
K. Paulkumar
Environmental Nanotechnology Division, Sri Paramakalyani Centre for Environmental Sciences, Manonmaniam Sundaranar University, Alwarkurichi-627412, TN, India
R. Arunachalam
Unit of Bio-monitoring Management, Department of Zoology, University of Madras, Guindy Campus, Chennai-600025, TN, India
R. Kameswaran
Department of Pharmaceutical Sciences, School of Pharmacy, University of Colorado Cancer Centre, University of Colorado Denver, Aurora, Colorado, USA
R. Ramanibai
Unit of Bio-monitoring Management, Department of Zoology, University of Madras, Guindy Campus, Chennai-600025, TN, India
G. Annadurai
Environmental Nanotechnology Division, Sri Paramakalyani Centre for Environmental Sciences, Manonmaniam Sundaranar University, Alwarkurichi-627412, TN, India
International Journal of Cancer Research
Year: 2010 | Volume: 6 | Issue: 1 | Page No.: 27-34







ABSTRACT
The present investigation made in documenting the in vitro anti-proliferative activity of indirubin-3’-monoxime on human epithelial larynx cell line (Hep-2 cells). The Indirubin propensity for anti-proliferative effects was studied and its mechanism of action was documented using the techniques Trypan blue staining, MTT assay and propidium iodide staining. We observed a dose dependent cell death on treatment with indirubin-3-monoxime. On the basis of results, we concluded, indirubin-3’-monoxime inhibits the proliferation of Human laryngeal cancer cells (Hep-2).
PDF Abstract XML References Citation
How to cite this article
K. Paulkumar, R. Arunachalam, R. Kameswaran, R. Ramanibai and G. Annadurai, 2010. Anti-Cancer Effect of Indirubin-3'-Monoxime for Human Laryngeal Carcinoma. International Journal of Cancer Research, 6: 27-34.
DOI: 10.3923/ijcr.2010.27.34
URL: https://scialert.net/abstract/?doi=ijcr.2010.27.34
DOI: 10.3923/ijcr.2010.27.34
URL: https://scialert.net/abstract/?doi=ijcr.2010.27.34
INTRODUCTION
Cancer research is increasingly being drawn towards the investigation of plant derived anti-cancer compounds many of which have been used traditionally as herbal remedies for centuries (Sunilson et al., 2009). In the traditional Chinese medicine, the preparation Danggui Longhui Wan, consisting of 11 herbal medicines, has been used for the treatment of various chronic diseases (Marko et al., 2001) such as leukemia (Sridhar et al., 2006). One of the major herbal products Indirubin had the capacity to stop the uncontrolled growth of tumor cells by inactivating enzymes called Cyclin-dependent kinases (CDKs). The CDKs play a central role in the initiation, ordering and completion of cell cycle events. Mutations that deregulate these kinases, result in cancer (Fabbro et al., 2002) tumor formation, neurodegenerative disorders (e.g., Alzheimer’s disease, stroke) (Geschwind, 2003), cardiovascular disorder. Some of the CDKs isozymes are specific to certain kinds of tissues. Aberrant expression and overexpression of these kinases are evidenced in many disease conditions. Inhibition of isozymes of CDKs specifically can yield beneficiary treatment modalities with minimum side effects (Sridhar et al., 2006).
One problematic issue with kinase inhibitors was specificity since they generally bind at the relatively conserved ATP binding pocket of the kinase family (Bain et al., 2003; Knight and Shokat, 2005). However, it has also recently been suggested that the biological activity of kinase inhibitors may rely on a multispecific inhibition of different kinases (Cohen, 2002; Dancey and Sausville, 2003). In order to be able to judge both positive and negative effects of multi-specificity, it is necessary to reliably identify the cellular targets of inhibitors. Conventionally, kinase inhibitor activity and specificity was tested in vitro within a defined panel of isolated kinases (Bain et al., 2003).
Marko et al. (2001) reported from Cooperative Group of Clinical Therapy of Indirubin-1980 reports that the cell cycle of cancer cells have been treated with indirubin or its derivative was arrested in the cell cycle thereby resulting in inhibition of cell proliferation and ultimately induction of apoptosis. Indirubin and its derivatives such as indirubin-3-monoxime had low toxicity, limited side effects and also it has been reported to inhibit DNA and protein synthesis in various cell lines. However, the underlying mechanism of action of indirubin has remained largely unknown.. In this study, in the in vitro condition, indirubin-3’-monoxime (powder) was examined on Human laryngeal cancer cells (Hep-2) and the indirubin propensity for anti-proliferative effects was studied and its mechanism of action was documented using the morphological assay, Tryphan blue staining, MTT assay and propidium iodide staining techniques.
MATERIALS AND METHODS
Human laryngeal cancer cell lines were purchased from King Institute of Technology, Chennai. Indirubin-3’-monoxime, Growth medium Fetal Bovine Serum (FBS), RPMI-1640 medium and Penicillin streptomycin were procured from Gibco laboratory. Indirubin-3’-monoxime was dissolved in Dimethyl sulfoxide (DMSO) which was purchased from Sigma, USA. All other chemicals used in this study were of analytical grade. The tissue culture flask (TC-flask) and plates were purchased from Tarson, India.
Morphology and Maintenance of the Hep-2 Cells
The investigation was performed during the period of November, 2006 to February, 2007. The morphology of the cells was determined by using light microscope. The cells were cultured from the mother culture. For maintaining the cell lines Hep-2 cells were isolated from Human epidermoid of larynx. The cells were grown in TC bottles containing growth medium MEM without FCS and it was incubated at 37°C in CO2 incubator. After incubation period, tissue culture flasks (TC flask) were showed a confluent monolayer was observing cell lines under an inverted microscope. The formation of monolayer proved the growth of cells in the TC flasks. Growth medium was removed from the flasks and the flask were washed with PBS/MEM without FCS and then 5 mL of trypsin phosphate buffered saline versene glucose (TPVG) was added on the monolayer and withstand for 2-3 min. After, TPVG was removed and the flasks were incubated at 37°C until all the cells were detached from the TC flask surface. Then the cells were re-suspended in 5 mL of growth media containing MEM without FCS (5% growth medium was used). The cell suspension was aspirated for few times to break the cell clumps. Cell lines were not passages >14 times due to maintenance of cell stability. The cell concentration (growth concentration) was determined by counting the cells in haemocytometer (Jun et al., 2007).
Trypan Blue Assay
Trypan blue assay was used for counting the cells (Allison and Ridolpho, 1980). The cell suspension 0.2 mL was assorted with 0.2 mL of trypan blue (0.1% trypan blue). It was mixed well with pipette and sufficient volume was aspirated to fill haemocytometer immediately. The viable cells (non viable cells are stained blue and viable cells remained unstained) were counted in each of the four corners of both chambers, omitting cells lying on the top and to the left. The cell clumping was observed and it was discarded and original cell suspension was resuspended. The total number of cells present in the cell suspension was calculated using the following formula:
Average cell countxdilution factorx104 = Lakhs mL-1 (or) C1 = txtbx¼x104
From this formula, C1 was a initial cell concentration per mL. t was a total viable cell count of 4 squares. tb was a correction to the trypan blue dilution. ¼ was a correction to gives mean cells/corner square. 104 was a conversion factor for counting chamber. Based on the cell count, sterile flasks were seeded and incubated at 36°C. For flasks 1.0x105 cells were seeds approximately. All the passages were recorded.
MTT Assay
The Microculture Tetrazolium viability (MTT) assay was used for finding the viability of cells. The cytotoxicity of the drug was determined by MTT assay (Mossman, 1983). A solution of 5 mg mL-1 of MTT was dissolved in Phosphate Buffered Saline (PBS) and it was filtered and sterilized. At the end of incubation of the 24 well plates, media was removed and 100 μL of MTT was added to each well containing the cells. This was incubated at 37°C in a CO2 incubator for 5 h. After incubation the MTT solution was removed and 1 mL of DMSO was added to each well and mixed the solution thoroughly to dissolve the crystals and incubated it for few min. The cell viability (or) cytotoxicity of the drug was determined by spectrophotometer. Based on the absorbance values the graph was plotted and viable cells were evaluated.
Propidium Iodide Staining of Apoptotic Cells
The programmed death of cells (apoptosis) was determined by the propidium iodide staining technique. The cells (Hep-2) were seeded on cover slips and placed in petriplates at a concentration of approximately 10 lakhs cells per plate. The plates were incubated in CO2 incubator for 48 h to obtain a sheet of cells on the coverslips. After incubation the media was removed and the coverslips were gently washed with phosphate buffered saline. Before the addition of drug, it was mixed with PBS and then slowly added to the plates until the coverslips were fully immersed. The plates were incubated for 24 h in CO2 incubator. After incubation the cover slips were fixed with ethanol fixative and then stained with 100 μL of propidium iodide and stand it for few min. After that the apoptotic cells were identified by using inverted fluorescent microscope (Idziorek et al., 1995).
RESULTS
Morphology
In morphological assay the matured cells adhered to the bottom of the TC flask. The young and dead cells are floating in the medium (Fig. 1a, b). Figure 1a and b showed young, matured and dead cells. The matured cells adhered at the bottom of the TC flask. The Morphology detection on 2nd Day.
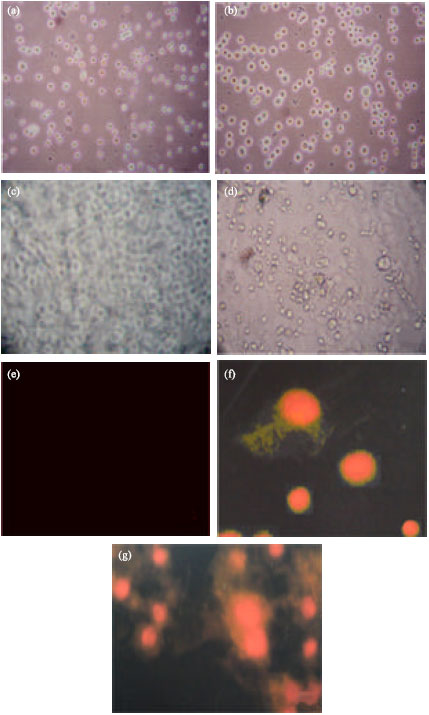 | |
| Fig. 1: | Cancer cell line (Hep-2) with indirubin at different assays. (a) (b) Morphology detection on 5th Day. (c) Tryphan blue assay-control. (d) Tryphan blue assay-cancer cell (Indirubin at 50 μg concentration). (e) Propidium iodide staining test for Control cells. (f) Propidium iodide staining for Hep-2 cells at initial stage. (g) Propidium iodide staining for Hep-2 cells at lateral (Indirubin at 50 μg concentration) |
young cells are floating in the medium and the dead cells also floating in the medium. After the maturation of young cells it adheres to the bottom of the TC flask. The cells were taken 5-7 days for maturation. The young cells were allowed the light through it, but the dead cells were not allowed the light, because of degranulation of nucleus.
Trypan Blue Assay
Continuation of the morphological assay, the trypan blue assay was used for calculating the viable cells. In this assay, we were examined that the viable cells were not absorbed blue dye, but the dead cells were absorbed blue dye due to the degraded cell membrane by the action of drug indirubin-3’-monoxime. Figure 1c and d show the viable cells without blue color and blue color emitting dead cells. The viable cells counted through haemocytometer. The 50 μg concentration of indirubin-3’-monoxime was used for trypan blue assay.
Propidium Iodide Staining
Apoptosis of the drug was determined by propidium iodide staining. The dead cells were emitted the red color, but the live cells were not emitted the red color, because of the indirubin was degraded the cell membrane and the stain was entered and emitted the red color. It was identified through fluorescent microscope. Figure 1e and f were showed the indirubin apoptosis effect on Hep-2 cell line. The apoptosis effect of the drug was performed and the red color emitted dead cells proved the apoptotic effect of the indirubin against cancer cell line Hep-2. The drug was showed the effect on 50 μg concentrations in propidium iodide staining.
MTT Assay
Cytotoxicity assay of the drug indirubin was determined by the MTT assay. The MTT salt was mixed with different concentration of the drug treated cell suspension. After the 24 h CO2 incubation at 37°C the reading was taken in the spectrophotometer. The OD values were obtained and it was showed in Fig. 2. In MTT assay, the increased OD value was showed that the effect of the drug at 10, 20, 30, 40 and 50 μg concentration. The Fig. 2 suggested that the concentration of the drug was increased the cytotoxicity effect of the drug may also increased. The indirubin-3’-monoxime was showed the high cytotoxic effect at 50 μg concentrations, the counted live cells were 285,000 lakhs mL-1 and the OD value was 3.1 (Fig. 2). The control (without drug) was showed, the counted live cells were 100,000 lakhs mL-1 and the OD value was 1.2.
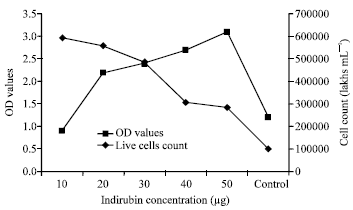 | |
| Fig. 2: | MTT assay X-axis shows Indirubin concentration (μg), Y-axis shows spectrophotometer OD values of indirubin at different concentration and second Y-axis shows the cell count (lakhs mL-1) |
DISCUSSION
Sridhar et al. (2006) were reported that several derivatives of (oxindoles) indirubin have been synthesized to improve their bioavailability and potency. Analogues such as indirubin sulfate and indirubin monoxime have shown good activity and moderate selectivity for CDKs. Bramson et al. (2001) described, some hydrazone analogs had high potency of inhibition for CDK2 and CDK1. Merz et al. (2007) accounted, the indirubin derivative; indirubin-5-carboxamides effectively inhibit the growth of a wide spectrum of human tumor cell lines with low micromolar to nanomolar IC50 values. This compound induce cell cycle arrest at G1/S and/or G2/M phase in a wide spectrum of human lung tumor cell lines LXFL529L, resulting in induction of apoptosis. Hoessel et al. (1999) results were suggested, indirubin-3’-monoxime was potent inhibitors of cyclin-dependent kinase and it had anti-proliferative activity, loading to a G2/M arrest in almost all cell types (Fig. 3) studied and to G1/S arrest in Jurkat cells, the most sensitive cell line. This lather arrest is associated with an inhibition of phosphorylation of the retinoblastoma protein. Briefly their observations are compatible with an inhibition of CDKs. Marko et al. (2001) and Lee et al. (2005) were reported, indirubin-3’-monoxime exerts potent tumors cell growth inhibition and that this relates to inhibition of CDKs. As a result, progress through the cell cycle is blocked at G1/S and G2/M events that might result in the induction of apoptosis. Goergens and Esser (2007) were found apoptosis in human acute myeloid leukemia cell line (HL-60) for association with anti-leukemic effects. In this study, indirubin-3’-monoxime; a derivative of indirubin was arrested the proliferation and induced the apoptosis of the Hep-2 cancer cells in G1/S or G2/M phase of cell cycle (Fig. 3). This experimental results is similar to Lee et al. (2005) and Hoessel et al. (1999) results mentioned above. These above the results were suggested that indirubin and its derivatives had acted against different cancer cells as an inhibitor and it was arrest the proliferation and induced the cell apoptosis.
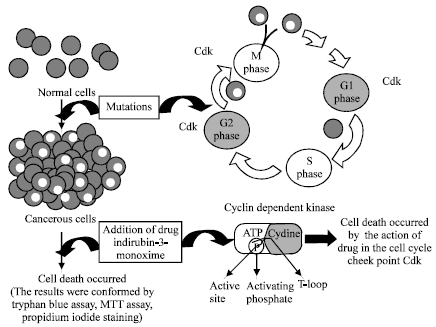 | |
| Fig. 3: | Normal cells are became a cancerous due to the role of CDKs mutation which might occurred in G1/S phase and or G2/M phase of cell cycle.Proliferation or degraded the cancer cells were done by indirubin-3’monoxime. Indirubin-3’-monoxime exerts potent tumors cell growth inhibition and that this relates to inhibition of CDKs. As a result, progress through the cell cycle is blocked at G1/S and or G2/M events that might result in the induction of apoptosis in almost all cell types (Hep-2) |
Goergens and Esser (2007) were reported the anti-proliferative activity of Indirubin-3’-monoxime against HL-60 cells by the induction of caspase-3 mediated apoptosis. Zahler et al. (2007) were accounted the proliferation assay for proliferation of MCF-7 cancer cells by using crystal violet test as described in NCI protocols. They were used the three derivatives of indirubin, namely 5-bromo-indirubin-3’oxime, 6-bromo-indirubin-3’oxime and 7-bromo-indirubin-3’oxime for examining their influence on proliferation of MCF-7 cancer cells. In MCF-7 cells, 6-bromo-indirubin-3’oxime completely blocked the proliferation at 10 μM, while the other compounds afforded only 50% inhibition at this concentration. Meijer et al. (2000) reported that Indirubin-3’-monoxime had limited effect IC50 values of 600 and 700 nM on CDK4/cyclin D1 and CDK6/cyclin D2. In this study, the high anti-proliferation activity was found at 50 μg concentration of Indirubin-3’-monoxime in Hep-2 cells (Fig. 2). These were suggested, the high concentration of drug might had high anti-proliferate and cytotoxic (counted live cells were 285,000 lakhs mL-1 and the OD value was 3.1) activity in Hep-2 cells and also indirubin derivatives had different sub group chains, on the basis of this sub group arrangement it could be activated against the different receptors including CDK/CDKs of cancer cell lines. Our observation reveals that high concentration of indirubin-3’-monoxime might had high anti-proliferate and cytotoxic effects than less concentration.
CONCLUSION
Results presented here demonstrated that several independent assays could be used in complementary fashion in Hep-2 cells to gain a better understanding of various underlying aspects of anti-proliferative activity of drug in different concentration were carried out successfully. Collectively, our easiest approach to inhibit and degrade the cancer cells was showed the drug effect against proliferation of cancer cells was ~>50% (the results dependent on the role of side chains group of the drug in CDKs). It was showed the indirubin-3’-monoxime had potent anti-proliferative effect on cancer cell line (Hep-2) (Fig. 3). In future, new and similar derivatives of indirubin should be applied to investigate the complex mechanism of anti-proliferative, anti-cancer activity and allowing the candidate drug for careful assessment of discovery and in vivo studies.
ACKNOWLEDGMENT
Authors thanks Manonmaniam Sundaranar University, Alwarkurichi, India and University of Madras, Chennai, India for provided necessary facilities.
REFERENCES
- Allison, D.C. and P. Ridolpho, 1980. Use of a trypan blue assay to measure the deoxyribonucleic acid content and radioactive labeling of viable cells. J. Histochem. Cytochem., 28: 700-703.
Direct Link - Bain, J., H. McLauchlan, M. Elliott and P. Cohen, 2003. The specificities of protein kinase inhibitors: An update. Biochem. J., 371: 199-204.
PubMed - Bramson, H.N., J. Corona, S.T. Davis, S.H. Dickerson and M. Edelstein et al., 2001. Oxindole-based inhibitors of cycline-dependent kinase 2 (CDK2) design, synthesis enzymatic activities and X-ray crystallographic analysis. J. Med. Chem., 44: 4339-4358.
PubMed - Cohen, P., 2002. Protein kinases: The major drug targets of the twenty-first century?. Nat. Rev. Drug Discov., 1: 309-315.
PubMed - Dancey, J. and E.A. Sausville, 2003. Issues and progress with protein kinase inhibitors for cancer treatment. Nat. Rev. Drug Discov., 2: 296-313.
PubMed - Fabbro, D., S. Ruetz, E. Buchdunger, S.W. Cowan-Jacob and G. Fendrich et al., 2002. Protein kinases as targets for anticancer agents: From inhibitors to useful drugs. Pharmacol. Ther., 93: 79-98.
PubMed - Geschwind, D.H., 2003. Tau phosphorylation, tangles and neurodegeneration: The chicken or the egg. Neuron, 40: 457-460.
PubMed - Hoessel, R., S. Leclerc, J.A. Endicott, M.E.M. Noble and A. Lawrie et al., 1999. Indirubin, the active constituent of a Chinese antileukemia medicine, inhibits cycline-dependent kinases. Nat. Cell Biol., 1: 60-67.
Direct Link - Idziorek, T., J. Estaquier, F. De-Bels and J.C. Amerisen, 1995. JC YOPRO-1 permist cytofluorometric analysis of programmed cell death (apoptosis) without interfering with cell viability. J. Immuno. Meth., 185: 249-258.
PubMed - Knight, Z.A. and K.M. Shokat, 2005. Features of selective kinase inhibitors. Chem. Biol., 12: 621-637.
PubMed - Lee, J.W., M.J. Moon, H.Y. Min, H.W. Chung and E.J. Park et al., 2005. nduction of apoptosis by a novel indirubin-5-nitro-3-monoxime, a CDK inhibitor, in human lung cancer cells. Bioorg. Med. Chem. Lett., 15: 3948-3952.
PubMed - Marko, D., S. Schatzle, A. Friedel, A. Genzlinger, H. Zankl, L. Meijer and G. Eisenbrand, 2001. Inhibition of cyclin-dependent kinase 1 (CDK1) by indirubin derivatives in human tumor cells. Br. J. Cancer, 84: 283-289.
PubMed - Meijer, L., A.M.W.H. Thunnissen, A.W. White, M. Garnier and M. Nikolic et al., 2000. Inhibition of cyclin-dependent kinases, GSK-3β and CK1 by hymenialdisine, a marine sponge constituent. Chem. Biol., 7: 51-63.
PubMed - Mosmann, T., 1983. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods, 65: 55-63.
CrossRefPubMedDirect Link - Sridhar, J., N. Akula and N. Pattabiraman, 2006. Selectivity and potency of cycline-dependent kinase inhibitors. AAPS J., 8: E204-E221.
PubMed - Jun, X.X., H.Q. Guo, P.Z. Cai., D.R. Dan and H.Z. Sheng, 2007. Morphological study on apoptosis Hela cells induced by soyasaponins. Toxicol. In vitro, 21: 820-826.
PubMed - Zahler, S., S. Tietze, F. Totzke, M. Kubbutat, L. Meijer, A.M. Vollmar and J. Apostolakis, 2007. Inverse in silico screening for identification of kinase inhibitor targets. Chem. Biol., 14: 1207-1214.
CrossRef - Goergens, A. and C. Esser, 2007. 48th spring meeting deutsche gesellschaft fur experimentelle und klinische pharmakologie und toxikologie mainz, March 13-15, 2007. Naunyn-Schmiedeberg's Archives Pharmacol., 375: 1-111.
CrossRefDirect Link - Sunilson, J.A.J., G. Rejitha, K. Anandarajagopal, A. Das, M. Muthappan and P. Promwichit, 2009. Cytotoxic effect of Cayratia carnosa leaves on human breast cancer cell lines. Int. J. Cancer Res., 5: 115-122.
CrossRefDirect Link