
Srinivas Chava
Department of Genetics, Bhagawan Mahavir Hospital and Research Centre, AC Guards, Hyderabad, India
M.D. Kaliq
Department of Genetics, Vasavi Medical and Research Centre, Lakdi ka Pool, Hyderabad, India
Pasupuleti Nagarjuna
Department of Molecular Biology, Kamineni Life Sciences, Moula-Ali, Hyderabad, India
Madhavi Latha
Department of Genetics, Vasavi Medical and Research Centre, Lakdi ka Pool, Hyderabad, India
V. Sireesha
Department of Genetics, Vasavi Medical and Research Centre, Lakdi ka Pool, Hyderabad, India
Preetha Shetty
Department of Genetics, Vasavi Medical and Research Centre, Lakdi ka Pool, Hyderabad, India
Ajith Kumar
Department of Gastroenterology, Nizam�s Institute of Medical Sciences, Hyderabad, India
Vishnu Priya
Department of Genetics, Osmania University, Hyderabad, India
Y.R. Ahuja
Department of Genetics, Vasavi Medical and Research Centre, Lakdi ka Pool, Hyderabad, India
Q. Hasan
Department of Genetics, Bhagawan Mahavir Hospital and Research Centre, AC Guards, Hyderabad, India
International Journal of Cancer Research
Year: 2011 | Volume: 7 | Issue: 3 | Page No.: 233-243







ABSTRACT
Among three epigenetic mechanisms, DNA methylation is a distinct and crucial mechanism to regulate a variety of genes in tissue specific manner in pathophysiology of cancer. Methylation status of four selected genes which include tumor suppressor genes like p53 and FHIT as well as oncogenes, Aurora-A and IGF2 was assessed to find the association with esophageal cancer by performing methylation specific restriction assay in both blood and tissue of patients who had undergone diagnostic endoscopy and results were correlated with the exogenous factors like tobacco, alcohol, hot beverages and diet. Individuals with all four types of exposure were found in a higher percentage in the cancer group when compared to normals and esophagitis, however, those exposed to tobacco and alcohol were significantly more in cancer (p<0.05). The methylation status of p53, FHIT and IGF2 did not show difference between cancer and normal samples whereas Aurora (p = 0.0002). But methylation was increased in p53, FHIT and Aurora-A gene and decreased in IGF2 in cancer tissue if the individuals were exposed to tobacco and alcohol. MDR analysis indicated that the four genes evaluated were acting independently whereas there was a stronger interaction between non vegetarian diet and hot beverages as well as tobacco and alcohol for the development of esophageal cancer. In conclusion it can be stated that exogenous factors affect the methylation status of genes in tissues which may act as an early step in esophageal carcinogenesis.
PDF Abstract XML References Citation
Received: May 10, 2011;
Accepted: July 26, 2011;
Published: August 13, 2011
How to cite this article
Srinivas Chava, M.D. Kaliq, Pasupuleti Nagarjuna, Madhavi Latha, V. Sireesha, Preetha Shetty, Ajith Kumar, Vishnu Priya, Y.R. Ahuja and Q. Hasan, 2011. Epigenetics and Esophageal Cancer: Role of Altered Methylation in Specific Genes. International Journal of Cancer Research, 7: 233-243.
DOI: 10.3923/ijcr.2011.233.243
URL: https://scialert.net/abstract/?doi=ijcr.2011.233.243
DOI: 10.3923/ijcr.2011.233.243
URL: https://scialert.net/abstract/?doi=ijcr.2011.233.243
INTRODUCTION
The development of cancerous lesions is caused by the acquisition of a series of changes in the DNA that give the cell a selective growth advantage. These changes can be genetic, (Chromosomal aberrations and mutations) analyzing cell culture and amplified DNA fragments provide genetic relatedness among chromosomes and genomic DNA (Ibrahim et al., 2010) or epigenetic (DNA methylation and histone acetylation). Methylation of DNA occurs predominantly at cytosines within CpG clusters which occur in the genome and tend to be associated with the promoter and exonic regions of functional genes (Takai and Jones, 2002). Methylation status of CpG dinucleotides are maintained through cell division by DNA methyltransferases, however, in cancer it is known to be altered in a specific pattern.
Esophageal cancer incidence is on the rise globally and this has been attributed to both genetic and exogenous factors (Darvishi et al., 2007). The absolute risk of esophageal cancer is not known but Gastroesophageal Reflux Disease (GERD) is considered as a relative risk factor (Rubenstein et al., 2011). The disease is one of the 10 common ones world wide and is the third one among common GIS involved malignancies (Semnani et al., 2005). The current challenges in the management of esophageal cancer are to obtain a better understanding of the underlying molecular biological alterations and exogenous risk factors to provide prevention, early diagnosis and new management/therapeutic options.
The present study evaluated the methylation status of four selected genes known to regulate different pathways relevant in the etiopathogenesis of esophageal malignancy. Hyper methylation within the promoter-associated CpG islands of suppressor genes could be particularly important, especially when this methylation affects tumor suppressor genes (Ibrahim, 2010). (1) p53 a commonly mutated tumor suppressor known to play an important role in several cancers including esophageal cancer (Mir et al., 2005). p53 promotor methylation status has been reported to down regulate its activity in vitro (Schroeder and Mass, 1997). To the best of our knowledge there are no reports which have evaluated p53 methylation status in esophageal tissues. (2) Fragile Histidine Traid (FHIT) exhibits loss of heterozygosity, abberant transcripts and reduced expression in esophageal cancer in a few reports (Lee et al., 2006; Vasavi et al., 2010). Hence association between FHIT methylation and exogenous factors in esophageal tissue is warranted. (3) Aurora A is a serine/threonine protein kinase and has the potential to play an important role in chromosome assembly and segregation in mitosis, its dysfunction may result in aneuploidy observed in esophageal cancer (Oliver et al., 2008; Vasavi et al., 2007). Altered expression of Aurora A gene has been found in several types of human tumors, including esophageal squamous cell carcinoma (Ouchi et al., 2004; Tong et al., 2004). Methylation studies of Aurora-A gene are less and there are none in esophageal cancer as indicated by our literature survey. (4) IGF2 an imprinted mitogen has demonstrated Loss of imprinting by epigenetic modifications in Wilm’s tumor and other cancers; however, there are few studies which have assessed its role in esophageal pathologies especially its methylation status (Michael et al., 1999; Xu et al., 2006).
In present study Methylation Specific Restriction Assay (MSRA) has been carried out to establish the methylation status of the above mentioned four genes in blood and esophageal tissue of the same individual and the results were correlated. Exogenous factors which are commonly considered as environmental insults in the pathogenesis of esophageal cancer were assessed for their association with esophageal pathologies, as well as, their role in regulating the selected genes by altering methylation.
MATERIALS AND METHODS
Study population: The present study included 180 consecutive patients suffering from upper gastrointestinal tract symptoms, who were referred for endoscopy to two separate Gastroenterology units during the period of 2009-2010. The patients were later categorized on the basis of endoscopy and histology reports into 3 groups (1) esophageal cancer (2) esophagitis and (3) normals.
The data about the patient’s personal history which included age, gender, diet, tobacco usage (smoking and chewing), alcohol intake and hot beverages (tea/coffee consumption), along with clinical details were collected at the time of sampling. Present study was approved by the institutional ethical committee and informed consent was obtained.
Sample collection: The 2 mL peripheral blood sample along with biopsy tissue sample was available for 74 cases and used for methylation analysis.
DNA extraction:
From tissue: Fresh tissue biopsy samples were collected at the time of endoscopy and transported to the lab in saline solution for isolating DNA by the method of Vasavi et al. (2006). Briefly biopsy samples were treated for 10 min with TKM1 buffer (10 mM Tris HCl, 10 mM KCl, 10 mM MgCl2, 2 mM EDTA along with 100 μL of Triton X) in order to lyse any red blood cells. These were centrifuged at 3000 rpm for 5 min. Supernatant was discarded and 260 μL of TKM2 buffer (10 mM Tris HCl, 10 mM KCl, 10 mM MgCl2 and 2 mM EDTA, 0.4 M NaCl) was added to the nuclear pellet. Along with this, 60 μL of 10% SDS and 30 μL proteinase K (1 mg mL-1) were added. At the end of 30 min incubation, 70 μL of 6M NaCl was added to precipitate proteins. The solution was spun down and supernatant was transferred into isopropanol to extract DNA. The DNA pellet was washed in 70% ethanol and dried. The DNA was dissolved in 40 μL of TE buffer. This was incubated at room temperature for overnight and stored in -20°C until further use.
From blood: DNA was isolated from 300 μL of whole blood by the salting out method which is routinely followed in our lab (Alluri et al., 2005). It was similar to the method described above except that TKM1 buffer step was repeated until all the RBC’s were removed.
Methylation specific restriction assay (MSRA): Equal amounts of amplifiable DNA from 74 paired blood and tissue samples were subjected to digestion using methyl sensitive restriction endonuclease HpaII (20 units of HpaII and 0.5 μg of DNA in 10 μL reaction; MBI Fermentas, USA). In parallel a mock digest was set up with digestion buffer and template DNA without the restriction enzyme. All the samples were incubated for 14 h at 37°C. A three step PCR by the method reported from our group (Vasavi et al., 2006) was carried out with digested and undigested DNA with the appropriate primers. The primer sequence of each gene, annealing temperatures and the expected band size are listed in Table 1. Thirty five cycles were performed in a thermal cycler with initial denaturation 94°C for 5 min, denaturation at 94 for 30 sec, annealing at 55, 60°C for 30 sec, 72°C for 45 sec and a final extension at 72°C for 5 min. PCR products were checked on ethidium bromide 2% agarose gels and bands were analyzed on a UVI gel documentation system.
Methylation assay analysis: HpaII enzyme recognizes C/CGG sites and cleaves the DNA in these regions when they are unmethylated and does not give the PCR product. However, methylated sites give PCR products of expected size (Fig. 1).
| Table 1: | Primers used for carrying out Methylation specific PCR (MSP) of the four genes selected for the study |
 | |
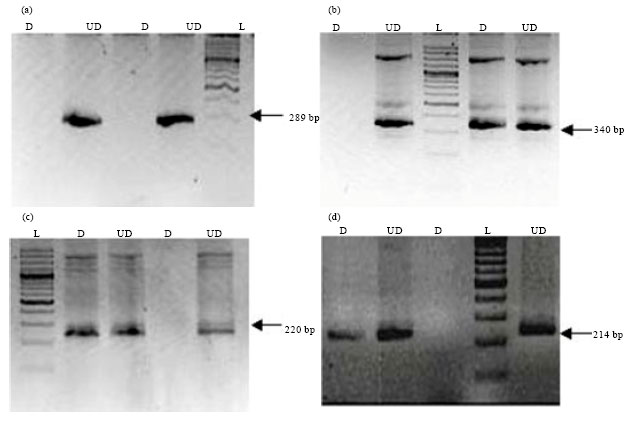 | |
| Fig. 1 (a-d): | Ethidium bromide stained agarose gels (Invreverse image) showing methylation status of the four genes (a): p53, (b): FHIT, (c): Aurora-A and (d): IGF2. Each image shows the results of two representative samples and DNA size ladder. D: Digested and UD: UnDigested. The presence of specific size band in digested sample indicates methylation while the band in undigested serves as an internal control |
Statistical analysis: Data was presented as percentages and Mean±SD. Chi square was used for comparisons and significance by medical software. MDR analysis was carried out to assess interactions between genes and exogenous factors using MDR software.
RESULTS
The study population consisted of 74/180 (41%) females and 106/180 (59%) males. The age group was 30-80 years with a mean age of 54.61±2.52 years and median age of 63 years. Out of 180 cases evaluated 45% had esophageal cancer, 11% esophagitis and remaining 46% were normals based on endoscopic observation and histopathological reports.
We had both blood and tissue samples available from 74 cases and these were processed for methylation studies. Methylated CpG promoter status was considered for analysis in p53, FHIT and Aurora-A genes while loss of methylation was taken into consideration for IGF2, since it is a known imprinted gene. The over all methylation percentage of the four genes, p53, FHIT, Aurora-A and IGF2 in normal esophageal tissue was 83, 21, 31 and 66%, respectively and 21, 24, 7 and 76%, respectively in paired blood samples suggesting that methylation of these genes is independent of each other (Table 2). To our surprise the methylation status of p53, FHIT and IGF2 was not statistically different between normal and cancer tissue samples (Table 2) while that of Aurora-A was significantly different (p = 0.002). In the esophagitis group also the methylation status of p53, FHIT and IGF2 was not significantly different from normal samples, where as only Aurora A showed higher methylation (76%) compared to normal samples (31%) (Table 2).
| Table 2: | Methylation status of three genes (p53, FHIT and Aurora-A) and hypomethylation of IGF2 in blood and tissue samples studied from cases categorized into 3 groups |
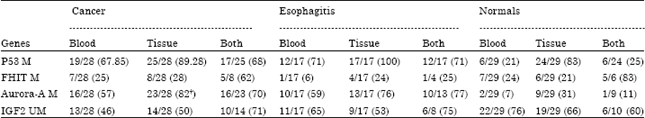 | |
| M: Methylated, UM: Unmethylated, †p = 0.0002. Values in brackets are percentage | |
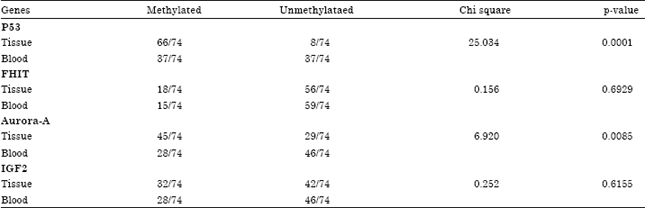
| Table 3: | Statistical analysis of MSR assay carried out in 74 paired blood and tissue samples showing that there is a tissue specific methylation of p53 and Aurora-A |
 | |
 | |
| Fig. 2: | Exogenous factors interactions by using MDR software |
The chi square test showed a significant difference (p<0.005) between p53 and Aurora-A methylation status in tissue when compared to blood indicating that these genes are methylated in a tissue specific manner (Table 3). However, the methylation status of FHIT and IGF2 genes in blood and tissue was not statistically different (Table 3).
| Table 4: | Percentages of individuals in three categories with exposure to exogenous factors |
 | |
| *Diet: Non-Vegetarian, **Tobacco: Both Chewers and Smokers, ***Hot Beverages: Tea/Coffee | |
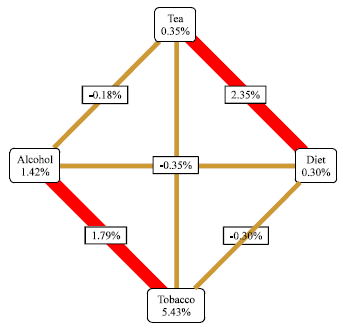
Interaction between four exogenous factors in the pathogenesis of esophageal cancer was assessed by MDR analysis (Fig. 2). Based on this the epigenetic interaction between aurora and p53 promotor methylation and IGF2 hypomethylation appears to confer a high risk of developing esophageal cancer irrespective of the methylation status of FHIT gene (Fig. 3a).
Since esophageal tissue is constantly exposed to exogenous risk factors, individuals were categorized as non vegetarians, tobacco users which included both chewers and smokers while consumers of hot beverages which included both tea and coffee and alcohol. All four exogenous factors (non-vegetarian diet, tobacco usage, alcohol and hot beverage consumption) were in a higher percentage in cancer group when compared to those in normal and esophagitis groups (Table 4), however, this difference was statistically significant only for tobacco (p = 0.0001) and alcohol (p = 0.0001).
Nonvegetarian diet, alcohol consumption and tobacco interaction confers a high risk of esophageal cancer while hot beverages (mostly tea in our study) reduced this risk (Fig. 3b).
We also looked at the interactive effect that the four studied exogenous factors had on the methylation status of the selected genes. In the case of p53 promoter methylation all four factors appear to be playing a role, however, interaction between non vegetarian diet, tobacco and tea effects methylation of this gene significantly (Fig. 3c).
FHIT gene methyaltion appears to be least effected significantly in cancer by any of the four exogenous agents (Fig. 3d) while Aurora-A gene methylation was effected by tobacco, alcohol and tea while the interactive effect of nonveg diet, alcohol and tobacco on methylation was reduced by tea (Fig. 3e). IGF2 hypomethylation was not regulated by exogenous factors (Fig. 3f) indicating that they may be important factors for methylation but not for hypomethylation.
The effect of exogenous factors on the methylation status of the individual genes was evaluated which showed that tobacco and alcohol increased methylation of p53, FHIT and Aurora-A genes and hypomethylation of IGF2 in cancer when compared with normal samples (Fig. 4a). While non vegetarian diet and hot beverages did not show difference in percentage of methylation of the four genes in cancer when compared to normal samples (Fig. 4b).
However, exogenous factors like non veg diet and hot beverages which did not show any association with cancer individually, exhibited an interaction which is stronger than the interaction seen between tobacco and alcohol (Fig. 2); however this conferred low risk and can be considered as protective.
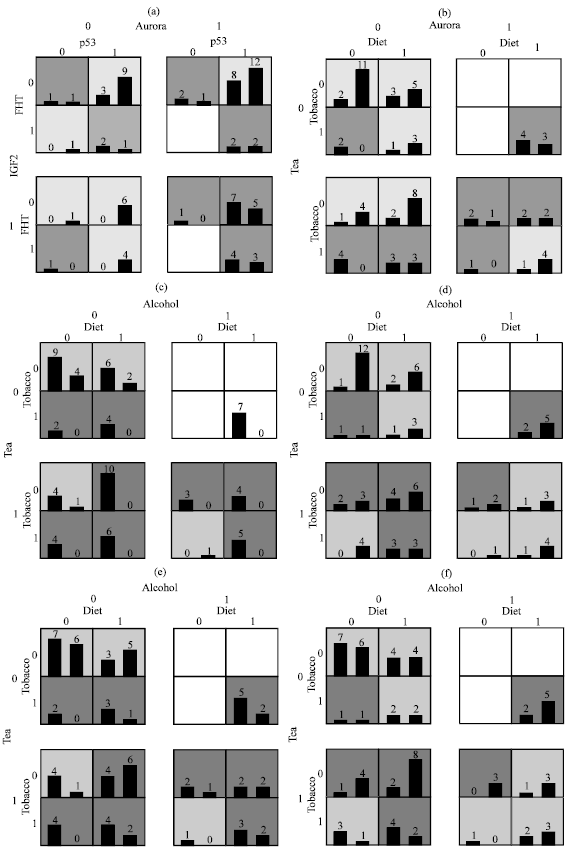 | |
| Fig. 3 (a-f): | Bars represent hypothetical distributions of cases (left) and controls (right) with each multifactor combination. Dark shaded cells represent high-risk combinations while light shaded cells represent low-risk combinations. No shading or white cells represent combinations for which no data was observed |
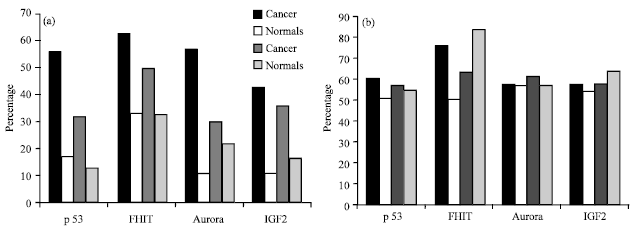 | |
| Fig. 4 (a-b): | (a) Methylation percentages of Tobacco and Alcohol usage of p53, FHIT and Aurora-A and loss of methylation of IGF2. (The 1st pair of bars is cancer and normal samples exposed to tobacco and 2nd pair to alcohol), (b) Methylation percentages of Diet and Hot beverages of p53, FHIT and Aurora-A and loss of methylation of IGF2. (The 1st pair of bars is cancer and normal samples exposed to diet and 2nd pair to hotbeverages) |
DISCUSSION
Esophageal cancer is one of the leading causes of mortality and morbidity. More than 30% of the adult population worldwide exhibits upper Gastrointestinal (GI) tract disorders associated with symptoms such as regurgitation, heartburn and dysphagia, warranting an endoscopic evaluation (Vasavi et al., 2007). These symptoms are some times considered to be caused due to exogenous risk factors like diet, tobacco usage (both smoking and chewing), hot beverage and alcohol consumption. However, only a small percentage of these patients exhibit premalignant or malignant pathologies. Different esophageal cancer incident rates among major ethnic groups may rise the old theory about possibly of genetic propensity and/or life style characteristics that predispose of a specific ethnic group (for example Googlans) to esophageal cancer (Semnani et al., 2006).
The present study evaluated 180 cases requiring endoscopy due to upper GI tract discomfort/symptoms, a majority of them were men, the ratio between males and females was not the expected 3:1 (Wabinga et al., 2000) and this may be because the cases belonged to individuals warranting an endoscopy but not EC. 87% of these were non vegetarians which is significantly higher than the established 57-58% non vegetarians in the Indian population (UN food and agriculture). This observation suggests that non vegetarian diet plays a role in the upper GI tract discomfort and our results are similar to a study from Bombay (Ganesh et al., 2009). India is a predominantly hot beverage drinking country with tea being more popular in north India and coffee in south India, hence it was not surprising that 71% of our patients have hot beverages (Table 4). 37% of the cases were alcohol consumers (n = 66) out of which 7 were females. 49% were exposed to tobacco and this include both smoking/chewing (n = 88), none of the women were smokers but seven of them were tobacco chewers with only 5 individuals who had predominantly coffee (Table 4). The percentage of cases using tobacco and alcohol were significantly higher in the cancer group when compared to endoscopically confirmed esophagitis and normals (p<0.05). This is similar to earlier reports of Ganesh et al. (2009). This is the first report to the best of our knowledge which has used MDR analysis to evaluate interaction between exogenous factors known as risk factors for EC.
The strength of our study is that the methylation status of the four genes, p53, FHIT, Aurora-A and IGF2 were evaluated in paired tissue and blood samples collected at the time of endoscopy and biopsy collected was confirmed by histopathology. The four genes selected were known to independently regulate different pathways and have evidence of playing a role in the pathogenesis of esophageal cancer.
P53 is an important tumor suppressor gene which has been seen to be both mutated and methylated in cancer to promote tumorogenesis (Kang et al., 2001). Our results showed that p53 was methylated in 83% of normal samples as compared to 89% of cancer samples suggesting that either p53 methylation is not relevant to esophageal cancer pathophysiology or that the CpG cluster evaluated may not be appropriate for regulating p53 gene activity. There are no studies of p53 methylation in EC and this is surprising the universal role of p53 in cancer pathophysiology.
Earlier MSI studies from our group reported that FHIT marker was associated with pre- cancerous and cancerous lesions of the esophagus (Vasavi et al., 2010). In the present study FHIT promoter methylation was seen in 29% of cancer tissue which was in the range of 15-45% reported by earlier workers (Lee et al., 2006). However, 21% of normal tissues also showed FHIT promoter methylation this may be due to exogenous factors as, indicated by earlier studies. Lee et al. (2006) showed that altered methylation/expression of FHIT have been associated with exposure to tobacco.
Aurora-A gene polymorphisms have been associated with esophageal cancer both by other workers and our group (Miao et al., 2004; Chava et al., 2011); however, to the best of our knowledge this is the first paper indicating that promoter methylation is important for esophageal cancer etiology, since 82% of cancer tissues were found to be methylated in comparison to 31% normal samples (p = 0.0002).
Increased expression of IGF2 due to loss of imprinting caused by altered methylation has been seen in several cancers including that of esophagus (Preetha et al., 2010). In the present study methylation status of DMR 2 which is known to be significantly altered in breast cancer based on our earlier study (Preetha et al., 2010) was evaluated but did not show any significant loss of methylation in esophageal cancer as compared to controls.
It appears that the exogenous factors studied are relevant for methylation but not for demethylation/hypomethylation. It was surprising that tobacco showed least effect on FHIT methylation as an earlier study by Lee et al. (2006) had indicated that smoking may be an important factor regulating the methylation of this gene. The reason for this difference may be because we have included individuals both tobacco chewers and smokers in the same category.
CONCLUSION
Despite the fact that the sample size is small, the preliminary results from the comprehensive analysis on methylation status of four genes in paired samples from the same patients indicates that these four genes are differently methylated in normal tissue.
In conclusion it can be stated that exogenous factors affect the methylation status of genes in tissues which may act as an early step in esophageal carcinogenesis.
REFERENCES
- Alluri, R.V., V. Mohan, S. Komandur, K. Chawda, J.R. Chaudhuri and Q. Hasan, 2005. MTHFR C677T gene mutation as a risk factor for arterial stroke: A hospital based study. Eur. J. Neurol., 12: 40-44.
CrossRef - Darvishi, K., S. Sharma, A.K. Bhat, E. Rai and R.N. Bamezai, 2007. Mitochondrial DNA G10398A polymorphism imparts maternal Haplogroup N a risk for breast and esophageal cancer. Cancer Lett., 249: 249-255.
Direct Link - Ganesh, B., S.D. Talole and R. Dikshit, 2009. Tobacco, alcohol and tea drinking as risk factors for esophageal cancer: A case control study from Mumbai, India. Cancer Epidemiol., 33: 431-434.
CrossRef - Kang, J.H., S.J. Kim, D.Y. Noh, I.A. Park, K.J. Choe, O.J. Yoo and H.S. Kang, 2001. Methylation in the p53 promoter is a supplementary route to breast carcinogenesis: Correlation betweenCpG methylation in the p53 promoter and the mutation of the p53 gene in the progression from ductal carcinoma in situ to invasive ductal carcinoma. Lab. Invest., 81: 573- 579.
Direct Link - Lee, E.J., B.B. Lee, J.W. Kim, Y.M. Shim and I. Hoseok et al., 2006. Aberrant methylation of Fragile Histidine Triad gene is associated with poor prognosis in early stage esophageal squamous cell carcinoma. Eur. J. Cancer, 42: 972-980.
Direct Link - Miao, X., T. Sun, Y. Wang, X. Zhang, W. Tan and D. Lin, 2004. Functional STK15 Phe31Ile polymorphism is associated with the occurrence and advanced disease status of esophageal squamous cell carcinoma. Cancer Res., 64: 2680-2683.
CrossRefDirect Link - Michael, J.S., T. Takanobu, J. Agnes, K. Natalie and E.R. Anthony, 1999. Relaxation of IGF2 imprinting in Wilms tumours associated with specific changes in IGF2 methylation. Oncogene, 18: 7527-7534.
Direct Link - Mir, M.M., N.A. Dar, S. Gochhait, A.S. Zargar, A.G. Ahangar and R.N.K. Bamezai, 2005. P53 Mutation profile of squamous cell carcinomas of the esophagus in Kashmir (India): A high-incidence area. Int. J. Cancer, 116: 62-68.
Direct Link - Oliver, G., H. Jim, C.M. Philip, R.P. Phillip, N.L. Jr. Primo and R.G. David, 2008. Aurora kinases as anticancer drug targets. Clin. Cancer Res., 14: 1639-1639.
CrossRef - Ouchi, M., N. Fujiuchi, K. Sasai, H. Katayam and Y.A. Minamishima et al., 2004. BRCA1 phosphorylation by Aurora-A in the regulation of G2 to M transition. J. Biol. Chem., 279: 19643-19648.
CrossRef - Preetha, J.S., S. Movva, N. Pasupuleti, B. Vedicherlla and K.K. Vattam et al., 2010. Regulation of IGF2 transcript and protein expression by altered methylation in breast cancer. J. Cancer Res. Clin. Oncol., 137: 339-345.
CrossRef - Schroeder, M. and M.J. Mass, 1997. CpG methylation inactivates the transcriptional activity of the promoter of the human p53 tumor suppressor gene. Biochem. Biophys. Res. Commun., 235: 403-406.
PubMed - Semnani, S., N. Abdollahi, K. Kalavi and R. Azarhoosh, 2005. Esophageal cancer in an iranian 20 years old young male-A case report. Int. J. Cancer Res., 1: 57-59.
CrossRefDirect Link - Semnani, S., S. Besharat, N. Abdolahi, A. Keshtkar, M. Sakhavi, A. Danesh and K. Kalavi, 2006. Esophageal cancer in Iranian Turkmens: An ethnic disparity concern. J. Medical Sci., 6: 1007-1010.
CrossRefDirect Link - Chava, S., V. Mohan, N. Pasupuleti, M.M. Latha and A.I. Khan et al., 2011. Evaluation of aurora-a gene polymorphism and esophageal cancer risk in a South Indian Population. Genet. Testing Mol. Biomarkers, 15: 185-189.
CrossRefDirect Link - Takai, D. and P.A. Jones, 2002. Comprehensive analysis of CpG islands in human chromosomes 21 and 22. Proc. Natl. Acad. Sci. USA., 99: 3740-3745.
CrossRef - Tong, T., Z. Yali, K. Jainping, D. Lijia and S. Yongmei et al., 2004. Over expression of Aurora-A contributes to malignant development of human esophageal squamous cell carcinoma. Clin. Cancer Res., 10: 7304-7310.
CrossRef - Vasavi, M., V. Kiran, B. Ravishankar, B. Prabhakar, Y.R. Ahuja and Q. Hasan, 2010. Microsatellite instability analysis and its correlation with hMLH1 repair gene hypermethylation status in esophageal pathologies including cancers. Cancer Biomark, 7: 1-10.
CrossRefDirect Link - Vasavi, M., P. Shivani, M.R. Hemakumar, S. Radha, A.J. Rachel, Y.R. Ahuja and Q. Hasan, 2007. Chromosome 11 aneusomy in esophageal cancers and precancerous lesions- an early event in neoplastic transformation: An interphase fluorescence in situ hybridization study from south India. World J. Gastroenterol., 13: 503-508.
Direct Link - Vasavi, M., P. Shivani, G. Kusumanjali, B. Prabhakar and S.B. Ravi et al., 2006. DNA methylation in esophageal diseases including cancer: Special reference to Hmlh1 gene promoter status. Tumori, 92: 155-162.
Direct Link - Wabinga, H.R., D.M. Parkin, F. Wabwire-Mangen and S. Nambooze, 2000. Trends in cancer incidence in Kyadondo County, Uganda, 1960-1997. Br. J. Cancer, 82: 1585-1592.
PubMed - Xu, W., H. Fan, X. He, J. Zhang and W. Xie, 2006. LOI of IGF2 is associated with esophageal cancer and linked to methylation status of IGF2 DMR. J. Exp. Clin. Cancer Res., 25: 543-547.
PubMed - Ibrahim, M.A., N. Saleh, E. Archoukieh, H.W. Al-Obaide, M.M. Al-Obaidi and H.M. Said, 2010. Detection of novel genomic polymorphism in acute lymphoblastic leukemia by random amplified polymorphic DNA analysis. Int. J. Cancer Res., 6: 19-26.
CrossRef - Ibrahim, M.A., 2010. Perspective of DNA methylation in cancer research. Int. J. Cancer Res., 6: 188-201.
CrossRefDirect Link