. Hida, Y. Sekido, Y. Ariyoshi and R. Ueda, 1991. The p53 gene is very frequently mutated in small-cell lung cancer with a distinct nucleotide substitution pattern. Oncogene, 6: 1775-1778.
PubMed - Temple, N. and K.K. Gladwin, 2003. Fruit, vegetables and the prevention of cancer: Research challenges. Nutrition, 19: 467-470.
Direct Link - Tomas-Barberan, F.A., M.I. Gil, P. Cremin, A.L. Waterhouse, B. Hess-Pierce and A.A. Kader, 2001. HPLC-DAD-ESIMS analysis of phenolic compounds in nectarines, peaches and plums. J. Agric. Food Chem., 49: 4748-4760.
CrossRefDirect Link - Van't Veer, P., M.C. Jansen, M. Klerk and F.J. Kok, 2000. Fruits and vegetables in the prevention of cancer and cardiovascular disease. Public Health Nutr., 3: 103-107.
CrossRef - Wattenberg, L.W. and J.B. Coccia, 1991. Inhibition of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone carcinogenesis in mice by D-limonene and citrus fruit oils. Carcinogenesis, 12: 115-117.
PubMed - Xu, B. and S.K.C. Chang, 2008. Characterization of phenolic substances and antioxidant properties of food soybeans grown in the North Dakota-Minnesota region. J. Agric. Food Chem., 56: 9102-9113.
CrossRefDirect Link - Boateng, J., M. Verghese, C.B. Chawan, L. Shackelford, L.T. Walker, J. Khatiwada and D.S. Williams, 2006. Red palm oil suppresses the formation of azoxymethane (AOM) induced aberrant crypt foci (ACF) in fisher 344 male rats. Food Chem. Toxicol., 44: 1667-1673.
CrossRefPubMedDirect Link - Brand-Williams, W., M.E. Cuvelier and C. Berset, 1995. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol., 28: 25-30.
CrossRefDirect Link