
D. Asiamah
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal AL, Alabama, USA
M. Verghese
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal AL, Alabama, USA
J. Boateng
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal AL, Alabama, USA
B. Kanda
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal AL, Alabama, USA
L. Shackelford
Nutritional Biochemistry and Carcinogenesis Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal AL, Alabama, USA
L.T. Walker
Food Chemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, Normal AL, Alabama, USA
International Journal of Cancer Research
Year: 2011 | Volume: 7 | Issue: 1 | Page No.: 36-46







ABSTRACT
Bitter Melon, (BM) Momordica charantia is known in Asia for regulation of blood glucose levels and as a treatment for Diabetes mellitus. However, studies on its effect in preventing cancer are scarce. This study was conducted to investigate the chemopreventive properties of bitter melon on azoxymethane (AOM)-induced Aberrant Crypt Foci (ACF) in Fisher 344 male rats and determine its effects on selected hepatic detoxification and antioxidant enzymes. Rats were fed AIN93G as control (C) and treatment diets containing 2 and 4% BM. At 7 and 8 weeks of age, ACF was induced by administering 2 s/c injections of AOM at 16 mg kg-1 b.wt. and killed by CO2 asphyxiation at 17 weeks of age. Total ACF in proximal and distal colons were 35, 26 and 21; 116, 54 and 38 in rats fed C, 2% BM and 4% BM, respectively. Catalase and SOD activities (μmol mg-1) were higher in rats fed treatment diets (0.293 and 0.03; 1.82 and 0.27 for rats fed 2 and 4% BM, respectively) compared to control (0.04). Glutathione-S-Transferase (GST) (μmol mg-1) activity and Glutathione (GSH) levels were higher in treatment groups compared to control. Physicochemical analysis was conducted on bitter melon fruit (total phenolics, free radical scavenging activity (DPPH) and Ferric Reducing Antioxidant Power (FRAP)). Total phenolic content was 34.31 mg/GAE/g fresh fruit, DPPH activity was 2.99% and FRAP was 0.652 μmol Fe2+/g/mL. Results indicate bitter melon reduced incidence of AOM induced ACF. Thus inclusion of BM in diets may have possible implications in reducing the risk of colon cancer.
PDF Abstract XML References Citation
Received: July 29, 2010;
Accepted: November 03, 2010;
Published: April 22, 2011
How to cite this article
D. Asiamah, M. Verghese, J. Boateng, B. Kanda, L. Shackelford and L.T. Walker, 2011. Chemopreventive Potential of Bitter Melon (Momordica charantia) against Precancerous Lesions in the Colon of Fisher 344 Male Rats. International Journal of Cancer Research, 7: 36-46.
DOI: 10.3923/ijcr.2011.36.46
URL: https://scialert.net/abstract/?doi=ijcr.2011.36.46
DOI: 10.3923/ijcr.2011.36.46
URL: https://scialert.net/abstract/?doi=ijcr.2011.36.46
INTRODUCTION
Colon cancer is the third leading cause of cancer deaths worldwide. It is estimated that there are about one million new cases each year and about half a million deaths caused by colon cancer each year worldwide (Parkin et al., 2005). One out of every four deaths in the United States is caused by cancer (American Cancer Society, 2008). Colon cancer is the third most diagnosed cancer in both males and females. In 2009 there were about 106,100 cases (American Cancer Society, 2009). Research has shown that a significant number of colon cancer cases can be prevented if nutritional intervention is pursued. Various studies have shown that diets rich in fruits and vegetables offer protection against the incidence of colorectal cancer (Riboli and Norat, 2003; Zampino et al., 2008).
Bitter melon is a tropical vegetable of the taxonomic family cucurbitaceae. Bitter melon is relatively new to the Western world and its bitter taste does not make it a vegetable of choice. However, the bitterness can be reduced through heat application (Wang et al., 2008a). Traditionally bitter melon is used in tropical countries for the treatment of diabetes. Though data on the chemical and biological properties of bitter melon remain scarce, its folk medicinal benefits are widely known. Bitter melon contains numerous compounds, few of which include ferulic acid, caffeic acid (Raj et al., 2005), momordin, momorcharin, cryptoxanthin (Grover and Yadav, 2004), isorhamnetin and fisetin (Lako et al., 2007). Ferulic acid has the potential of scavenging superoxides (Ogiwara et al., 2002), caffeic acid which is a phenolic acid is found in most plants (Arimboor et al., 2008) and momordin a ribosome-inactivation protein (Leamon and Low, 1992) has cytotoxic effects on leukemia cells (Kim et al., 2002). The proteins α and β-momorcharin have the potential of reducing the activity of the HIV virus (Taylor, 2002). Fisetin is a flavonol which was shown to improve memory in laboratory rats (Troup et al., 2007), isorhamnetin is also a flavoniod inhibits superoxide radicals and lipid peroxidation (Zielinska et al., 2001). Bitter melons also possess antineoplastic properties (Basch et al., 2003), however information on cancer studies using bitter melon are scarce, specifically dietary chemoprevention studies with respect to colon carcinogenesis. The selection of bitter melon was warranted due to studies indicating its numerous medical benefits (Lee-Huang et al., 1995). Bitter melon contains momorcharin (Grover and Yadav, 2004), a plant protein which has exhibited antitumor properties in other studies (Ng et al., 1992). The vegetable has also been shown to contain the protein MAP30, which has anti-HIV properties (Lee-Huang et al., 1995). However, there is no published work on the potential of bitter melon in colon cancer prevention.
Azoxymethane is a colon specific carcinogen (Yamada and Mori, 2003). It is used to induce Aberrant Crypt Foci (ACF) which are preneoplastic lesions (Ochiai et al., 1996). ACF are the earliest identifiable precursors of colon cancer (Pretlow et al., 1991). In most colon cancer preventive studies, Fisher 344 male rats are used due to their reliability and clinical features resembling human colon cancer (Takahashi and Wakabayashi, 2004). Therefore, the objectives of this study were to investigate the chemopreventive properties of bitter melon on AOM-induced ACF in Fisher 344 male rats and to determine its effects on selected hepatic detoxification and antioxidant enzymes.
The selected hepatic detoxification enzyme was glutathione S-transferase (GST), selected antioxidant enzymes were superoxide dismutase (SOD) and catalase (CAT). Catalase is a peroxisomal enzyme which breaks down hydrogen peroxide into water and oxygen (Sato et al., 1992). It works together with other enzymes in providing defense against oxidants. Glutathione S-transferase enzymes can metabolize carcinogens and they also play a role against other xenobiotics (Pool-Zobel et al., 2005). These enzymes are found in yeast, bacteria, insects and mammals. Superoxide dismutase protects the cell by preventing damage caused by Reactive Oxygen Species (ROS) which readily attack cells (Miyazawa et al., 2009). Hydrogen peroxide which is produced from superoxide anion (O2-) is metabolized into water and oxygen by glutathione (Miyazawa et al., 2009). These enzymes were selected due to their role in chemoprevention. They may also serve as biomarkers for colon cancer.
MATERIALS AND METHODS
Reagents and standards: Methanol, buffered formalin, Azoxymethane (NCI Chemical Repository, Kansas City, MO), 1-chloro 2, 4-dinitrobenzene, glutathione, hydrogen peroxide, potassium hydroxide, purpald (chromogen), potassium periodate, formaldehyde standard, 0.05 M carbonate buffer (pH 10.2), 0.3 mM adrenaline, Ellman’s reagent, 4% sulfosalicylic acid, Folin-Ciocalteu’s phenol reagent, 7% NaCO3, gallic acid standard, FRAP reagent, 300 mM acetate buffer (pH 3.6), 10 mM 2,4,6-tri (2-pyridyl)-s-triazine (TPTZ), 40 mM HCl, 20 mM ferric chloride (FeCL3.H2O), ferrous sulphate (FeSO4.7H2O) (0.1-1.0 mM), DPPH (1, 1-Diphenyl-2-picryl-hydrazyl) radical solution and 10% buffered formalin were purchased from Fisher Scientific (Suwannee, Georgia).
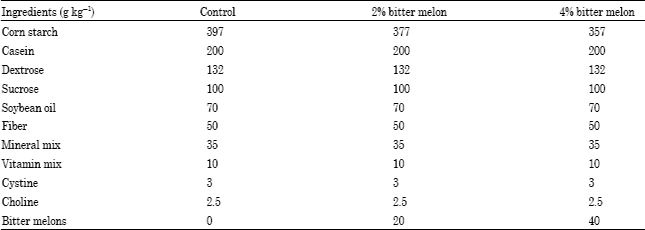
Preparation of diets: Mature Bitter Melons (BM) were purchased from a local farmers market, Huntsville Al. They were cleaned, sliced and freeze dried (Virtis genesis freeze drier) (Woodstock, Georgia) and ground to fine powder (Robot Coupe food processor Blixer RSIBX3) (Ridgeland, Mississippi). The powdered sample was mixed in an AIN93G basal diet (powdered) at 2 and 4% levels (Table 6). For chemical analysis, extracts were prepared using fresh bitter melon with 80% methanol. Samples were concentrated using a Welch rotary evaporator (Model 2025, Sheboygan, Wisconsin).
Animal study: This study was conducted in the Department of Food and Animal Sciences at Alabama A and M University (2009). A total of 18 rats (6 rats/group) were used in this study. Rats were obtained from Harlan (IN) and were randomly assigned in pairs and housed in wire cages. Temperature and humidity were kept at 21°C and 50%, respectively and light and dark cycles were maintained at 12 h each for the entire duration of the study (13 week). Animals were allowed a one week acclimatization period, after which they were randomly assigned into three groups. The control group was fed a diet based on formulations from the American Institute of Nutrition (AIN-G) (Reeves et al., 1993) and the treatment groups were fed an AIN93G based diet containing 2 and 4% BM which was added at the expense of corn starch. Feed intakes (g day-1) and body weight (g) were recorded daily and weekly, respectively. All work with animals was approved by the Alabama A and M University.
Sample and tissue collection: At 7 and 8 weeks of age, rats received injections of Azoxymethane (AOM) (NCI Chemical Repository, Kansas City, MO) in saline at 16 mg kg-1 b.wt. Rats were killed by CO2 asphyxiation at 17 weeks of age. Livers were excised and stored at -80°C until enzyme analysis. Cecal weight and cecal pH was noted. Colons were removed and fixed in buffered formalin and Aberrant Crypt Foci (ACF) were enumerated as described by Bird (1987).
Enzyme analysis
Total Glutathione S-Transferase (GST) activity: Glutathione S-transferase activity in the liver of the rats was assayed according to protocol outlined by Habig et al. (1974), using CDNB as the substrate. GST was determined at an absorbance of 340 nm at 1 min intervals for 5 min and activity was expressed as μmol mg-1.
Catalase (CAT) and superoxide dismutase (SOD) activity: Liver catalase activity was determined at ambient temperature according to the procedure described by Aebi (1984) with modifications. To a 96 well culture plate, 20 μL of appropriately diluted sample was added to 100 μL of assay buffer and 30 μL of methanol. In each well, catalytic reactions were initiated by adding 20 μL of hydrogen peroxide noting the precise time the reaction is initiated. The samples (wells) were gently agitated using the shaker, simultaneously at room temperature for 20 min. After 20 min, 30 μL of potassium hydroxide was added to each well to terminate the reaction. Each well had 30 μL of purpald (chromogen) subsequently added to it. For an additional 10 min, the plate was gently agitated using a shaker and incubated at room temperature. Finally, 10 μL of potassium periodate was added to each well. The plate was gently agitated and incubated for an additional 5 min at room temperature. Catalase activity was determined against a formaldehyde standard, at an absorbance of 540 nm using a Microplate reader (Biotek Synergy HT, Winooski, Vermont) and expressed as μM of H2O2 decomposed/min/mL protein.
Hepatic superoxide dismutase was assayed using Fridovich (1989) �s method . Homogenized livers were added to 2.5 mL of 0.05 M carbonate buffer (pH of 10.2). The reaction was started with the addition of 0.3 mL freshly prepared 0.3 mM adrenaline to the mixture which was quickly mixed by inversion. Buffer (2.5 mL), 0.3 mL of substrate (adrenaline) and 0.2 mL of water was placed in the reference cuvette. A change in the absorbance at 480 nm was monitored every 30 for 150 sec. A single unit of enzyme is defined as the quantity of superoxide dismutase added to produce 50% inhibition of autoxidation.
Glutathione (GSH) levels: Hepatic glutathione was assayed using the colorimetric method and Ellman’s reagent was used. Briefly, livers were homogenized in 4 volume of ice-cold homogenizing buffer at pH 7.4. The aliquot of the homogenate was deproteinized by addition of an equal volume of 5% metaphosphoric acid and centrifuged at 17,000 g for 15 min at 2°C. An aliquot of the supernatant (0.5 mL) was added to 4.5 mL of Ellman’s reagent. GSH levels were determined at an absorbance of 412 nm. A standard curve was prepared by plotting different concentrations of GSH.
Chemical analysis
Total phenolic: The total phenolic content in bitter melon was determined using Folin-Ciocalteu’s reagent. The method was modified for the micro plate reader. Briefly, bitter melon extracts (25 μL) were added to 50 μL of distilled water. Further, 125 μL of Folin-Ciocalteu’s phenol reagent was added to the mixture (1:10). After 5 min 100 μL of 7% NaCO3 was added the mixture was allowed to incubate for 90 min at 25°C in the dark. Absorbance was measured using the micro plate reader (Biotek Synergy HT, Winooski, Vermont), at 750 nm versus a blank consisting of distilled water. Gallic acid standard solution was used to develop a standard curve for total phenolics. Total phenolics was expressed as mg Gallic Acid Equivalents (GAE) per 100 g of bitter melon.
Free Radical Scavenging Activity (FRSA) (DPPH): FRSA of bitter melon was analyzed as described by Brand-Williams (Brand-Williams et al., 1995). Extracts were diluted to various concentrations and 0.1 mL of each sample volume was added to 2.9 mL 1,1-Diphenyl-2-picryl-hydrazyl (DPPH) radical solution. The DPPH radical was dissolved in 80% methanol. The mixture was agitated and incubated at room temperature for 30 min. The experiment was conducted under restricted light. Free radical scavenging activity was measured as the amount of extract required to decrease the initial absorbance (wavelength 517 nm) of the DPPH radical concentration by 50% as compared to the control according to the equation below:
Ferric Reducing Antioxidant Power (FRAP): The ferric reducing antioxidant power assay was conducted as explained by Benzie and Strain (1996). Approximately 100 μL of diluted bitter melon extracts was combined with 3 mL of FRAP reagent consisting of 300 mM acetate buffer (pH 3.6), 10 mM 2,4,6-tri (2-pyridyl)-s-triazine (TPTZ) in 40 mM HCL and 20 mM ferric chloride (FeCL3.H2O). The diluted mixture was then incubated for 4 min at a temperature of 37°C. Following incubation, the samples were read at an absorbance of 593 nm. Absorbance readings were taken for 2 min over a period of 10 min. The change in absorbance was compared to standard ferrous sulphate (FeSO4.7H2O) (0.1-1.0 mM). The samples were analyzed in 4 replicates and the concentration of Fe3+ were expressed as μmol of Fe2+/g/mL. The FRAP reagent was freshly prepared as needed.
Statistical analysis: Data were analyzed using SAS system version 9.0 (SAS, 2004) by analysis of variance. The values were given as Means±SEM. A one way ANOVA was used in determining the means. A follow up test, Tukey’s studentized range test was used in separating the means. Significance were tested at p<0.05 level.
RESULTS
Body weight, feed intake, cecal weight and cecal pH in rats: Table 1 shows the weight gain (g 13 week-1) and feed intake (g day-1) in rats fed control and BM diets. The group fed 4% BM had significantly (p<0.05) higher feed intake compared to the control and 2% BM. This was reflected in their weight gain, where the group fed 4% BM had significantly (p<0.05) higher weight gain compared to the control group. There were no significant differences observed in cecal weight and cecal pH among control and treatment groups (Table 1).
Aberrant Crypt Foci (ACF): The incidence of Aberrant Crypt Foci (ACF) is shown in Table 2. Rats fed BM had significantly (p<0.05) lower ACF numbers compared to the control. ACF incidence in the distal colon was significantly (p<0.05) higher compared to the proximal colon in all experimental groups. However, the number of ACF in both the proximal and distal colon in rats fed control was significantly (p<0.05) higher compared to the treatment groups (151±9.44, 80.86±8.69 and 59±7.95, respectively). ACF with 3, 4 and 5 crypts have a higher propensity of developing into tumors.
| Table 1: | Weight gain, feed intake cecal weight and pH of Fisher 344 male rats |
 | |
| Means in a column with different superscripts are significantly (p<0.05) different using Tukey’s studentized range test. BM: Bitter melon, C: Control | |
| Table 2: | Incidence of Aberrant Crypt Foci (ACF) in Fisher 344 male rats fed bitter melon and control |
 | |
| BM: Bitter melon, C: Control. Mean in a row with different superscripts are significantly different (p<0.05) using Tukey’s studentized range test. xyMean in a column with different superscripts are significantly different (p<0.05) using Tukey’s studentized range test | |
| Table 3: | Crypt multiplicity in Fisher 344 male rats fed bitter melon and control |
 | |
| BM: Bitter melon, C: Control | |
| Table 4: | Total abnormal crypts in Fisher 344 male rats fed bitter melon and control diet |
 | |
| Means in a column with different superscripts are significantly (p<0.05) different using Tukey’s studentized range test. BM: Bitter melon C: Control | |
| Table 5: | Activity of selected enzymes and glutathione levels in Fisher 344 male rats |
 | |
| Means in a row with different superscripts are significantly (p<0.05) different using Tukey’s studentized range test. BM: Bitter melon C: Control, GST: Glutathione S-Transferase activity, SOD: Superoxide dismutase activity, CAT: Catalase activity, GSH: Glutathione levels | |
The control group had a higher incidence of ACF with larger crypts compared to the treatment groups (Table 3). Present results (Table 4) show that the control group had significantly (p<0.05) higher number of abnormal crypts in both the proximal and distal colon, compared to the rats fed 2 and 4% BM (514, 200 and 130, respectively). There was about a 60% and 75% decrease in total crypts in the groups fed 2 and 4% BM, respectively, compared to the control group.
Effects of diets on enzyme activity: Glutathione S-Transferase is a phase II detoxification enzyme. We observed that the rats fed 4% BM had significantly (p<0.05) higher GST activity compared to rats fed 2% BM and control (21.57, 16.89 and 10.55, respectively). Glutathione (GSH) levels, in the rats fed 4 and 2% BM diet were significantly (p<0.05) higher compared to the control (0.68, 0.49 and 0.17, respectively).
GST uses GSH as a substrate in the detoxification process. Hence, the higher the GSH levels the higher the detoxification process. The treatment groups (2 and 4% BM) had significantly (p<0.05) higher selected antioxidant enzymes activities compared to the control. Superoxide dismutase (SOD) activity in treatment groups was significantly (p<0.05) higher compared to the control. Catalase (CAT) activity in rats fed 4 and 2% was significantly higher compared to the control (1.82, 0.293 and 0.04, respectively). CAT activity in rats fed 4% BM diet was about 98% higher compared to the control group. Rats fed the diet containing 4% BM also had significantly (p<0.05) higher enzyme activities compared to the group fed 2% BM and the control group (Table 5).
| Table 6: | Composition of diets based on AIN93G |
 | |
Chemical analysis: Total phenolic content in bitter melon was 34.31 mg/GAE/g sample (fresh weight). Free radical scavenging activity (DPPH) was conducted on the fresh sample of bitter melon. Free radical scavenging activity was 0.647 mg mL-1, in percentage the value obtained was 2.99%. The ferric reducing antioxidant power of bitter melon was recorded to be 0.652 μmol Fe2+/g/mL.
DISCUSSION
The present animal study was conducted to evaluate the chemopreventive potential of bitter melon. The results of the study indicate that administration of bitter melon significantly (p<0.05) reduced the incidence of ACF. Several studies have supported the use of ACF as a reliable biomarker in colon cancer (Verghese, 2000; Williams et al., 2007; Boateng et al., 2007). The ability of bitter melon to reduce the incidence of ACF may be due to the compound momordin, which is found in bitter melon. Momordin has been reported to have antiproliferative effects (Kim et al., 2002). Comparable results were observed in a similar animal study conducted by Boateng et al. (2007), where selected fruits were fed to Fisher 344 male rats. Another animal study carried out using lycopene at different levels resulted in a reduced incidence of ACF in Fisher 344 male rats. This supports the hypothesis that fruits and vegetable may have antiproliferative properties. In another study a fermented soy product was shown to have no preventive role in the development of ACF in Wistar male rats (Silva et al., 2009). It was suspected that the soy fermented product was unable to prevent the clonal proliferation of the abnormal cells. Enumeration of ACF can be used to determine factors that affect the initiation phase of colon carcinogenesis (Fenoglo-Preiser and Noffsinger, 1999). Bitter melon may have reduced the incidence of ACF due to its antiproliferative properties. Bitter melon contains cryptoxanthin (Grover and Yadav, 2004), which was shown to have had mild protective effects against colorectal cancer in males in an epidemiological study (Park et al., 2009). Cryptoxanthin has also been shown to reduce the incidence of colon carcinogenesis in Fisher 344 male rats (fed mandarin oranges) by inducing apoptosis (Tanaka et al., 2000). In the present study cryptoxanthin in the bitter melon may have aided in the reduction of ACF incidence.
It was also observed that rats fed BM had higher activities of hepatic detoxification enzymes (GST) and antioxidant enzymes (SOD and CAT). The treatment groups had significantly (p<0.05) higher GST, SOD and CAT activity compared to the control. The GSH levels in the rats fed BM was also found to be significantly (p<0.05) higher compared to the control. GSH is utilized by GST as a substrate in the detoxification process. Superoxide dismutase catalyzes the dismutation of superoxides, which are potent carcinogens. They are converted to hydrogen peroxide (H2O2) and molecules of oxygen. CAT and SOD work in sync in the prevention ACF formation, where the H2O2 produced by the SOD is further reduced to molecules of water by CAT. The groups fed 2 and 4% BM diets had significantly (p<0.05) higher CAT and SOD activities. This is supported by an earlier study, where pretreatment of momordin sustained the activities of glutathione, glutathione reductase, glutathione S-transferase, glutathione peroxidase, catalase and superoxide dismutase in laboratory rats (Kim et al., 2005).
Fruits and vegetables from earlier studies (Verghese, 2000; Williams et al., 2007; Boateng et al., 2007) have been proven to induce the activity of critical hepatic enzymes (GST) and antioxidative enzymes (SOD and CAT) which may reduce oxidative stress. We observed a correlation between enzyme activity and the incidence of ACF. A study conducted to investigate whether diets containing cruciferous vegetables such as broccoli, watercress, green cabbage and red cabbage offered protection against ACF formation in rats, indicated that cabbage increased hepatic GST activity in rats (Arikawa and Gallaher, 2008). Rats fed cabbage also had lower incidence of ACF. In the present study, rats fed bitter melon had higher GST activity and lower ACF incidence. Previous experiments in our laboratory have shown a similar trend. Research show that diets rich in protein could increase oxidative stress (Gu et al., 2008) hence leading to chronic diseases like colon cancer. In a recent study, mice fed high protein diets (60% casein or soy protein) were observed to have low catalase and superoxide dismutase activities (Gu et al., 2008). Proving that a healthy diet is one which consists of a good balance of fruits, vegetable and protein.
Phenolics may inhibit cancer through mechanisms such as antioxidant effects, activation of detoxifying enzymes and increasing cell to cell communication (Malin et al., 2003). In the present study it was observed that bitter melon had phenolic content, indicating that it may play a role in the cancer prevention as the results from the animal study showed. Total phenolic content of bitter melon (34.31 mg/GAE/g fresh fruit) is lower compared to blueberry (190.3 mg/100 g for f.wt.) (Wang et al., 2008b), onion, asparagus and tomatoes (53, 44 and 35 mg of CAE 100 g-1 of FW, respectively), however in comparison to French beans, lettuce and cucumber (34, 32 and 20 mg of CAE 100 g-1 of FW, respectively) (Keevers et al., 2007) bitter melon’s total phenolic content was observed to be higher. These results were expressed in milligrams of Chlorogenic Acid Equivalents (CAE) per 100 g of Fresh Weight (FW). Free radicals are unstable molecules which destabilize surrounding molecules by obtaining electrons from them. Free radicals in food and in the human body can be harmful, hence natural sources of antioxidants is important in the prevention of the damage they cause (Yang et al., 2009). Activities of free radicals can lead to several chronic diseases such as cancer. The present study showed that bitter melon had some level of free radical scavenging activity (DPPH). This indicates that bitter melon may be a natural source of antioxidants. Free radical scavenging activity of bitter melon (0.647 mg mL-1) is higher when compared to vegetables such as egg plant (0.228 mg mL-1) (Nisha et al., 2009), however its ability to scavenge free radicals is made evident by results from the present research. The 0.647 mg mL-1 or 2.99% value indicates the level at which bitter melon scavenged the free radical, 1,1-Diphenyl-2-picryl-hydrazyl. Reduction of ACF by bitter melon in the animal study could have been due to its antioxidative potential. Consumption of fruits and vegetable is encouraged due to their antioxidant activities (Lohachoompol et al., 2004). Bitter melon in the present study showed ferric reducing antioxidant power activity (FRAP). This supports the hypothesis that bitter melon may have antioxidant potential. FRAP assay measures alteration in absorbance, brought about by the formation of blue colored ferrous-tripyridyltriazine from colorless oxidized ferric by the action of electron donating antioxidants (Hazem et al., 2008). In a study conducted aimed at investigating the ability of red cabbage to ameliorate diabetes in rats, FRAP activity was found to be low in kidney due to Reactive Oxygen Species (ROS) activity (Hazem et al., 2008). However, bitter melon in the present study seems to have increased antioxidant activity in rats.
Based on the results of the study it can be deduced that bitter melon may have antiproliferative and antioxidative properties. Bitter melon significantly reduced the incidence of ACF, reduced crypt multiplicity as well as total crypts. Hepatic and antioxidant enzymes were increased in rats fed bitter melon. With colon cancer being the second leading cause of cancer deaths, cutting edge research needs to be conducted to stamp out the staggering statistics of colon cancer cases. Bitter melon proves to be a vegetable which possesses the medicinal properties to prevent colon cancer as results from this study has made obvious. Further endpoint (long term) studies needs to be conducted to determine the chemopreventive effects of bitter melon consumption over a longer period.
ACKNOWLEDGMENT
The funding for this research was provided by The Alabama Experimental Research Station, Normal, AL 35762.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Arikawa, A.Y. and D.D. Gallaher, 2008. Cruciferous vegetables reduce morphological markers of colon cancer risk in dimethylhydrazine-treated rats. J. Nutr., 138: 526-532.
PubMed - Basch, E., S. Gabardi and C. Ulbricht, 2003. Bitter melon (Momordica charantia): A review of efficacy and safety. Am. J. Health Syst. Pharm., 60: 356-359.
PubMedDirect Link - Brand-Williams, W., M.E. Cuvelier and C. Berset, 1995. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol., 28: 25-30.
CrossRefDirect Link - Bird, R., 1987. Observation and quantification of aberrant crypt foci in murine colon treated with colon carcinogen. Cancer Lett., 37: 147-151.
PubMed - Boateng, J., M. Verghese, L. Shackelford, L.T. Walker and J. Khatiwada et al., 2007. Selected fruits reduce azoxymethane (AOM)-induced aberrant crypt foci (ACF) in fisher 344 male rats. Food Chem. Toxicol., 45: 725-732.
CrossRefPubMedDirect Link - Fridovich, I., 1989. Superoxide dismutase. An adaption to paramagnetic gas. J. Biol. Chem., 254: 7761-7764.
PubMed - Gu, C., Y. Shi and G. Le, 2008. Effects of dietary protein levels and origin on the redox status in the digestive tract of mice. Int. J. Mol. Sci., 9: 464-475.
PubMed - Grover, J.K. and S.P. Yadav, 2004. Pharmacological actions and potential uses of Momordica charantia: A review. J. Ethnopharm., 93: 123-132.
CrossRefPubMedDirect Link - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Kevers, C., M. Falkowski, J. Tabart, J.O. Defraigne, J. Dommes and J. Pincemail, 2007. Evolution of antioxidant capacity during storage of selected fruits and vegetables. J. Agric. Food Chem., 55: 8596-8605.
CrossRefPubMedDirect Link - Kim, N.Y., M.K. Lee, M.J. Park, S.J. Kim and H.J. Park et al., 2005. Momordin Ic and oleanolic acid from Kochiae Fructus reduce carbon tetrachloride-induced hepatotoxicity in rats. J. Med. Food, 8: 177-183.
CrossRefPubMedDirect Link - Kim, S.J., E. Ju, D. Lee and H. Hwang, 2002. Induction of apoptosis by momordin 1 in promyelocytic leukemia (HL-60) cells. Anticancer Res., 22: 1885-1889.
Direct Link - Lako, J., V.C. Trenerry, M. Wahlqvist, N. Wattanapenpaiboon, S. Sotheeswaran and R. Premier, 2007. Phytochemical flavonols, carotenoids and the antioxidant properties of a wide selection of Fijian fruit, vegetables and other readily available foods. Food Chem., 101: 1727-1741.
CrossRefDirect Link - Leamon, C. and P. Low, 1992. Cytotoxicity of momordin-folate conjugates in cultured human cells. J. Biol. Chem., 267: 24966-24971.
PubMed - Nisha, P., P. Abdul Nazar and P. Jayamurthy, 2009. A comparative study on antioxidant activities of different varieties of Solanum melongena. Food Chem. Toxicol., 47: 2640-2644.
CrossRef - Ng, T.B., W.Y. Chan and H.W. Yeung, 1992. Proteins with abortifacient, ribosome inactivating, immunomodulatory, antitumor and anti-AIDS activities from Cucurbitaceae plants. Gen. Pharmacol., 23: 579-590.
CrossRef - Ogiwara, T., K. Satoh, Y. Kadoma, Y. Murakami and S. Unten et al., 2002. Radical scavenging activity and cytotoxicity of ferulic acid. Anticancer Res., 22: 2711-2717.
Direct Link - Ochiai, M., H. Nakagama, M. Watanabe, Y. Ishiguro, T. Sugimura and M. Nagao, 1996. Efficient method for rapid induction of aberrant crypt foci in rats with 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine. Jpn. J. Cancer Res., 87: 1029-1033.
PubMed - Parkin, D.M., F. Bray, J. Ferlay and P. Pisani, 2005. Global cancer statistics, 2002. CA: Cancer J. Clin., 55: 74-108.
CrossRefPubMedDirect Link - Pretlow, T.P., B.J. Barrow, W.S. Ashton, M.A. Riordan, T.G. Pretlow, J.A. Jurcisek and T.A. Stellato, 1991. Aberrant crypt: putative preneoplastic foci in human colonic mucosa. Cancer Res., 51: 1564-1567.
PubMed - Riboli, E. and T. Norat, 2003. Epidemiologic evidence of the protective effect of fruit and vegetables on cancer risk. Am. J. Clin. Nutr., 78: 559S-569S.
Direct Link - Sato, K., K. Ito, H. Kohara, Y. Yamaguchi, K. Adachi and H. Endo, 1992. Negative regulation of catalase gene expression in hepatoma cells. Mol. Cell. Biol., 12: 2525-2533.
PubMed - Takahashi, M. and K. Wakabayashi, 2004. Gene mutations and altered gene expression in azoxymethane-induced colon carcinogenesis in rodents. Cancer Sci., 95: 475-480.
PubMed - Wang, S., C. Chen, W. Sciarappa, C. Wang and M. Camp, 2008. Fruit quality, antioxidant capacity and flavonoid content of organically and conventionally grown blueberries. J. Agric. Food Chem., 56: 5788-5796.
CrossRefPubMedDirect Link - Wang, L., M. Wang, Q. Li, T. Cai and W. Jiang, 2008. Partial properties of an aspartic in bitter gourd (Momordica charantia L.) fruit and its activation by heating. Food Chem., 108: 496-502.
CrossRef - Williams, D., M. Verghese, L.T. Walker, J. Boateng, L. Shackelford and C.B. Chawan, 2007. Flax seed oil and flax seed meal reduce the formation of Aberrant Crypt Foci (ACF) in azoxymethane-induced colon cancer in Fisher 344 male rats. Food Chem. Toxicol., 45: 153-159.
PubMedDirect Link - Yamada, Y. and H. Mori, 2003. Pre-cancerous lesions for colorectal cancers in rodents: A new concept. Carcinogenesis, 24: 1015-1019.
PubMed - Zampino, M., R. Labianca, G. Beretta, E. Magni and G. Gatta et al., 2008. Rectal cancer. Crit. Rev. Oncol. Hematol., 70: 160-182.
PubMed - Zielinska, M., A. Kostrzewa, E. Ignatowicz and J. Budzianowski, 2001. The flavoniods, quercetin and isorhamnetin 3-O-acylglucosides diminish neutrophil oxidative metabolism and lipid peroxidation. Acta. Biochimica. Polonica., 48: 183-189.
PubMed - Reeves, P.G., F.H. Nielsen and G.C. Fahey Jr., 1993. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr., 123: 1939-1951.
CrossRefPubMedDirect Link - Benzie, I.F.F. and J.J. Strain, 1996. The ferric reducing ability of plasma (FRAP) as a measure of "antioxidant power": The FRAP assay. Anal. Biochem., 239: 70-76.
CrossRefPubMedDirect Link