
Sanem Yildiz Arikan
Department of Biophysics, Faculty of Medical, Pamukkale University, Kinikli, Denizli, Turkey
Ayfer Atalay
Department of Biophysics, Faculty of Medical, Pamukkale University, Kinikli, Denizli, Turkey
Erol Omer Atalay
Department of Biophysics, Faculty of Medical, Pamukkale University, Kinikli, Denizli, Turkey
International Journal of Cancer Research
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 251-256







ABSTRACT
The ability of a peptide to target a specific protein in vitro has a potential for recognizing the cell membrane structure, protein-protein and receptor-ligand interaction. On the basis of molecular interactions, cell specific peptides were selected via phage library approach and their functions on the cells were determined by XTT based viability assay. We aimed to find phage displayed peptides from 12-mer peptide library that interact with K562-dox cell membrane in association with cellular functions and to study the effects of peptides selected from artificial peptide library on K562-dox cell viability. Peptides recognizing K562-dox cells were identified according to peptide sequences and amino acid properties. We selected 29 different phages from biopannings with K562-dox cells. Three clones were identified (KPB7, KPB10, KPP8) and their negative effects on cell viability were determined by XTT assay. According to our cell viability assay results, selected phages were effected negatively to the viability of the K562-dox cells. Depending on the present results, peptides were obtained that could be potential candidates for molecular recognition and cell targeting approaches.
PDF Abstract XML References Citation
Received: July 20, 2010;
Accepted: October 21, 2010;
Published: November 04, 2010
How to cite this article
Sanem Yildiz Arikan, Ayfer Atalay and Erol Omer Atalay, 2010. Interaction of Peptides Selected from Artificial Peptide Library with Doxorubicin Resistant K562 Cells. International Journal of Cancer Research, 6: 251-256.
DOI: 10.3923/ijcr.2010.251.256
URL: https://scialert.net/abstract/?doi=ijcr.2010.251.256
DOI: 10.3923/ijcr.2010.251.256
URL: https://scialert.net/abstract/?doi=ijcr.2010.251.256
INTRODUCTION
Cell lines are being used to investigate the structural and functional properties of living organisms. Cell culture is one of the laboratory methods to observe cancer cells activity such as proliferation and differentiation. Phage display is also another laboratory method based on peptide libraries to obtain peptides with high affinity and specificity with the target molecule under interest. K562 cell lines were described as human erythroleukemia cells derived from chronic myeloid leukemia patient (Lozzio and Lozzio, 1975). K562-dox cells are well characterized K562 cell line, which are resistant to doxorubicin. These doxorubicin resistant cells were cultured in vivo or in vitro by various groups (Hamada and Tsuruo, 1988; Hait et al., 1993; Arora et al., 2004). K562 cell lines can proliferate to erythroid series by hemin, hydroxyurea or various antracyclines and well detected by cell surface markers. The difference of doxorubicin resistant cell lines from wild type K562 is the over expression of drug resistance proteins, especially p-glycoprotein. Therefore, these cells constitute models for cell viability and proliferation assays and for understanding drug resistance mechanisms of cancer cells (Koeffler and Golde, 1980; Tsiftsoglou et al., 2003; Dos-Santos et al., 2009).
Artificial peptide libraries are composed of recombinant phages each displaying different peptide or proteins. Phage display is a powerful tool for the selection and identification of peptides allowing the presentation of large peptide libraries on the coat protein of filamentous phage. Biopanning with artificial phage library presents us how to select the best specific peptide to the target under interest (Smith and Petrenko, 1997; Hoogenboom and Chames, 2000). This approach has been used in a number of applications as protein-protein interactions, epitope-mimotope mappings, studies of cancer cells, drug discovery etc (Benhar, 2001; Sergeeva, 2006; Chua et al., 2008).
In this study we aimed to find phage displayed peptides that interact with cell membrane in association with cellular functions. We aimed to study the effects of peptides selected from artificial peptide library on K562-dox cell viability.
MATERIALS AND METHODS
Cells and Cell Culture
Wild type K562 and doxorubicin resistant K562 (K562-dox) cell lines are obtained from Dr. Akça (Pamukkale University, School of Medicine, Denizli, Turkey) in 2008. Cells were incubated in a humidified environment with 5% CO2, 95% air at 37°C in RPMI 1640 medium supplemented with 10% Fetal Bovine Serum (FBS), 100 units mL-1 penicillin-streptomycin. K562-dox cells were cultured in 3 μM doxorubicin.
Selection Procedure (Biopanning)
PhD-12TM Phage Display Peptide Library Kit was purchased from New England Biolabs Inc. (NEB). Biopanning procedures were done according to the manufacturer’s instructions with certain modifications. PhD-12TM phage display peptide library was used with K562 and K562-dox cells for five biopannings. The titer of the library is 1,5x1013 pfu mL-1 and the complexity is 2.7x109 transformants. We used 1x106 cells and 2x1011 phages for each biopanning. First biopanning was done with K562-dox cells, the second and third were with K562 cells and last two pannings were done with K562-dox cells. Bound phages to K562-dox cells were eluted with 0.2 M glycine-HCl (pH: 2.2) and neutralized with 1 M Tris-HCl. On the other hand, the integrated phages were separated from cells at room temperature by vortexing vigorously. After each panning, phage solution was titered to measure the complexity of bound and integrated phages. After the fifth round, from the titered phage stock we collected 50 phage plaques randomly. Each phage was amplified with ER2738 bacteria to 2x1013 phages mL-1.
Phage DNA Isolation and Nucleotide Sequencing
Single-stranded phage DNA was isolated from each amplified phage clone according to the manufacturer’s instructions. Sequence analysis of the selected phage clones was done by Beckman Coulter CEQ 8000 automated sequencing system. In total 29 different DNA sequences out of 50 clones were identified.
XTT Assay
K562-dox cells were seeded onto 96-well plate at a concentration of 1x105 cells/well and treated with or without phage. Phage concentrations were 1x1011 phages per well. After incubation at 37°C with 5% CO2 for 24 h, XTT (Biological Industries, Israel Beit Haemek Ltd.) was added to each well at a volume of 50 μL. The cells were further incubated for 24 , 48 and 72 h to allow XTT production. Absorbance (A), which was proportional to cell viability, was then measured at a wavelength of 450 nm. Cell viability was measured using the following equation:
where, A1 and A0 are the absorbance values obtained from treated and untreated cells, respectively.
The assay was repeated four times for each phage and mean value was taken for the percentage of cell viability calculations. By using phage display and XTT assay, we report the peptides that specifically recognize the membrane of K562-dox cells and their effects to cell viability by measuring the survival rate of the K562-dox cells.
Statistical Analysis
We used XTT proliferation assay data for Chi square (χ2) analysis to determine the survival rate of cells.
RESULTS AND DISCUSSION
In this study, we used PhD-12 phage display peptide library for targeting K562-dox cells to select the peptides which recognized doxorubicin resistant K562 cells specifically instead of wild type K562 cells. The phages were named as bound (KPB) and integrated (KPP) phages and amplified 25 KPB and 25 KPP phages after the fifth biopanning. By using Beckman CEQ8000 DNA sequencing analysis system we observed 29 different clones out of 50 selected phages. According to the results of cell viability assay, we determined different effects of phages on K562-dox cells viability. We compared the effects of phages on cell viability as phage in the presence and absence of doxorubicin. All control doxorubicin resistant K562 cells were treated only with doxorubicin. XTT assay results of control cells were measured as shown in Fig. 1a.
The amino acid sequence of phage KPB7, which binds to K562-dox cells, was determined as SLNTTFTGPSRP and XTT assay results were shown in graphics (Fig. 1b). The % viability of cells were measured 24, 48 and 72 h as 51% (χ2: 4.9; p<0.05), 30% (χ2: 4.2; p<0.05) and 24% (χ2: 14.1; p<0.05), respectively. According to the peptide sequence of phage KPB7, the hydrophobicity was calculated as 66% and the peptide includes 50% uncharged polar R groups.
The phage KPB10 amino acid sequence was determined as TSHHDSHGLHRV by DNA sequence analysis. XTT assay results were calculated for 24, 48 and 72 h as 35% (χ2:6.7; p<0.05), 48% (χ2:4.8; p<0.05) and 22% (χ2:21,7; p<0.05), respectively (Fig. 1c). The hydrophobicity was determined as 50% and the peptide includes 42% positively charged groups.
The phage KPP8 amino acid sequence was determined as FMPKSHKSRLPS by DNA sequence analysis. XTT assay results were calculated for 24, 48 and 72 h as 58% (χ2:14.3; p<0.05), 48% (χ2:9.3; p<0.05) and 27% (χ2:41.5; p<0.05), respectively (Fig. 1d). The hydrophobicity was determined as 50% and the peptide includes 42% non-polar groups. Phage KPP8 is more negatively effects on cell viability (39%) than the other phages and phage KPP8 without doxorubicin.
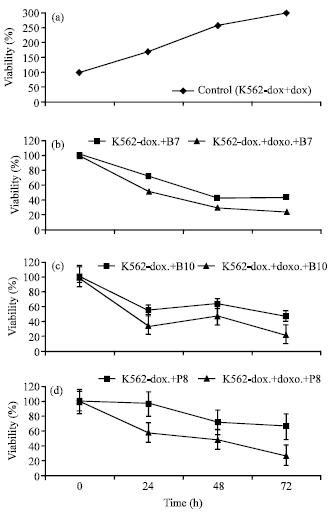 | |
| Fig. 1: | (a) XTT assay results of control cells, (b) XTT assay results of Phage KPB7, (c) XTT assay results of Phage KPB10 and (d) XTT assay results of Phage KPP8 |
According to the results of XTT assay, three phage clones (KPB7, KPB10 and KPP8) effected cell viability negatively by doxorubicin. We investigated the physicochemical properties of these peptides and not found any similarities between peptide sequences.
Phage display describes a selection technique in which a peptide or protein is expressed as a fusion with a coat protein of a bacteriophage, resulting in display of the fused protein on the surface of the bacteriophage. Phage display has been used to create a physical linkage between a vast library of random peptide sequences to the DNA encoding each sequence, allowing rapid identification of peptide ligands for a variety of target molecules(antibodies, enzymes, cell-surface receptors, etc.) by an in vitro selection process called biopanning as eluting the specifically-bound phage to the target molecule. The eluted phage is then amplified and taken through additional binding/amplification cycles to enrich the pool in favor of binding sequences. After several rounds, individual clones are characterized by DNA sequencing (Hoogenboom and Chames, 2000; Azzazy and Highsmith, 2002).
In this study, according to the viability assay and statistical results, the phages KPB7, KPB10 and KPP8 were found negatively effected phages on cell viability together with doxorubicin by recognizing the different structures of the cell membrane. So that this makes us to think this effect should be about doxorubicin resistance mechanism and recognizing the cell membrane should be on different membrane structures. These findings show that peptides which are expressed on phages could recognize the K562-dox cells and effects on different mechanisms of cell viability.
Phage KPB10 (TSHHDSHGLHRV) was found negatively effected the viability of K562-dox cells with doxorubicin. This peptide sequence was found after the biopannings with Salmonella enterica serovar Typhi Vi antigen. This peptide mimics the immune dominant mimotopes of capsular polysaccharide (ViCPS) antigen (Tang et al., 2003). This results gives an opinion about the peptide could bind the carbohydrate structures on K562-dox cell membrane.
Except for these three clones we investigated another five clones (KPB1, KPB17, KPB25, KPP10 and KPP20). Phage KPB1 (LLADTTHHRPWT) was also negatively affected on cell viability but less than the other phages (data not shown). According to Su et al. (2006) this phage peptide sequence was specific for single-walled carbon nanotubes that used in biotechnological approaches as coating tubes with different molecules. Na et al. (2003) showed that this peptide could also recognize TSH receptors for defining the auto antibodies in Grave’s disease.
Phage KPB17 (EHMALTYPFRPP) has a potential to recognize the non-small cell lung cancer cells (NSCLC) and can be useful to produce drug carriers according to Zang et al. (2009). In present study, KPB17 can recognize K562-dox cells and effected without doxorubicin negatively to cell viability. In this respect, common membrane structures should be investigated between K562 and NSCL cells.
Lee et al. (2007) showed the same peptide sequence as phage KPB25 (WPTYLNPSSLKA) in an in vivo study of phage display to find tumor homing peptides. This suggests that phage display technique has advantageous to find cell specific (in vitro) or organ specific (in vivo) peptides and to develop new drug carriers (Lee et al., 2007).
Another phage KPP20 was also specific for K562-dox cells but not found any effect on cell viability. According to Dickerson et al. (2004) this phage has a special feature for perceiving germania nanoparticle.
In conclusion using artificial peptide libraries plays an important role on targeting cancer cells and getting peptides that effect cell functions by recognizing different cell structures. These findings make us to think the peptides specifically recognize different structures as cell, nanoparticles or proteins. This point out those similar structures between different targets can recognize same peptides and these peptides can be used as potential candidates for biophysical and molecular studies. These results suggest that the mechanisms of doxorubicin resistance, cell proliferation and apoptosis should be studied by new generation technologies as AFM, SPR, flow cytometry, patch clamp etc.
ACKNOWLEDGMENTS
This study was supported by Pamukkale University Research Fund, Project Number: 2008SBE005.
REFERENCES
- Lozzio, C.B. and B.B. Lozzio, 1975. Human chronic myelogenous leukemia cell-line with positive Philadelphia chromosome. Blood, 45: 321-334.
PubMed - Hait, W.N., S. Choudhury, S. Srimatkandada and J.R. Murren, 1993. Sensitivity of K562 human chronic myelogenous leukemia blast cells transfected with a human multidrug resistance cDNA to cytotoxic drugs and differentiating agents. J. Clin. Invest., 91: 2207-2215.
Direct Link - Koeffler, H.P. and D.W. Golde, 1980. Human myeloid leukemia cell line: A review. Blood, 56: 344-350.
PubMed - Tsiftsoglou, A.S., I.S. Pappas and I.S. Vizirianakis, 2003. Mechanisms involved in the induced differentiation of leukemia cells. Pharmacol. Therapeutics, 100: 257-290.
CrossRef - Dos-Santos, G.G., J. Reinders, K. Ouwehand, T. Rustemeyer, R.J. Scheper, S. Gibbs, 2009. Progress on the development of human in vitro dendritic cell based assays for assessment of the sensitizing potential of a compound. Toxicol. Applied Pharmacol., 236: 372-382.
PubMed - Hoogenboom, H.R. and P. Chames, 2000. Natural and designer binding sites made by phage display technology. Immunol. Today, 21: 371-378.
CrossRef - Azzazy, H.M.E. and W.E. Highsmith Jr., 2002. Phage display technology: Clinical applications and recent innovations. Clin. Biochem., 35: 425-445.
CrossRef - Tang, S.S., W.S. Tan, S. Devi, L.F. Wang, T. Pang and K.L. Thong, 2003. Mimotopes of the Vi Antigen of Salmonella enterica serovar Typhi identified from phage display peptide library. Clin. Diagn. Lab. Immunol., 10: 1078-1084.
CrossRef - Su, Z., T. Leung and J. Honek, 2006. Conformational selectivity of peptides for single walled carbon nanotube. J. Phys. Chem. B., 110: 23623-23627.
CrossRef - Na, C.H., M.H. Lee, B.Y. Cho and C. Chae, 2003. A method for ıdentification of the peptides that bind to a clone of thyroid-stimulating antibodies in the serum of graves disease patients. J. Clin. Endocrinol. Metab., 88: 1570-1576.
Direct Link - Zang, L., L. Shi, J. Guo, Q. Pan, W. Wua, X. Pan and J. Wang, 2009. Screening and identification of a peptide specifically targeted to NCI-H1299 from a phage display peptide library. Cancer Lett., 281: 64-70.
Direct Link - Lee, S.J., K.H. Kim, J.S. Park, J.W. Jung and Y.H. Kim et al., 2007. Comparative analysis of cell surface proteins in chronic and acute leukemia cell lines. Biochem. Biophys. Res. Commun., 357: 620-626.
PubMed - Dickerson, M.B., R.R. Naik, M.O. Stone, Y. Cai and K.H. Sandhage, 2004. Identification of peptides that promote the rapid precipitation of germania nanoparticle networks via use of a peptide display library. Chem. Commun., 7: 1776-1777.
PubMed - Chua, K.H., C.C. Hwa and S.D. Puthucheary, 2008. Identification of Burkholderia pseudomallei mimotope using phage display approach. J. Applied Sci., 8: 3999-4003.
CrossRefDirect Link - Sergeeva, A., M.G. Kolonin, J.J. Molldrem, R. Pasqualini and W. Arap, 2006. Display technologies: application for the discovery of drug and gene delivery agents. Adv. Drug Delivery Rev., 58: 1622-1654.
CrossRefPubMedDirect Link - Benhar, I., 2001. Biotechnological applications of phage and cell display. Biotechnol. Adv., 19: 1-33.
Direct Link - Arora, A., K. Seth and Y. Shukla, 2004. Reversal of P-glycoprotein-mediated multidrug resistance by diallyl sulfide in K562 leukemic cells and in mouse liver. Carcinogenesis, 25: 941-949.
PubMed