
K. Nirmala
Food and Drug Toxicology Research Centre, National Institute of Nutrition, Indian Council of Medical Research, Hyderabad-500 007, India
Virendra V. Panpatil
Food and Drug Toxicology Research Centre, National Institute of Nutrition, Indian Council of Medical Research, Hyderabad-500 007, India
A.K. Raja Kumar
Food and Drug Toxicology Research Centre, National Institute of Nutrition, Indian Council of Medical Research, Hyderabad-500 007, India
N. Balakrishna
Department of Statistics, National Institute of Nutrition, Indian Council of Medical Research, Hyderabad-500 007, India
R. Balansky
National Specialized Hospital for Active Treatment of Cancer, Str. Plovdivsko pole 6, Sofia-1756, Bulgaria, India
K. Polasa
Food and Drug Toxicology Research Centre, National Institute of Nutrition, Indian Council of Medical Research, Hyderabad-500 007, India
International Journal of Cancer Research
Year: 2011 | Volume: 7 | Issue: 2 | Page No.: 114-124







ABSTRACT
Epidemiological, in vitro and in vivo studies indicate that a plant-based diet can reduce the risk of cancer and other chronic diseases. The mechanisms through which some of the nutrients present in diet might protect against many diseases like cancer are not clear. Therefore, development of biomarkers suitable for investigating the molecular effects of dietary factors in animal and human studies is of great importance. The purpose of the present study was to determine the in vivo genotoxic effects of benzo(a)pyrene [B(a)P] exposure in tissues of rats and the protective role of turmeric fed through diet. Rats were divided into four groups of twelve animals each and were fed diets containing turmeric at 1, 3, 5% and a control group without turmeric for a period of one month. At the end of the feeding period, half the animals from each group were given B(a)P (5 mg/rat; intra-peritoneal) and after 24 h all the animals were sacrificed and tissues such as liver, lung, kidney and femurs were collected and analyzed using different biomarkers of genotoxicity and oxidative damage. The results of the study showed that turmeric had no genotoxic or cytotoxic effect but had a protective effect against B(a)P toxicity by modulating the lipid peroxidation, urinary 8-hydroxydeoxyguanosine (8-OHdG) and DNA damage. Inhibition of the induction of micronuclei (MN) by turmeric indicates its protective potential against cytogenetic damage.
PDF Abstract XML References Citation
Received: September 09, 2010;
Accepted: December 23, 2010;
Published: April 22, 2011
How to cite this article
K. Nirmala, Virendra V. Panpatil, A.K. Raja Kumar, N. Balakrishna, R. Balansky and K. Polasa, 2011. Turmeric Alleviates Benzo(a)pyrene Induced Genotoxicity in Rats: Micronuclei Formation in Bone Marrow Cells and DNA Damage in Tissues. International Journal of Cancer Research, 7: 114-124.
DOI: 10.3923/ijcr.2011.114.124
URL: https://scialert.net/abstract/?doi=ijcr.2011.114.124
DOI: 10.3923/ijcr.2011.114.124
URL: https://scialert.net/abstract/?doi=ijcr.2011.114.124
INTRODUCTION
Numerous studies have found that chemoprevention can prevent or improve the outcome of a wide variety of cancers. By far, the major source of exposure to naturally occurring chemicals is the diet (Patil et al., 2009). The safety of selected phytochemicals with chemopreventive potential should be validated by the use of biomarkers as an early surrogate endpoint that is pivotal in conducting prospective chemopreventive trials. There have been many studies during the past two decades that have looked for potential naturally occurring cancer chemopreventive agents. Rapid changes in cancer incidence over time around the world suggest that they could be mostly due to environmental rather than genetic factors (Peto, 2001).
Some examples of promising diet-derived compounds are folate, epigallocatechin gallate present in tea, curcumin in turmeric, genistein in soya etc. that have been demonstrated to have tumor-suppressing properties in rodent models of carcinogenesis (Moiseeva and Manson, 2009; Banerjee et al., 2010). Major risk factors for cancer include tobacco chewing, smoking and alcohol consumption. A large number of chemical carcinogens lead to cancer through Reactive Oxygen Species (ROS) causing oxidative stress in the system which induces strand breaks and can modify DNA bases which in turn contribute to mutagenesis and carcinogenesis.
The relationship between diet and cancer has been implicated in several epidemiological studies. Many compounds in the diet exert anticarcinogenic effects in animal models of cancer and progress has been made in defining their biological activities at the molecular level that include excretion of carcinogens, suppression of cox-2 expression, inhibition of mitosis and induction of apoptosis at various stages of cancer (Johnson, 2007). One of the important components of diet is turmeric. It has been used for its medicinal properties for various ailments and through routes of administration, including topical, oral and inhalation. Research on turmeric has been focused on its antioxidant, hepatoprotective, anti-inflammatory, antimicrobial and its ability to immunomodulatory properties system (Jagetia and Aggarwal, 2007).
Curcuminoids constitute 5% of the turmeric rhizome and have antioxidant and anti-inflammatory effects (Itokawa et al., 2008). This study has been conducted to evaluate the in vivo genotoxic effects of B(a)P exposure in tissues and the preventive role of turmeric fed through diet. Measurement of oxidative damage to lipids has been determined which would indicate the tissue damage as a consequence of carcinogen exposure.
MATERIALS AND METHODS
The project was conducted in National Institute of Nutrition (NIN), Hyderabad, India from December 2009 to May 2010.
Animals: Male Wistar NIN (WNIN) rats were obtained from the National Centre for Laboratory Animal Science (NCLAS), located in NIN and housed in the animal facility where the temperature was maintained at 24-25°C with 12 h. dark/light cycle. The experimental protocol was approved by the Institutional Animal Ethics Committee (IAEC) under Committee for Purpose of Control and Supervision on Experiments on Animals (CPCSEA), Ministry of Environment and Forests, Government of India. Turmeric powder was obtained from commercially available standard grade (AGMARK).
Study design: Age (8-9 weeks) and weight (100-150 g) matched inbred male WNIN rats (n=48) were divided into 4 groups and fed stock diet containing turmeric powder at 1, 3, 5% levels and diet without turmeric (control) for a period of one month. Animals had free access to food and water.
At the end of one month feeding, half the animals from each group were given B(a)P (5 mg/rat) intraperitoneally and after 24 h all the animals were euthanised by placing them in a chamber containing carbon dioxide (CO2) and liver, kidney and lungs were collected. Whole blood was collected from the orbital plexus prior to euthanisation and used for the comet assay whereas serum was separated and used for lipid peroxidation estimation.
Urine was collected 24 h prior and after B(a)P injection for the analysis of 8-hydroxydeoxyguanosine (8-OHdG). Tissue samples were also processed for comet assay. Femurs were collected for rodent bone marrow micronucleus test to see the effect of turmeric on B(a)P exposure.
The comet assay: Single cell suspension was made using cell dissociation technique (Hu et al., 2002) and the resulting cell suspension was taken for the Alkaline Comet assay (Singh et al., 1988).
Briefly, cell suspension/whole blood (40 μL) was mixed with 100 μL of 0.5% low melting point agarose (LMPA) and placed on frosted microscope slides that have already been pre-layered with 1% normal melting point agarose (NMPA). After solidification, slides were covered with a third layer IC of 0.5% LMPA (200 μL). Later they were immersed into a lysing solution (1% sodium sarcosinate, 2.5 M NaCl, 100 mM sodium EDTA, 10 mM Tris-HCl (pH-10), 1% Triton X-100 for 1 h at 4°C.
The slides were kept in alkaline electrophoresis and DNA unwinding was allowed for 20 min at room temperature and then were subjected to electrophoresis for further 20 min at 25 V and 300 mA. The slides were rinsed thrice with 0.4 M Tris (pH 7.4) and then dipped in 70% ethanol followed by 100% ethanol for a minute. They were allowed to dry, stained with ethidium bromide and examined using Flourescent Microscope (Olympus) with monochrome CCD camera with grabber and image analysis software (Komet 5.5) that was equipped with an excitation filter of 515-560 nm and a barrier filter of 590 nm.
The extent of DNA damage in the tissues was quantified by the Olive Tail Moment (OTM) and Tail Length (TL). Olive tail moment was calculated by multiplying %DNA tail with tail moment length i.e., measurement from centre of the head to centre of tail. Quantification of the comet images in blood was done by the visual scoring system using an ocular micrometer.
8-hydroxydeoxyguanosine (8-OHdG): 8-OHdG, one of the by product of oxidative DNA damage, is physiologically found and enhanced by chemical carcinogenesis. 8-OHdG formed during the repair of DNA damage in vivo by exonucleases, is excreted in urine without further metabolism.
The quantity of 8-OHdG in unknown sample was determined by comparing its absorbance with that of a known amount of 8-OHdG using a standard curve. Unknown 8-OHdG samples/standards were first added to an 8-OHdG/BSA (bovine serum albumin) conjugate preadsorbed on an EIA plate (Wu et al., 2004). After a brief incubation, anti-8-OHdG monoclonal antibody was added followed by HRP conjugated secondary antibody.
Urine was collected 24 h prior and after B(a)P injection. The urine was centrifuged at 3000 rpm for 10 min. 1.0 mL of supernatant was stored at -80°C until analysis. 8-OHdG levels were measured using oxiselect TM oxidate DNA damage ELISA kit (8-OHdG quantitation, Cell Biolabs, Inc., USA). Absorbance of each well was read at 450 nm by a microplate reader. The measurement range was 0.5-200 ng mLG1. The urinary 8-OHdG was expressed as total amounts excreted in 24 h.
Lipid peroxidation (MDA): Lipid peroxidation in plasma was determined using Aldetect (MA-Specific) Lipid peroxidation Assay kit (AK 171, Biomol Intenational Inc., USA). In this assay, a calibration curve was prepared using MDA standard. The concentration of MDA in an unknown sample was determined from the absorbance of the unknown at 586 nm and the standard curve (Botsoglou et al., 1994).
Rodent bone marrow micronucleus test: The dissected femurs were opened and the bone marrow was gently flushed out using fetal calf serum and made fine colloid using the syringe and centrifuged at 800 rpm for 5 min. The supernatant was poured out and the sediment is over layered with one or two drops of calf serum. The cells were smeared on glass slide and air-dried and then stained successively with May-Gruenwald and Giemsa stain. 2000 polychromatic erythrocytes (PCEs) per animal were scored to determine the frequency of micronucleated polychromatic erythrocytes (MNPCEs) (Salamone and Mavourin, 2005) and the ratio of PCE/NCE in 200 normochromatic erythrocytes (NCEs) were counted.
Microscopic observations were carried out using Leica microscope with plane objective at 100X/1.25 oil mount magnification. The percentage reduction in the frequency of MNPCEs was also calculated (Celikler et al., 2009).
Statistical analysis: Statistical Package for Social Science (SPSS) windows version 15.0 was used for statistical analysis. Mean and SD values were calculated for all the variables and groups. Mean values were compared by one way ANOVA with post hoc test of Least Significant Differences (LSD) among groups. Paired t-test was used for comparison of mean differences of dependent samples for each group. Non-parametric tests of Kruskal wallis-wilcoxn signed rank test was performed whenever the assumptions of parametric tests violated. For the analysis of micronuclei, 2000 PCEs were scored to calculate the MN frequencies and 200 NCE were examined to determine the ratio of PCE to NCE. Differences in the incidence of MNPCE per group and of PCE per 2000 erythrocytes (PCE+NCE) were compared between B(a)P control and B(a)P+turmeric fed groups using the Mann-Whitney U-test (two-tailed).
RESULTS
Difference of comet ratios (y/x i.e., width of the head/length of tail) was observed between the normal and B(a)P treated groups in blood. There was decrease in the extent of DNA damage in the turmeric fed groups compared to B(a)P treated groups (Table 1).
Reduction in the malondialdehyde (MDA) levels was observed in turmeric fed groups compared to control. Significant decrease in the MDA levels was also seen in turmeric fed groups treated with B(a)P compared to B(a)P group without turmeric (Fig. 1).
B(a)P group exhibited significantly higher mean levels of DNA damage in OTM and TL parameters in all the organs namely liver, kidney and lung compared to the control group (Table 2-4). Dose related decrease in DNA migration in response to B(a)P+turmeric fed groups was observed in kidney at 1, 3 and 5% whereas in lung decrease in DNA migration was seen in all the groups treated with B(a)P+turmeric groups compared to B(a)P treated group with respect to OTM. Decrease in the TL was observed at 3 and 5% groups treated with B(a)P compared to the B(a)P group in all the organs (Table 3-4).
| Table 1: | Effect of turmeric on B(a)P induced DNA damage in rat blood |
 | |
| Values are mean±SD of 6 rats/group. Different superscripts are significant, at p<0.05 between B(a)P treated control and B(a)P treated turmeric groups by ANOVA | |
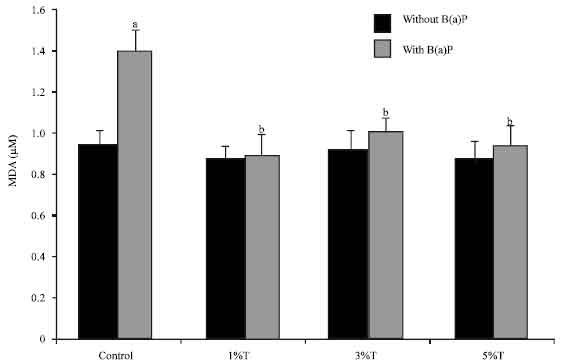 | |
| Fig. 1: | MDA levels in plasma of control and Turmeric (T) fed rats with and without B(a) p-values are Mean±SD of 6 rats/ group. Means with different superscripts differ significantly (p<0.05) analysed by one-way ANOVA |
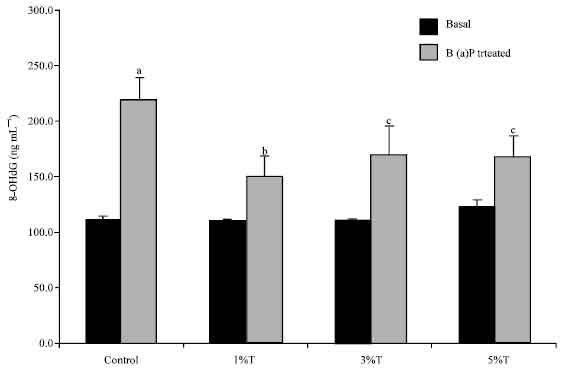 | |
| Fig. 2: | Urinary 8-OHdG levels in B(a)P treated rats fed with turmeric. Each bar represents Mean±SEM of 6 rats/group. Different letters indicate significant differences among the B(a)P+Turmeric groups at p<0.05 analysed by ANOVA with post hoc test of LSD |
| Table 2: | B(a)P induced DNA damage parameters determined by comet assay in liver of rats fed with Turmeric(T) |
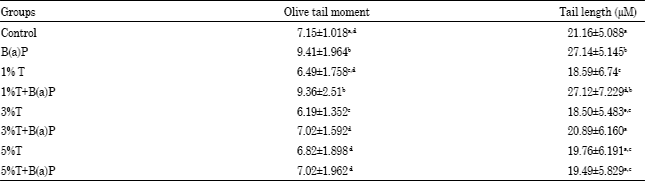 | |
| Values are mean±SD of 6 rats/group. Means in the same column with different superscripts differ significantly (p<0.05) | |
Urinary 8-OHdG levels in the basal samples were similar in the turmeric fed groups and control but were higher in B(a)P group. However in B(a)P+turmeric fed groups significant decrease in the urinary excretion of 8-OHdG was observed at all the levels of turmeric compared to the B(a)P group. Feeding of turmeric inhibited the formation of urinary 8-OHdG (Fig. 2).
| Table 3: | B(a)P induced DNA damage parameters determined by comet assay in kidney of rats fed with Turmeric (T) |
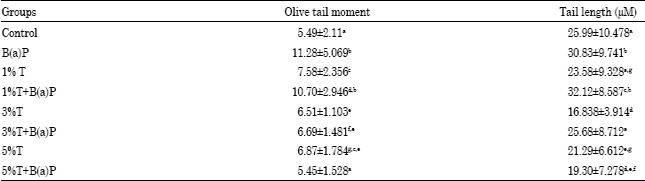 | |
| Values are mean±SD of 6 rats/group. Means in the same column with different superscripts differ significantly (p<0.05) | |
| Table 4: | B(a)P induced DNA damage parameters determined by comet assay in lung of rats fed with Turmeric (T) |
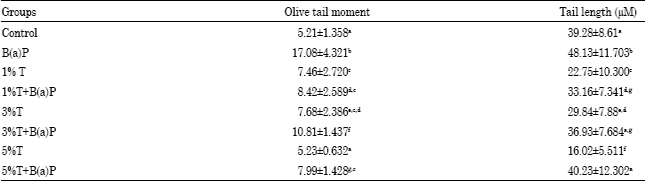 | |
| Values are mean±SD of 6 rats/group. Means in the same column with different superscripts differ significantly (p<0.05) | |
| Table 5: | Frequencies of mironucleated polychromatic erythrocytes induced by B(a)P in Bone marrow of rats fed with turmeric (T) |
 | |
| Values are mean±SD of 6 rats per group. Different superscripts are significant at p<0.05 between B(a)P and T+B(a)P groups by one-way ANOVA. | |
B(a)P treated group showed a significant increase in the formation of micronuclei (MN) which was approximately 3-fold higher compared to the normal control group. The frequencies of micronucleated polychromatic erythrocytes (MNPCE) in the control group and B(a)P group were 5.17±2.56 and 15.33±4.32 respectively. Turmeric feeding significantly inhibited the MN formation in 1, 3 and 5%, respectively. PCE/NCE ratio was also determined to see whether B(a)P had a cytotoxic effect on the bone marrow cells. The results showed a decrease in the PCE/NCE ratio in B(a)P group which is an indication of cytotoxicity. However the PCE/NCE ratios observed in B(a)P+turmeric groups at all the levels were found to be similar to the normal control compared to B(a)P group (Table 5). These observations reveal that turmeric has no genotoxic or cytotoxic effect but it has a protective effect in reducing the chromosomal damage and toxicity of B(a)P in the rodent bone marrow micronucleus assay.
DISCUSSION
There are large numbers of biomarkers available for assessing genotoxicity. Evidence from epidemiological, in vitro and in vivo studies indicate that a plant-based diet can reduce the risk of cancer and other chronic diseases (Rafter, 2002). The molecular mechanisms through which various nutrients might enhance or protect against carcinogenesis, the development of such biomarkers suitable for use in investigating the molecular effects of dietary factors in animal and human studies and also in vitro studies is of great importance.
Extensive research has been carried out in NIN on some important spices like turmeric, ginger, garlic, onion etc. for their genome protective effects. Turmeric, ginger and garlic have been demonstrated their antimutagenic effect using in vivo rat model (Polasa et al., 1991; Polasa et al., 2006; Nirmala et al., 2007a). The cytogenetic damage in terms of micronuclei was significantly reduced due to ginger and it could also inhibit DNA damage in peripheral blood lymphocytes (Nirmala et al., 2008, 2007b).
In this study turmeric was fed to rats in diet containing 0, 1, 3 and 5%. Significant decrease in the malondialdehyde (MDA) levels was observed in turmeric fed groups treated with B(a)P. This inhibition may be due to the active compound curcumin present in turmeric that may be protective against lipoperoxidation of subcellular membranes. In a study curcumin at 10 μM produced 40-85% inhibition of Low-density Lipoprotein (LDL) oxidation which was comparable to butylated hydroxyanisole (BHA) and it also could inhibit both initiation and propagation phases of LDL oxidation (Naidu and Thippeswamy, 2002).
In some in vitro studies turmeric prevented lipid peroxidation and protein glycosylation induced by higher concentrations of glucose (Jain et al., 2006). Protection of turmeric was observed against liver oxidative damage and induced genotoxicity by lead acetate in Swiss albino mice fed with turmeric at 5% level (El-Ashmawy et al., 2006). A new paradigm for diet and cancer prevention, research must be developed to include the nutrition modulation of the carcinogenesis pathways which include DNA damage and repair mechanisms, DNA methylation influencing gene expression, signaling pathways, cell cycle control and oxidative stress so that the studies will allow us to understand diet and cancer prevention research that includes identification of biomarkers for studies in various biological mechanisms underlying putative diet and cancer relationship (Shukla and Pal, 2004).
The Comet assay is a sensitive and rapid method to examine DNA damage and repair (Wasson et al., 2008). Dose related decrease in DNA migration in response to turmeric fed groups and B(a)P+turmeric fed groups was observed in liver, kidney and lung with respect to OTM and TL. The suppression of lipid peroxidation and oxidative DNA damage in rats by the intake of turmeric could be related to the prevention of carcinogenesis. Inhibitory effects of turmeric on the genotoxic effect of a powerful mutagen, urethane was observed in a dose dependent effect in a study (El-Hamss et al., 1999). Curcumin is a hydrophobic polyphenol. It is a diferuloylmethane and is able to scavenge or neutralize free radicals. In an intervention study curcumin was found to be very effective as a preventive approach in reducing the genotoxicity and oxidative stress induced by arsenic (Sinha et al., 2009). Curcumin which is one of the most exclusively studied compounds isolated from dietary source turmeric inhibited NF-KB and NF-KB regulated gene expression in various cancer cell lines and in vitro and in vivo studies showed that curcumin inhibited inflammation and carcinogenesis in animal models which include breast, oesophageal, stomach and colon cancer models (Basile et al., 2009). Curcumin was found to be non-toxic in six human trials with 25 subjects using up to 8000 mg of curcumin per day for 3 months (Chainani-Wu, 2003). Many studies have provided the evidence that cancer may be prevented by major life style changes (Anand et al., 2008b). Protective effect of curcumin on B(a)P induced DNA damage was reported in a study conducted in human peripheral blood lymphocytes in smokers, non-smokers and females (Polasa et al., 2004).
Urinary 8-OHdG has been widely used as a sensitive marker of the in vivo oxidative DNA damage and total system oxidative stress. It is formed in a promutagenic DNA lesion induced by the reaction of hydroxyl radicals with guanosine at the C8 site in DNA. Upon DNA repair, 8-OHdG is excreted in the urine. Numerous studies have indicated that 8-OHdG is not only a biomarker for cellular oxidative stress but also might be a risk factor for many diseases like cancer, diabetes etc. (Wu et al., 2004). To evaluate the potential oxidative stress markers the effect of turmeric at different levels was studied on oxidative damage. In all the experimental groups that received turmeric with B(a)P treated groups resulted in the decrease of 8-OHdG levels compared to normal control. One of the products of DNA oxidation, 8-OHdG has been extensively studied and its levels in target tissues are correlated with oxidative stress and the incidence of cancer.
The results obtained in this study shows the suppressive role of turmeric on the carcinogenic effect of B(a)P. In an animal study feeding of 1% curcumin in diet for 4 weeks abolished the formation of 4-hydroxynonenal-modified protein adducts, 8-OHdG and protein carbonyl formation in the kidneys of Ferric nitrilo triacetate and these protective effects may be mediated via its antioxidant properties (Iqbal et al., 2009). Administration of curcumin also inhibited diabetes-induced elevated retinal 8-OHdG levels suggesting that curcumin could inhibit the development of diabetic retinopathy, in part, via inhibiting accumulation of oxidized DNA in the retina (Kowluru and Kanwar, 2007).
Rodent bone marrow micronucleus assay is a widely used genotoxic assay to detect both clastogenic and aneugenic potencies of genotoxic agents or radiation (Ramalho et al., 1991). Numerous epidemiological studies have suggested that chromosomal alterations including formation of micronuclei may serve as an effective biomarker to estimate cancer risk. In this study it is evident from the results that feeding turmeric reduced the number of micronuclei in the groups that was given turmeric + B(a)P in comparison with the B(a)P group. An increased number of micronuclei in PCE in comparison with the control group indicate that B(a)P produces chromosomal damage in erythrocytes of bone marrow and this damage is associated with the appearance and/or progression of tumors with adverse reproductive and developmental outcomes (Krishna and Hayashi, 2000). The effect of curcumin when tested against gamma radiation induced in vivo chromosomal damage using the bone marrow micronucleus test significantly reduced the frequency of micronucleated PCE induced by the whole body exposure to gamma radiation (Abraham et al., 1993).
In a curry leaf supplemented diet in rats treated with dimethyl hydrazine chloride a 50% reduction was seen in the micronuclei and a 30% reduction in the activity of γ-glutamyl transpeptidase suggesting that curry leaves have a high potential as a reducer of the toxicity of DMP (Khanum et al., 2000). The overall mean percentage reduction of micronuclei was nearly 80% in the B(a)P+turmeric fed groups in our study. In another study Mentha piperita (peppermint) extract reduced the frequency of chromosomal aberrations and MN formation in bone marrow cells and decreased the levels of lipid peroxides and also enhanced the antioxidant enzymes and proved chemopreventive and antigenotoxicity when given an initiating dose of B(a)P in new born Swiss albino mice (Samarth et al., 2006).
The results of this study showed that turmeric has no genotoxic or cytotoxic effect but it has a protective effect in reducing the chromosomal damage induced by B(a)P in the rodent bone marrow micronucleus test. Turmeric exerted protective effect against B(a)P toxicity by modulating the lipid peroxidation, urinary 8-OHdG and DNA damage even at 5% level of feeding turmeric. The protective effect of turmeric against B(a)P induced DNA damage may be through free radical scavenging and modulation of DNA repair processes. Inhibition in the induction of micronuclei by turmeric feeding at different levels indicates its protective potential against genotoxicity.
The active compound present in turmeric is diet derived chemopreventive agent that provide mechanistic and pharmacological profile that might be useful to characterize the chemopreventive potential of the extract (Kelloff et al., 2000; Epstein et al., 2010). The discovery of disease prevention biomarkers associated with life style related diseases will give a way for understanding the risk of disease occurrence with the use of these biomarkers that may lead to the development of new health check that will contribute to preventive medicines (Anand et al., 2008a).
REFERENCES
- Anand, P., C. Sundaram, S. Jhurani, A.B. Kunnumakkara and B.B. Aggarwal, 2008. Curcumin and cancer: An old-age disease with an age-old solution. Cancer Lett., 267: 133-164.
Direct Link - Basile, V., E. Ferrari, S. Lazzari, S. Belluti, F. Pignedole and C. Imbriano, 2009. Curcumin derivatives: Molecular basis of their anticancer activity. Biochem. Pharmacol., 78: 1305-1315.
PubMed - Botsoglou, N.A., D.J. Fletouris, G.E. Papageorgiou, V.N. Vassilopoulos, A.J. Mantis and A.G. Trakatellis, 1994. Rapid, sensitive and specific thiobarbituric acid method for measuring lipid peroxidation in animal tissue, food and feedstuff samples. J. Agric. Food Chem., 42: 1931-1937.
CrossRefDirect Link - Celikler, S., S. Tas, O. Vatan, S. Ziyanok-Ayvalik, G. Yildiz and R. Bilaloglu, 2009. Anti-hyperglycemic and antigenotoxic potential of Ulva rigida ethanolic extract in the experimental diabetes mellitus. Food Chem. Toxicol., 47: 1837-1840.
PubMed - El-Ashmawy, I.M., K.M. Ashry, A.F. El-Nahas and O.M. Salama, 2006. Protection by turmeric and myrrh against liver oxidative damage and genotoxicity induced by lead acetate in mice. Basic Clin. Pharmacol. Toxicol., 98: 32-37.
Direct Link - El-Hamss, R., M. Analla, J. Compos-Sanchez, A. Alonso-Moraga, A. Munoz-Serrano and M. Idaomar, 1999. A dose-dependent antigenotoxic effect of turmeric. Mutat. Res., 446: 135-139.
PubMed - Epstein, J., I.R. Sanderson and T.T. Macdonald, 2010. Curcumin as a therapeutic agent: The evidence from in vitro, animal and human studies. Br. J. Nutr., 103: 1545-1557.
PubMed - Hu, M.L., C.H. Chuang, H.M. Sio and S.L. Yeh, 2002. Simple cryoprotection and cell dissociation techniques for application of the comet assay to fresh and frozen rat tissues. Free Radic. Res., 36: 203-209.
PubMed - Itokawa, H., Q. Shi, T. Akiyama, S.L. Morris-Natschke and K.H. Lee, 2008. Recent advances in the investigation of curcuminoids. Chin. Med., 3: 11-11.
CrossRef - Jagetia, G.C. and B.B. Aggarwal, 2007. Spicing up the immune system by curcumin. J. Clin. Immunol., 27: 19-35.
Direct Link - Jain, S.K., J. Rains and K. Jones, 2006. Effect of curcumin on protein glycosulation, lipid peroxidation and oxygen radical generation in human red blood cells exposed to high glucose levels. Free Radic. Biol. Med., 41: 92-96.
CrossRefPubMedDirect Link - Kelloff, G.J., J.A. Crowell, V.E. Steele, R.A. Lubet and W.A. Malone et al., 2000. Progress in cancer chemoprevention: Development of diet-derived chemopreventive agents. J. Nutr., 130: 467S-471S.
Direct Link - Khanum, F., K.R. Anilakumar, K.K.R. Sudarshana, K.R. Viswanathan and K. Santhanam, 2000. Anticarcinogenic effects of curry leaves in dimethylhydrazine-treated rats. Plant Foods Hum. Nutr., 55: 347-355.
CrossRefDirect Link - Kowluru, R.A. and M. Kanwar, 2007. Effect of curcumin on retinal oxidative stress and inflammation in diabetes. Nutr. Metab., 4: 8-8.
PubMed - Krishna, G. and M. Hayashi, 2000. In vivo rodent micronucleus assay: Protocol, conduct and data interpretation. Mutat. Res./Fundam. Mol. Mech. Mutagen., 455: 155-166.
CrossRefPubMedDirect Link - Moiseeva, E.P. and M.M. Manson, 2009. Dietary chemopreventive phytochemicals: Too little or too much. Cancer Prev. Res., 2: 611-616.
PubMed - Naidu, K.A. and N.B. Thippeswamy, 2002. Inhibition of human low density lipoprotein oxidation by active principles from spices. Mol. Cell Biochem., 229: 19-23.
Direct Link - Nirmala, K., T.P. Krishna and K. Polasa, 2007. In vivo antimutagenic potential of ginger on formation and excretion of urinary mutagens. Int. J. Cancer Res., 3: 134-142.
CrossRefDirect Link - Nirmala, K., T.P. Krishna and K. Polasa, 2008. Inhibition of induced micronuclei formation in human lymphocytes by ginger. Int. J. Cancer Res., 4: 12-19.
CrossRefDirect Link - Nirmala, K., T.P. Krishna and K. Polasa, 2007. Protective effect of ginger against benzo(a)pyrene induced DNA damage. Int. J. Cancer Res., 3: 13-24.
CrossRefDirect Link - Patil, B.S., G.K. Jayaprakasha, K.N.C. Murthy and A. Vikram, 2009. Bioactive compounds: Historical perspectives, opportunities and challenges. J. Agric. Food Chem., 57: 8142-8160.
CrossRefPubMedDirect Link - Peto, J., 2001. Cancer epidemiology in the last century and the next decade. Nature, 411: 390-395.
Direct Link - Polasa, K., A.N. Naidu, I. Ravindranath and K. Krishnaswamy, 2004. Inhibition of B(a)P induced strand breaks in presence of curcumin. Mutat. Res. Genetic Toxicol. Environ. Mutag., 557: 203-213.
CrossRef - Polasa, K., B. Sesikeran, T.P. Krishna and K. Krishnaswamy, 1991. Turmeric (Curcuma longa) induced reduction in urinary mutagens. Food Chem. Toxicol., 29: 699-706.
Direct Link - Anand, P., A.B. Kunnumakkara, C. Sundaram, K.B. Harikumar and S.T. Tharakan et al., 2008. Cancer is preventable disease that requires major life style changes. Pharm. Res., 25: 2097-2116.
PubMed - Rafter, J.J., 2002. Scientific basis of biomarkers and benefits of functional foods for reduction of disease risk: Cancer. Br. J. Nutr., 88: S219-S224.
PubMed - Ramalho, A.T., A.C.H. Nascimento, L.G. Littlefield, A.T. Natarajan and M.S. Sasaki, 1991. Frequency of chromosomal aberrations in a subject accidentally exposed to 137Cs in the Gorania(Brazil) radiation accident: Inter comparison among four laboratories. Mutat. Res. Environ. Mutagen. Related subjects, 252: 157-160.
CrossRef - Salamone, M.F. and K.H. Mavourin, 2005. Bone marrow Micronucleus assay: A review of the mouse stocks used and their published mean spontaneous micronucleus frequencies. Environ. Mol. Mutagen., 23: 239-273.
PubMed - Samarth, R.M., P. Meenakshi and A. Kumar, 2006. Modulatory effects of mentha piperita on lung tumor incidence, genotoxicity and oxidative stress in B(a)P treated Swiss albino mice. Environ. Mol. Mutagen., 47: 192-198.
PubMed - Singh, N.P., M.T. McCoy, R.R. Tice and E.L. Schneider, 1988. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res., 175: 184-191.
CrossRefPubMedDirect Link - Sinha, D., S. Mukherjee, S. Roy, R.K. Bhattacharya and M. Roy, 2009. Modulation of arsenic induced genotoxicity by curcumin in human lymphocytes. J. Environ. Chem. Ecotoxicol., 1: 1-11.
Direct Link - Wasson, G.R., V.J. McKelvey-Martin and C.S. Downes, 2008. The use of the comet assay in the study of human nutrition and cancer. Mutagenesis, 23: 153-162.
Direct Link - Wu, L.L., C.C. Chiou, P.Y. Chang and J.T. Wu, 2004. Urinary 8-OhdG: A marker of oxidative stress to DNA and a risk factor for cancer, atherosclerosis and diabetics. Clin. Chim. Acta, 339: 1-9.
CrossRefPubMedDirect Link - Chainani-Wu, N., 2003. Safety and anti-inflammatory activity of curcumin: A component of tumeric (Curcuma longa). J. Altern. Complementary Med., 9: 161-168.
CrossRefDirect Link - Banerjee, S., G. Bandyopadhyaya, K. Chattopadhyay and B.D. Chattopadhyay, 2010. Amelioration of nicotine-induced damage of blood cells in protein malnourished female rats by curcumin. Int. J. Pharmacol., 6: 444-455.
CrossRefDirect Link